Flavonoids and Other Polyphenols Act as Epigenetic Modifiers in Breast Cancer

Abstract
:1. Introduction
2. Methodology
2.1. Flavonoids
2.2. Worldwide Flavonoid Intake
2.3. Flavonoid/Polyphenol Intake and Incidence of Breast Cancer
