
Vitamin D as a Nutri-Epigenetic Factor in Autoimmunity—A Review of Current Research and Reports on Vitamin D Deficiency in Autoimmune Diseases


{kind=link}
Abstract
:1. Introduction to Epigenetic Modifications
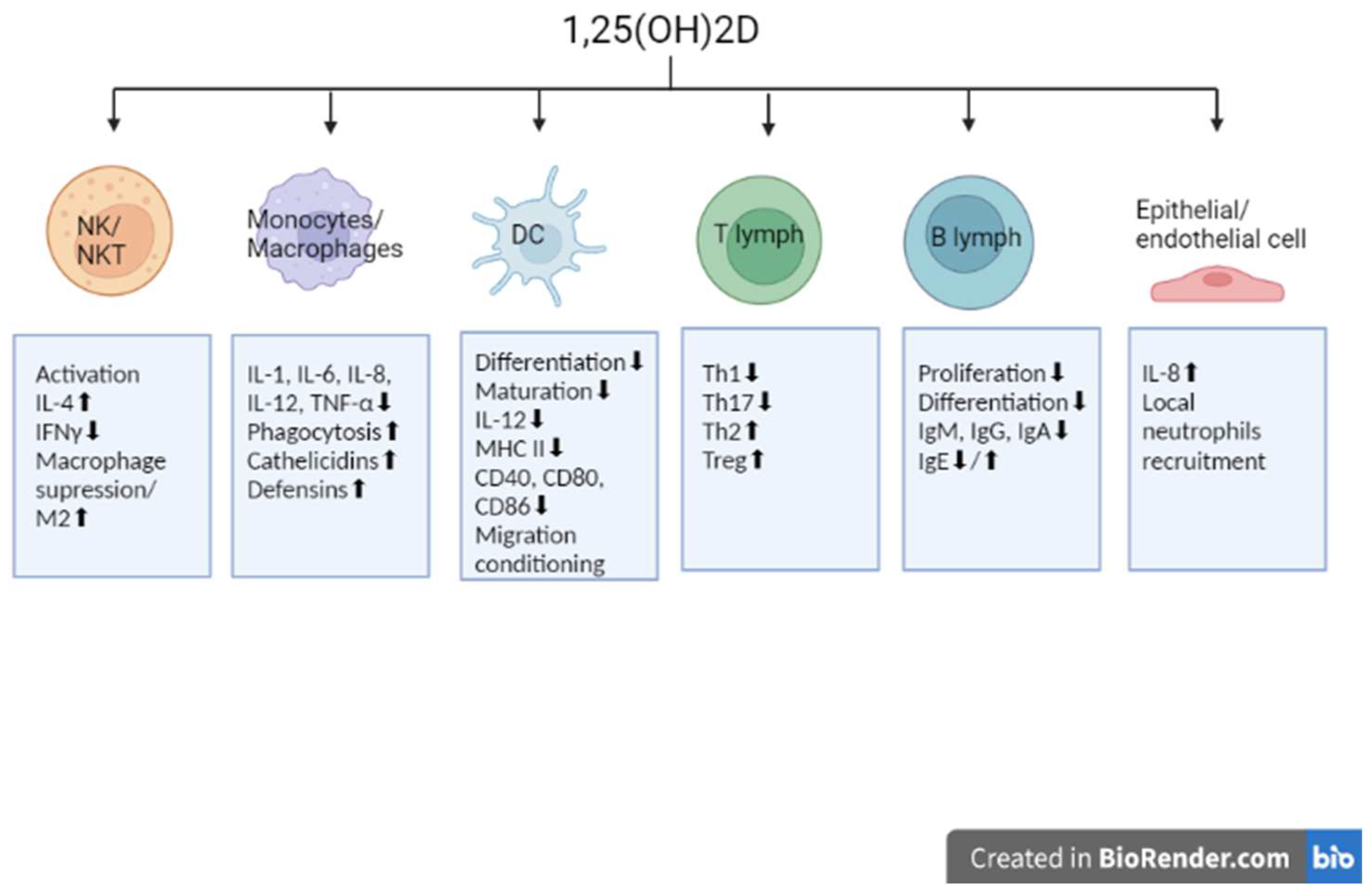
2. Involvement of Vitamin D in Immunomodulation
3. Vitamin D Deficiency and Epigenetic Dysregulations in Autoimmunity
3.1. Systemic Autoimmune Diseases
3.2. Organ-Specific Autoimmune Diseases
4. Current Clinical Trials Using Vitamin D in Autoimmune Diseases
- “Pilot Study of OMEGA-3 and Vitamin D in High-Dose in Type I Diabetic Patients (POSEIDON)”—NCT03406897. A recruiting interventional open-label study with an estimated 56 participants. The authors aim to evaluate the efficacy and safety of a treatment regimen based on omega-3 fatty acids and vitamin D in patients with T1D. The authors hypothesize that the used combination administered to patients with new or established forms of the disease will be safe and preserve insulin secretion [120];
- “Early High-Dose Vitamin D and Residual β-Cell Function in Pediatric Type 1 Diabetes”—NCT05270343. Not yet recruiting interventional open-label study with an estimated 198 participants. The aim of the project is to evaluate the effect of high-dose vitamin D supplementation on early T1D in children and adolescents. Patients simultaneously require intensive insulin therapy [121];
- “Effect of Vitamin D Supplement on Disease Activity in SLE”—NCT05260255. The purpose of the ongoing study is to evaluate the effect of vitamin D supplementation on SLE activity (Systemic Lupus Erythematosus Disease Activity Index 2000). A recruiting interventional double-blind study with an estimated 100 participants. The study is also expected to assess IL-6 levels and anti-dsDNA titers at specific intervention intervals [122];
- “The Vitamin D in Pediatric Crohn’s Disease (ViDiPeC-2) (ViDiPeC-2)”—NCT03999580. The aim of the study is to evaluate the effectiveness of high-dose vitamin D supplementation in children with CD. The researchers expect to achieve a reduction in the frequency of relapses and improved patient quality of life [123];
- “High Dose Interval Vitamin D Supplementation in Patients with Inflammatory Bowel Disease Receiving Biologic Therapy”—NCT04331639. A recruiting interventional open-label study with an estimated 50 participants. Vitamin D will be administered concurrently with IBD biologic therapy every 4–8 weeks. The researchers aim to evaluate, using laboratory tests and a questionnaire, inflammatory markers as well as markers of bone health after intervention with vitamin D [124];
- “Longitudinal Effect of Vitamin D3 Replacement on Cognitive Performance and MRI Markers in Multiple Sclerosis Patients”—NCT03610139. A recruiting interventional single-blind study with an estimated 162 participants. The researchers hypothesize that high-dose vitamin D supplementation will result in improvements in cognitive performance at 6 and 12 months after supplementation [125].
5. Conclusions
Funding
Conflicts of Interest
References
- Sedley, L. Advances in Nutritional Epigenetics—A Fresh Perspective for an Old Idea. Lessons Learned, Limitations, and Future Directions. Epigenet. Insights 2020, 13, 2516865720981924. [Google Scholar] [CrossRef] [PubMed]
- Felsenfeld, G. A brief history of epigenetics. Cold Spring Harb. Perspect. Biol. 2014, 6, a018200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, J.K.; Dawlaty, M.M.; Verma, A.; Maitra, A. Roles and Regulations of TET Enzymes in Solid Tumors. Trends Cancer 2021, 7, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Zhang, Z.M.; Xia, Y.; Liao, G.-Q.; Pan, Y.; Liu, S.; Zhang, Y.; Yan, Z.-S. LSD1-mediated epigenetic modification contributes to proliferation and metastasis of colon cancer. Br. J. Cancer 2013, 109, 994–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fetahu, I.S.; Höbaus, J.; Kállay, E. Vitamin D and the epigenome. Front. Physiol. 2014, 5, 164. [Google Scholar] [CrossRef] [Green Version]
- Anaya, J.M.; Shoenfeld, Y.; Rojas-Villarraga, A.; Levy, R.A.; Cervera, R. (Eds.) Autoimmunity: From Bench to Bedside; El Rosario University Press: Bogota, Colombia, 2013. [Google Scholar]
- Tapp, H.S.; Commane, D.M.; Bradburn, D.M.; Arasaradnam, R.; Mathers, J.C.; Johnson, I.; Belshaw, N.J. Nutritional factors and gender influence age-related DNA methylation in the human rectal mucosa. Aging Cell 2013, 12, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Rawson, J.B.; Sun, Z.; Dicks, E.; Daftary, D.; Parfrey, P.S.; Green, R.C.; Gallinger, S.; McLaughlin, J.R.; Wang, P.P.; Knight, J.A.; et al. Vitamin D intake is negatively associated with promoter methylation of the Wnt antagonist gene DKK1 in a large group of colorectal cancer patients. Nutr. Cancer 2012, 64, 919–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, N.; Carvalho, J.; Durães, C.; Sousa, B.; Gomes, M.; Costa, J.L.; Oliveira, C.; Parades, J.; Schmitt, F. 1Alpha,25-dihydroxyvitamin D3 induces de novo E-cadherin expression in triple-negative breast cancer cells by CDH1-promoter demethylation. Anticancer Res. 2012, 32, 249–257. [Google Scholar]
- Vanoirbeek, E.; Eelen, G.; Verlinden, L.; Carmeliet, G.; Mathieu, C.; Bouillon, R.; O’Connor, R.; Xiao, G.; Verstuyf, A. PDLIM2 expression is driven by vitamin D and is involved in the pro-adhesion, and anti-migration and -invasion activity of vitamin D. Oncogene 2014, 33, 1904–1911. [Google Scholar] [CrossRef]
- Pereira, F.; Barbáchano, A.; Silva, J.; Bonilla, F.; Campbell, M.J.; Muñoz, A.; Larriba, M.J. KDM6B/JMJD3 histone demethylase is induced by vitamin D and modulates its effects in colon cancer cells. Hum. Mol. Genet. 2011, 20, 4655–4665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padi, S.K.; Zhang, Q.; Rustum, Y.M.; Morrison, C.; Guo, B. MicroRNA-627 mediates the epigenetic mechanisms of vitamin D to suppress proliferation of human colorectal cancer cells and growth of xenograft tumors in mice. Gastroenterology 2013, 145, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, W.; Wei, B.; Wu, K.; Liu, D.; Zhu, D.; Zhang, C.; Wen, F.; Fan, Y.; Zhao, S. MicroRNA let-7i Inhibits Histone Lysine Demethylase KDM5B to Halt Esophageal Cancer Progression. Mol. Ther. Nucleic Acids. 2020, 22, 846–861. [Google Scholar] [CrossRef]
- Mazzone, R.; Zwergel, C.; Artico, M.; Taurone, S.; Ralli, M.; Greco, A.; Mai, A. The emerging role of epigenetics in human autoimmune disorders. Clin. Epigenet. 2019, 11, 34. [Google Scholar] [CrossRef] [Green Version]
- Surace, A.E.A.; Hedrich, C.M. The Role of Epigenetics in Autoimmune/Inflammatory Disease. Front. Immunol. 2019, 10, 1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslani, S.; Mahmoudi, M.; Karami, J.; Jamshidi, A.R.; Malekshahi, Z.; Nicknam, M.H. Epigenetic alterations underlying autoimmune diseases. Autoimmunity 2016, 49, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Bikle, D.D. Vitamin D: Production, Metabolism and Mechanisms of Action. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dhatariya, K., Dungan, K., Hershman, J.M., Hofland, J., Kalra, S., et al., Eds.; MDText.com, Inc.: Dartmouth, MA, USA, 2021. [Google Scholar]
- Taha, R.; Abureesh, S.; Alghamdi, S.; Hassan, R.Y.; Cheikh, M.M.; Bagabir, R.A.; Almoallim, H.; Abdulkhaliq, A. The Relationship between Vitamin D and Infections Including COVID-19: Any Hopes? Int. J. Gen. Med. 2021, 14, 3849–3870. [Google Scholar] [CrossRef] [PubMed]
- Snegarova, V.; Naydenova, D. Vitamin D: A Review of its Effects on Epigenetics and Gene Regulation. Folia Med. 2020, 62, 662–667. [Google Scholar] [CrossRef]
- Bizzaro, G.; Antico, A.; Fortunato, A.; Bizzaro, N. Vitamin D and Autoimmune Diseases: Is Vitamin D Receptor (VDR) Polymorphism the Culprit? Isr. Med. Assoc. J. 2017, 19, 438–443. [Google Scholar] [PubMed]
- Christakos, S.; Dhawan, P.; Verstuyf, A.; Verlinden, L.; Carmeliet, G. Vitamin D: Metabolism, Molecular Mechanism of Action, and Pleiotropic Effects. Physiol. Rev. 2016, 96, 365–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, S.; Cai, J.; Li, Y.; Yang, R. 1,25-Dihydroxy-Vitamin D3 induces macrophage polarization to M2 by upregulating T-cell Ig-mucin-3 expression. Mol. Med. Rep. 2019, 19, 3707–3713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nouari, W.; Ysmail-Dahlouk, L.; Aribi, M. Vitamin D3 enhances the bactericidal activity of macrophage against Pseudomonas aeruginosa. Int. Immunopharmacol. 2016, 30, 94–101. [Google Scholar] [CrossRef]
- Selvaraj, P. Vitamin D, vitamin D receptor, and cathelicidin in the treatment of tuberculosis. Vitam. Horm. 2011, 86, 307–325. [Google Scholar] [CrossRef]
- Qayyum, S.; Mohammad, T.; Slominski, R.M.; Hassan, I.; Tuckey, R.C.; Raman, C.; Slominski, A.T. Vitamin D and lumisterol novel metabolites can inhibit SARS-CoV-2 replication machinery enzymes. Am. J. Physiol. Endocrinol. Metab. 2021, 321, E246–E251. [Google Scholar] [CrossRef]
- Tomaszewska, A.; Rustecka, A.; Lipińska-Opałka, A.; Piprek, R.P.; Kloc, M.; Kalicki, B.; Kubiak, J.Z. The Role of Vitamin D in COVID-19 and the Impact of Pandemic Restrictions on Vitamin D Blood Content. Front. Pharmacol. 2022, 13, 836738. [Google Scholar] [CrossRef]
- Chen, L.; Eapen, M.S.; Zosky, G.R. Vitamin D both facilitates and attenuates the cellular response to lipopolysaccharide. Sci. Rep. 2017, 7, 45172. [Google Scholar] [CrossRef] [Green Version]
- Aranow, C. Vitamin D and the immune system. J. Investig. Med. 2011, 59, 881–886. [Google Scholar] [CrossRef] [Green Version]
- Bscheider, M.; Butcher, E.C. Vitamin D immunoregulation through dendritic cells. Immunology 2016, 148, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Fisher, S.A.; Rahimzadeh, M.; Brierley, C.; Gration, B.; Doree, C.; Kimber, C.E.; Cajide, A.P.; Lamikanra, A.A.; Roberts, D.J. The role of vitamin D in increasing circulating T regulatory cell numbers and modulating T regulatory cell phenotypes in patients with inflammatory disease or in healthy volunteers: A systematic review. PLoS ONE 2019, 14, e0222313. [Google Scholar] [CrossRef] [Green Version]
- Cantorna, M.T.; Snyder, L.; Lin, Y.D.; Yang, L. Vitamin D and 1,25(OH)2D regulation of T cells. Nutrients 2015, 7, 3011–3021. [Google Scholar] [CrossRef] [Green Version]
- Slominski, A.T.; Kim, T.K.; Takeda, Y.; Janjetovic, Z.; Brożyna, A.A.; Skobowiat, C.; Wang, J.; Postlethwaite, A.; Li, W.; Tuckey, R.C.; et al. RORα and ROR γ are expressed in human skin and serve as receptors for endogenously produced noncalcemic 20-hydroxy- and 20,23-dihydroxyvitamin D. FASEB J. 2014, 28, 2775–2789. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Sims, G.P.; Chen, X.X.; Gu, Y.Y.; Chen, S.; Lipsky, P.E. Modulatory effects of 1,25-dihydroxyvitamin D3 on human B cell differentiation. J. Immunol. 2007, 179, 1634–1647. [Google Scholar] [CrossRef]
- Rolf, L.; Muris, A.H.; Hupperts, R.; Damoiseaux, J. Vitamin D effects on B cell function in autoimmunity. Ann. N. Y. Acad. Sci. 2014, 1317, 84–91. [Google Scholar] [CrossRef]
- Cyprian, F.; Lefkou, E.; Varoudi, K.; Girardi, G. Immunomodulatory Effects of Vitamin D in Pregnancy and Beyond. Front. Immunol. 2019, 10, 2739. [Google Scholar] [CrossRef] [Green Version]
- Mora, J.R.; Iwata, M.; von Andrian, U.H. Vitamin effects on the immune system: Vitamins A and D take centre stage. Nat. Rev. Immunol. 2008, 8, 685–698. [Google Scholar] [CrossRef] [Green Version]
- Amrein, K.; Scherkl, M.; Hoffmann, M.; Neuwersch-Sommeregger, S.; Köstenberger, M.; Berisha, A.T.; Martucci, G.; Pilz, S.; Malle, O. Vitamin D deficiency 2.0: An update on the current status worldwide. Eur. J. Clin. Nutr. 2020, 74, 1498–1513. [Google Scholar] [CrossRef]
- Rusińska, A.; Płudowski, P.; Walczak, M.; Borszewska-Kornacka, M.K.; Bossowski, A.; Chlebna-Sokół, D.; Czech-Kowalska, J.; Dobrzańska, A.; Franek, E.; Helwich, E.; et al. Zasady Suplementacji I Leczenia Witaminą D—Nowelizacja 2018 R. Stand. Med. Pediatr. 2018, 15, 531–559. [Google Scholar]
- Siddiqee, M.H.; Bhattacharjee, B.; Siddiqi, U.R.; MeshbahurRahman, M. High prevalence of vitamin D deficiency among the South Asian adults: A systematic review and meta-analysis. BMC Public Health 2021, 21, 1823. [Google Scholar] [CrossRef]
- Lips, P.; Cashman, K.D.; Lamberg-Allardt, C.; Bischoff-Ferrari, H.A.; Obermayer-Pietsch, B.; Bianchi, M.L.; Stepan, J.; El-Hajj Fuleihan, G.; Bouillon, R. Current vitamin D status in European and Middle East countries and strategies to prevent vitamin D deficiency: A position statement of the European Calcified Tissue Society. Eur. J. Endocrinol. 2019, 180, P23–P54. [Google Scholar] [CrossRef] [Green Version]
- Cashman, K.D. Vitamin D Deficiency: Defining, Prevalence, Causes, and Strategies of Addressing. Calcif. Tissue Int. 2020, 106, 14–29. [Google Scholar] [CrossRef]
- Kanike, N.; Kannekanti, N.; Camacho, J. Vitamin D Deficiency in Pregnant Women and Newborn. In Vitamin D; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Özdemir, A.A.; Gündemir, Y.E.; Küçük, M.; Sarıcı, D.Y.; Elgörmüş, Y.; Çağ, Y.; Bilek, G. Vitamin D Deficiency in Pregnant Women and Their Infants. J. Clin. Res. Pediatr. Endocrinol. 2018, 10, 44–50. [Google Scholar] [CrossRef]
- Palacios, C.; Gonzales, L. Is vitamin D deficiency a major global public health problem? J. Steroid Biochem. Mol. Biol. 2014, 144, 138–145. [Google Scholar] [CrossRef]
- Prentice, A. Vitamin D deficiency: A global perspective. Nutr. Rev. 2008, 66, 153–164. [Google Scholar] [CrossRef]
- Wagner, C.L.; Hollis, B.W. The Implications of Vitamin D Status During Pregnancy on Mother and her Developing Child. Front. Endocrinol. 2018, 9, 500. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, B.J.; Snoddy, A.M.E.; Munns, C.; Simm, P.; Siafarikas, A.; Jefferies, C. A Brief History of Nutritional Rickets. Front. Endocrinol. 2019, 10, 795. [Google Scholar] [CrossRef]
- Hegazy, A.M.; Shinkar, D.M.; Mohamed, N.R.; Gaber, H.A. Association between serum 25 (OH) vitamin D level at birth and respiratory morbidities among preterm neonates. J. Matern. Fetal Neonatal Med. 2018, 31, 2649–2655. [Google Scholar] [CrossRef]
- Vinkhuyzen, A.A.E.; Eyles, D.W.; Burne, T.H.J.; Blanken, L.M.; Kruithof, C.J.; Verhulst, F.; Jaddoe, V.W.; Tiemeier, H.; McGrath, J.J. Gestational vitamin D deficiency and autism-related traits: The Generation R Study. Mol. Psychiatry 2018, 23, 240–246. [Google Scholar] [CrossRef] [Green Version]
- McGrath, J.J.; Burne, T.H.; Féron, F.; Mackay-Sim, A.; Eyles, D.W. Developmental vitamin D deficiency and risk of schizophrenia: A 10-year update. Schizophr. Bull. 2010, 36, 1073–1078. [Google Scholar] [CrossRef]
- Zold, E.; Szodoray, P.; Gaal, J.; Kappelmayer, J.; Csathy, L.; Gyimesi, E.; Zeher, M.; Szegedi, G.; Bodolay, E. Vitamin D deficiency in undifferentiated connective tissue disease. Arthritis. Res. Ther. 2008, 10, R123. [Google Scholar] [CrossRef] [Green Version]
- Guan, S.Y.; Cai, H.Y.; Wang, P.; Lv, T.; Liu, L.; Mao, Y.; Zhao, C.; Wu, Q.; Dan, Y.; Sam, N.B.; et al. Association between circulating 25-hydroxyvitamin D and systemic lupus erythematosus: A systematic review and meta-analysis. Int. J. Rheum. Dis. 2019, 22, 1803–1813. [Google Scholar] [CrossRef]
- Mazzone, J.W.; Montoya, T.; Castejón, M.L.; Muñoz-García, R.; Alarcón-de-la-Lastra, C. Epigenetic linkage of systemic lupus erythematosus and nutrition. Nutr. Res. Rev. 2021, 1–21. [Google Scholar] [CrossRef]
- Yang, S.K.; Liu, N.; Zhang, W.J.; Song, N.; Yang, J.-P.; Zhang, H.; Gui, M. Impact of Vitamin D Receptor Gene Polymorphism on Systemic Lupus Eryt.hematosus Susceptibility: A Pooled Analysis. Genet. Test. Mol. Biomark. 2022, 26, 228–238. [Google Scholar] [CrossRef]
- Yaylim-Eraltan, I.; Ergen, H.A.; Arikan, S.; Okay, E.; Öztürk, O.; Bayrak, S.; Isbir, T. Investigation of the VDR gene polymorphisms association with susceptibility to colorectal cancer. Cell Biochem. Funct. 2007, 25, 731–737. [Google Scholar] [CrossRef]
- Aparicio-Soto, M.; Sánchez-Hidalgo, M.; Alarcón-de-la-Lastra, C. An update on diet and nutritional factors in systemic lupus erythematosus management. Nutr. Res. Rev. 2017, 30, 118–137. [Google Scholar] [CrossRef]
- Karouzakis, E.; Gay, R.E.; Gay, S.; Neidhart, M. Increased recycling of polyamines is associated with global DNA hypomethylation in rheumatoid arthritis synovial fibroblasts. Arthritis Rheum. 2012, 64, 1809–1817. [Google Scholar] [CrossRef]
- Zhang, T.P.; Li, H.M.; Huang, Q.; Wang, L.; Li, X.M. Vitamin D Metabolic Pathway Genes Polymorphisms and Their Methylation Levels in Association with Rheumatoid Arthritis. Front. Immunol. 2021, 12, 731565. [Google Scholar] [CrossRef] [PubMed]
- Punceviciene, E.; Gaizevska, J.; Sabaliauskaite, R.; Venceviciene, L.; Puriene, A.; Vitkus, D.; Jarmalaite, S.; Butrimiene, I. Vitamin D and VDR Gene Polymorphisms’ Association with Rheumatoid Arthritis in Lithuanian Population. Medicina 2021, 57, 346. [Google Scholar] [CrossRef]
- Garcia-Carrasco, M.; Jiménez-Herrera, E.A.; Gálvez-Romero, J.L.; de Lara, L.V.; Mendoza-Pinto, C.; Etchegaray-Morales, I.; Munguía-Realpozo, P.; Ruíz-Argüelles, A.; Jose, R.; Vera-Recabarren, M.; et al. Vitamin D and Sjögren syndrome. Autoimmun. Rev. 2017, 16, 587–593. [Google Scholar] [CrossRef]
- Agmon-Levin, N.; Kivity, S.; Tzioufas, A.G.; Hoyos, M.L.; Rozman, B.; Efes, I.; Shapira, Y.; Shamis, A.; Amital, H.; Youinou, P.; et al. Low levels of vitamin-D are associated with neuropathy and lymphoma among patients with Sjögren’s syndrome. J Autoimmun. 2012, 39, 234–239. [Google Scholar] [CrossRef]
- Kiraly, S.J.; Kiraly, M.A.; Hawe, R.D.; Makhani, N. Vitamin D as a neuroactive substance: Review. Sci. World J. 2006, 6, 125–139. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, R.; Bunya, V.Y.; Pistilli, M.; Ying, G.-S.; Macchi, I.; Vivino, F.; Massaro-Giordano, M. Vitamin D and Neuropathy in Patients with Sjogren’s Syndrome. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6162. [Google Scholar]
- Ambrosi, A.; Salomonsson, S.; Eliasson, H.; Zeffer, E.; Skog, A.; Dzikaite, V.; Bergman, G.; Fernlund, E.; Tingström, J.; Theander, E.; et al. Development of heart block in children of SSA/SSB-autoantibody-positive women is associated with maternal age and displays a season-of-birth pattern. Ann. Rheum. Dis. 2012, 71, 334–340. [Google Scholar] [CrossRef] [Green Version]
- Tincani, A.; Andreoli, L.; Cavazzana, I.; Doria, A.; Favero, M.; Fenini, M.-G.; Franceschini, F.; Lojacono, A.; Nascimbeni, G.; Santoro, A.; et al. Novel aspects of Sjögren’s syndrome in 2012. BMC Med. 2013, 11, 93. [Google Scholar] [CrossRef] [PubMed]
- Imgenberg-Kreuz, J.; Rasmussen, A.; Sivils, K.; Nordmark, G. Genetics and epigenetics in primary Sjögren’s syndrome. Rheumatology 2021, 60, 2085–2098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zilahi, E.; Chen, J.Q.; Papp, G.; Szántó, A.; Zeher, M. Lack of association of vitamin D receptor gene polymorphisms/haplotypes in Sjögren’s syndrome. Clin. Rheumatol. 2015, 34, 247–253. [Google Scholar] [CrossRef] [Green Version]
- Sampaio-Barros, M.M.; Takayama, L.; Sampaio-Barros, P.D.; Bonfá, E.; Pereira, R.M. Low vitamin D serum levels in diffuse systemic sclerosis: A correlation with worst quality of life and severe capillaroscopic findings. Rev. Bras. Reumatol. 2016, 56, 337–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groseanu, L.; Bojinca, V.; Gudu, T.; Saulescu, I.; Predeteanu, D.; Balanescu, A.; Berghea, F.; Opriş, D.; Borangiu, A.; Constantinescu, C.; et al. Low vitamin D status in systemic sclerosis and the impact on disease phenotype. Eur. J. Rheumatol. 2016, 3, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, S.Y.; Liu, H.H.; Yin, X.-D.; Cao, L.-T.; Xu, J.-H.; Li, X.-M.; Ye, D.-Q.; Wang, J. Associations of Vitamin D Receptor Single Nucleotide Polymorphisms with Susceptibility to Systemic Sclerosis. Arch. Med. Res. 2019, 50, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Schneider, L.; Hax, V.; Monticielo, O.; Macedo, T.F.; Barreto, R.K.M.; Marcondes, N.A.; Chakr, R. Dualities of the vitamin D in systemic sclerosis: A systematic literature review. Adv. Rheumatol. 2021, 61, 34. [Google Scholar] [CrossRef]
- Liu, W.; Chen, Y.; Golan, M.A.; Annunziata, M.L.; Du, J.; Dougherty, U.; Kong, J.; Musch, M.; Huang, Y.; Pekow, J.; et al. Intestinal epithelial vitamin D receptor signaling inhibits experimental colitis. J. Clin. Investig. 2013, 123, 3983–3996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakke, D.; Sun, J. Ancient Nuclear Receptor VDR with New Functions: Microbiome and Inflammation. Inflamm. Bowel Dis. 2018, 24, 1149–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Zhang, H.; Wu, H.; Li, H.; Liu, L.; Guo, J.; Li, C.; Shih, D.Q.; Zhang, X. Protective role of 1,25(OH)2 vitamin D3 in the mucosal injury and epithelial barrier disruption in DSS-induced acute colitis in mice. BMC Gastroenterol. 2012, 12, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vernia, F.; Valvano, M.; Longo, S.; Cesaro, N.; Viscido, A.; Latella, G. Vitamin D in Inflammatory Bowel Diseases. Mechanisms of Action and Therapeutic Implications. Nutrients 2022, 14, 269. [Google Scholar] [CrossRef] [PubMed]
- López-Muñoz, P.; Beltrán, B.; Sáez-González, E.; Alba, A.; Nos, P.; Iborra, M. Influence of Vitamin D Deficiency on Inflammatory Markers and Clinical Disease Activity in IBD Patients. Nutrients 2019, 11, 1059. [Google Scholar] [CrossRef] [Green Version]
- Schäffler, H.; Herlemann, D.P.; Klinitzke, P.; Berlin, P.; Kreikemeyer, B.; Jaster, R.; Lamprecht, G. Vitamin D administration leads to a shift of the intestinal bacterial composition in Crohn’s disease patients, but not in healthy controls. J. Dig. Dis. 2018, 19, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Garg, M.; Hendy, P.; Ding, J.N.; Shaw, S.; Hold, G.; Hart, A. The Effect of Vitamin D on Intestinal Inflammation and Faecal Microbiota in Patients with Ulcerative Colitis. J. Crohn’s Colitis 2018, 12, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Pagnini, C.; Di Paolo, M.C.; Graziani, M.G.; Delle Fave, G. Probiotics and Vitamin D/Vitamin D Receptor Pathway Interaction: Potential Therapeutic Implications in Inflammatory Bowel Disease. Front. Pharmacol. 2021, 12, 747856. [Google Scholar] [CrossRef] [PubMed]
- Kohil, A.; Al-Asmakh, M.; Al-Shafai, M.; Terranegra, A. The Interplay between Diet and the Epigenome in the Pathogenesis of Type-1 Diabetes. Front. Nutr. 2021, 7, 612115. [Google Scholar] [CrossRef]
- Hyöty, H.; Leon, F.; Knip, M. Developing a vaccine for type 1 diabetes by targeting coxsackievirus B. Expert Rev. Vaccines 2018, 17, 1071–1083. [Google Scholar] [CrossRef]
- Rewers, M.; Ludvigsson, J. Environmental risk factors for type 1 diabetes. Lancet 2016, 387, 2340–2348. [Google Scholar] [CrossRef] [Green Version]
- Butalia, S.; Kaplan, G.G.; Khokhar, B.; Rabi, D.M. Environmental risk factors and type 1 diabetes: Past, present, and future. Can. J. Diabetes 2016, 40, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Cerna, M. Epigenetic Regulation in Etiology of Type 1 Diabetes Mellitus. Int. J. Mol. Sci. 2019, 21, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Chen, L.M.; Zou, Y.; Zhang, S.; Xiong, F.; Wang, C.Y. Implication of epigenetic factors in the pathogenesis of type 1 diabetes. Chin. Med. J. 2021, 134, 1031–1042. [Google Scholar] [CrossRef]
- Ziegler, A.G.; Schmid, S.; Huber, D.; Hummel, M.; Bonifacio, E. Early infant feeding and risk of developing type 1 diabetes-associated autoantibodies. JAMA 2003, 290, 1721–1728. [Google Scholar] [CrossRef] [Green Version]
- Greer, R.M.; Rogers, M.A.; Bowling, F.G.; Buntain, H.M.; Harris, M.; Leong, G.; Cotterill, A.M. Australian children and adolescents with type 1 diabetes have low vitamin D levels. Med. J. Aust. 2007, 187, 59–60. [Google Scholar] [CrossRef]
- Borkar, V.V.; Devidayal; Verma, S.; Bhalla, A.K. Low levels of vitamin D in North Indian children with newly diagnosed type 1 diabetes. Pediatr. Diabetes 2010, 11, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Littorin, B.; Blom, P.; Schölin, A.; Arnqvist, H.J.; Blohmé, G.; Bolinder, J.; Ekbom-Schnell, A.; Eriksson, J.W.; Gudbjörnsdottir, S.; Nyström, L.; et al. Lower levels of plasma 25-hydroxyvitamin D among young adults at diagnosis of autoimmune type 1 diabetes compared with control subjects: Results from the nationwide Diabetes Incidence Study in Sweden (DISS). Diabetologia 2006, 49, 2847–2852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svoren, B.M.; Volkening, L.K.; Wood, J.R.; Laffel, L.M. Significant vitamin D deficiency in youth with type 1 diabetes mellitus. J. Pediatr. 2009, 154, 132–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habibian, N.; Amoli, M.M.; Abbasi, F.; Rabbani, A.; Alipour, A.; Sayarifard, F.; Rostami, P.; Dizaji, S.P.; Saadati, B.; Setoodeh, A. Role of vitamin D and vitamin D receptor gene polymorphisms on residual beta cell function in children with type 1 diabetes mellitus. Pharmacol. Rep. 2019, 71, 282–288. [Google Scholar] [CrossRef]
- Aljabri, K.S.; Bokhari, S.A.; Khan, M.J. Glycemic changes after vitamin D supplementation in patients with type 1 diabetes mellitus and vitamin D deficiency. Ann. Saudi Med. 2010, 30, 454–458. [Google Scholar] [CrossRef]
- Hyppönen, E.; Läärä, E.; Reunanen, A.; Järvelin, M.R.; Virtanen, S.M. Intake of vitamin D and risk of type 1 diabetes: A birth-cohort study. Lancet 2001, 358, 1500–1503. [Google Scholar] [CrossRef]
- Bierschenk, L.; Alexander, J.; Wasserfall, C.; Haller, M.; Schatz, D.; Atkinson, M. Vitamin D levels in subjects with and without type 1 diabetes residing in a solar rich environment. Diabetes Care 2009, 32, 1977–1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marjamäki, L.; Niinistö, S.; Kenward, M.G.; Uusitalo, U.; Ovaskainen, M.; Kronbergkippila, C.; Simell, O.; Veijola, R.; Ilonen, J.; Knip, M.; et al. Maternal intake of vitamin D during pregnancy and risk of advanced beta cell autoimmunity and type 1 diabetes in offspring. Diabetologia 2010, 53, 1599–1607. [Google Scholar] [CrossRef]
- Zhai, N.; Bidares, R.; Makoui, M.H.; Aslani, S.; Mohammadi, P.; Razi, B.; Imani, D.; Yazdchi, M.; Mikaeili, H. Vitamin D receptor gene polymorphisms and the risk of the type 1 diabetes: A meta-regression and updated meta-analysis. BMC Endocr. Disord. 2020, 20, 121. [Google Scholar] [CrossRef] [PubMed]
- Tizaoui, K.; Kaabachi, W.; Hamzaoui, A.; Hamzaoui, K. Contribution of VDR polymorphisms to type 1 diabetes susceptibility: Systematic review of case–control studies and meta-analysis. J. Steroid. Biochem. Mol. Biol. 2014, 143, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Altieri, B.; Muscogiuri, G.; Barrea, L.; Mathieu, C.; Vallone, C.V.; Mascitelli, L.; Bizzaro, G.; Altieri, V.M.; Tirabassi, G.; Balercia, G.; et al. Does vitamin D play a role in autoimmune endocrine disorders? A proof of concept. Rev. Endocr. Metab. Disord. 2017, 18, 335–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, E.R.; Zwermann, O.; Segni, M.; Meyer, G.; Reincke, M.; Seissler, J.; Herwig, J.; Usadel, K.H.; Badenhoop, K. A promoter polymorphism of the CYP27B1 gene is associated with Addison’s disease, Hashimoto’s thyroiditis, Graves’ disease, and type 1 diabetes mellitus in Germans. Eur. J. Endocrinol. 2004, 151, 193–197. [Google Scholar] [CrossRef] [Green Version]
- Jennings, C.E.; Owen, C.J.; Wilson, V.; Pearce, S.H. A haplotype of the CYP27B1 promoter is associated with autoimmune Addison’s disease but not with Graves’ disease in a UK population. J. Mol. Endocrinol. 2005, 34, 859–863. [Google Scholar] [CrossRef] [Green Version]
- Fichna, M.; Zurawek, M.; Januszkiewicz-Lewandowska, D.; Gryczynska, M.; Fichna, P.; Sowinski, J. Association of the CYP27B1 C(-1260)A polymorphism with autoimmune Addison’s disease. Exp. Clin. Endocrinol. Diabetes 2010, 118, 544–549. [Google Scholar] [CrossRef]
- Weetman, A.P. The immunopathogenesis of chronic autoimmune thyroiditis one century after hashimoto. Eur. Thyroid J. 2013, 1, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Stefan-Lifshitz, M.; Li, C.W.; Tomer, Y. Genetics and epigenetics of autoimmune thyroid diseases: Translational implications. Best Pract. Res. Clin. Endocrinol. Metab. 2022, 101661. [Google Scholar] [CrossRef]
- Feng, M.; Li, H.; Chen, S.F.; Li, W.F.; Zhang, F.B. Polymorphisms in the vitamin D receptor gene and risk of autoimmune thyroid diseases: A meta-analysis. Endocrine 2013, 43, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Galușca, D.; Popoviciu, M.S.; Babeș, E.E.; Vidican, M.; Zaha, A.A.; Babeș, V.V.; Jurca, A.D.; Zaha, D.C.; Bodog, F. Vitamin D Implications and Effect of Supplementation in Endocrine Disorders: Autoimmune Thyroid Disorders (Hashimoto’s Disease and Grave’s Disease), Diabetes Mellitus and Obesity. Medicina 2022, 58, 194. [Google Scholar] [CrossRef] [PubMed]
- Sheriba, N.; Elewa, A.A.; Mahdy, M.; El Din, A.B.; Ibrahim, N.; Marawan, D.; Moneim, T.A.E. Effect of vitamin D3 in treating hyperthyroidism in patients with Graves’ disease. Egypt. J. Intern. Med. 2017, 29, 64. [Google Scholar] [CrossRef]
- Küçükali, C.İ.; Kürtüncü, M.; Çoban, A.; Çebi, M.; Tüzün, E. Epigenetics of multiple sclerosis: An updated review. Neuromol. Med. 2015, 17, 83–96. [Google Scholar] [CrossRef]
- Gacias, M.; Casaccia, P. Epigenetic mechanisms in multiple sclerosis. Rev. Esp. Escler. Mult. 2014, 6, 25–35. [Google Scholar]
- Sintzel, M.B.; Rametta, M.; Reder, A.T. Vitamin D and Multiple Sclerosis: A Comprehensive Review. Neurol. Ther. 2018, 7, 59–85. [Google Scholar] [CrossRef] [Green Version]
- Sotirchos, E.S.; Bhargava, P.; Eckstein, C.; Van Haren, K.; Baynes, M.; Ntranos, A.; Gocke, A.; Steinman, L.; Mowry, E.M.; Calabresi, P.A. Safety and immunologic effects of high- vs. low-dose cholecalciferol in multiple sclerosis. Neurology 2016, 86, 382–390. [Google Scholar] [CrossRef]
- Díez, B.C.; Pérez-Ramírez, C.; Maldonado-Montoro, M.D.M.; Carrasco-Campos, M.I.; Martín, A.S.; Lancheros, L.E.P.; Martínez-Martínez, F.; Calleja-Hernández, M.; Ramírez-Tortosa, M.C.; Jiménez-Morales, A. Association between polymorphisms in the vitamin D receptor and susceptibility to multiple sclerosis. Pharm. Genom. 2021, 31, 40–47. [Google Scholar] [CrossRef]
- Imani, D.; Razi, B.; Motallebnezhad, M.; Rezaei, R. Association between vitamin D receptor (VDR) polymorphisms and the risk of multiple sclerosis (MS): An updated meta-analysis. BMC Neurol. 2019, 19, 339. [Google Scholar] [CrossRef] [Green Version]
- Ayuso, T.; Aznar, P.; Soriano, L.; Olaskoaga, A.; Roldán, M.; Otano, M.; Ajuria, I.; Soriano, G.; Lacruz, F.; Mendioroz, M. Vitamin D receptor gene is epigenetically altered and transcriptionally up-regulated in multiple sclerosis. PLoS ONE 2017, 12, e0174726. [Google Scholar] [CrossRef] [Green Version]
- Papagni, R.; Pellegrino, C.; Di Gennaro, F.; Patti, G.; Ricciardi, A.; Novara, R.; Cotugno, S.; Musso, M.; Guido, G.; Ronga, L.; et al. Impact of Vitamin D in Prophylaxis and Treatment in Tuberculosis Patients. Int. J. Mol. Sci. 2022, 23, 3860. [Google Scholar] [CrossRef] [PubMed]
- Crowle, A.J.; Ross, E.J.; May, M.H. Inhibition by 1,25(OH)2-vitamin D3 of the multiplication of virulent tubercle bacilli in cultured human macrophages. Infect. Immun. 1987, 55, 2945–2950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fletcher, J.; Bishop, E.L.; Harrison, S.R.; Swift, A.; Cooper, S.C.; Dimeloe, S.K.; Raza, K.; Hewison, M. Autoimmune disease and interconnections with vitamin D. Endocr. Connect. 2022, 11, e210554. [Google Scholar] [CrossRef]
- Feige, J.; Moser, T.; Bieler, L.; Schwenker, K.; Hauer, L.; Sellner, J. Vitamin D Supplementation in Multiple Sclerosis: A Critical Analysis of Potentials and Threats. Nutrients 2020, 12, 783. [Google Scholar] [CrossRef] [Green Version]
- Magro, R.; Saliba, C.; Camilleri, L.; Scerri, C.; Borg, A.A. Vitamin D supplementation in systemic lupus erythematosus: Relationship to disease activity, fatigue, and the interferon signature gene expression. BMC Rheumatol. 2021, 5, 53. [Google Scholar] [CrossRef]
- Hahn, J.; Cook, N.R.; Alexander, E.K.; Friedman, S.; Walter, J.; Bubes, V.; Kotler, G.; Lee, I.-M.; Manson, J.E.; Costenbader, K.H. Vitamin D and marine omega 3 fatty acid supplementation and incident autoimmune disease: VITAL randomized controlled trial. BMJ 2022, 376, e066452. [Google Scholar] [CrossRef]
- Available online: https://clinicaltrials.gov/ct2/show/study/NCT03406897 (accessed on 4 September 2022).
- Available online: https://clinicaltrials.gov/ct2/show/NCT05270343 (accessed on 4 September 2022).
- Available online: https://clinicaltrials.gov/ct2/show/NCT05260255 (accessed on 4 September 2022).
- Available online: https://clinicaltrials.gov/ct2/show/NCT03999580 (accessed on 4 September 2022).
- Available online: https://clinicaltrials.gov/ct2/show/NCT04331639 (accessed on 4 September 2022).
- Available online: https://clinicaltrials.gov/ct2/show/NCT03610139 (accessed on 4 September 2022).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazur, A.; Frączek, P.; Tabarkiewicz, J. Vitamin D as a Nutri-Epigenetic Factor in Autoimmunity—A Review of Current Research and Reports on Vitamin D Deficiency in Autoimmune Diseases. Nutrients 2022, 14, 4286. https://doi.org/10.3390/nu14204286
Mazur A, Frączek P, Tabarkiewicz J. Vitamin D as a Nutri-Epigenetic Factor in Autoimmunity—A Review of Current Research and Reports on Vitamin D Deficiency in Autoimmune Diseases. Nutrients. 2022; 14(20):4286. https://doi.org/10.3390/nu14204286
Chicago/Turabian StyleMazur, Artur, Paulina Frączek, and Jacek Tabarkiewicz. 2022. "Vitamin D as a Nutri-Epigenetic Factor in Autoimmunity—A Review of Current Research and Reports on Vitamin D Deficiency in Autoimmune Diseases" Nutrients 14, no. 20: 4286. https://doi.org/10.3390/nu14204286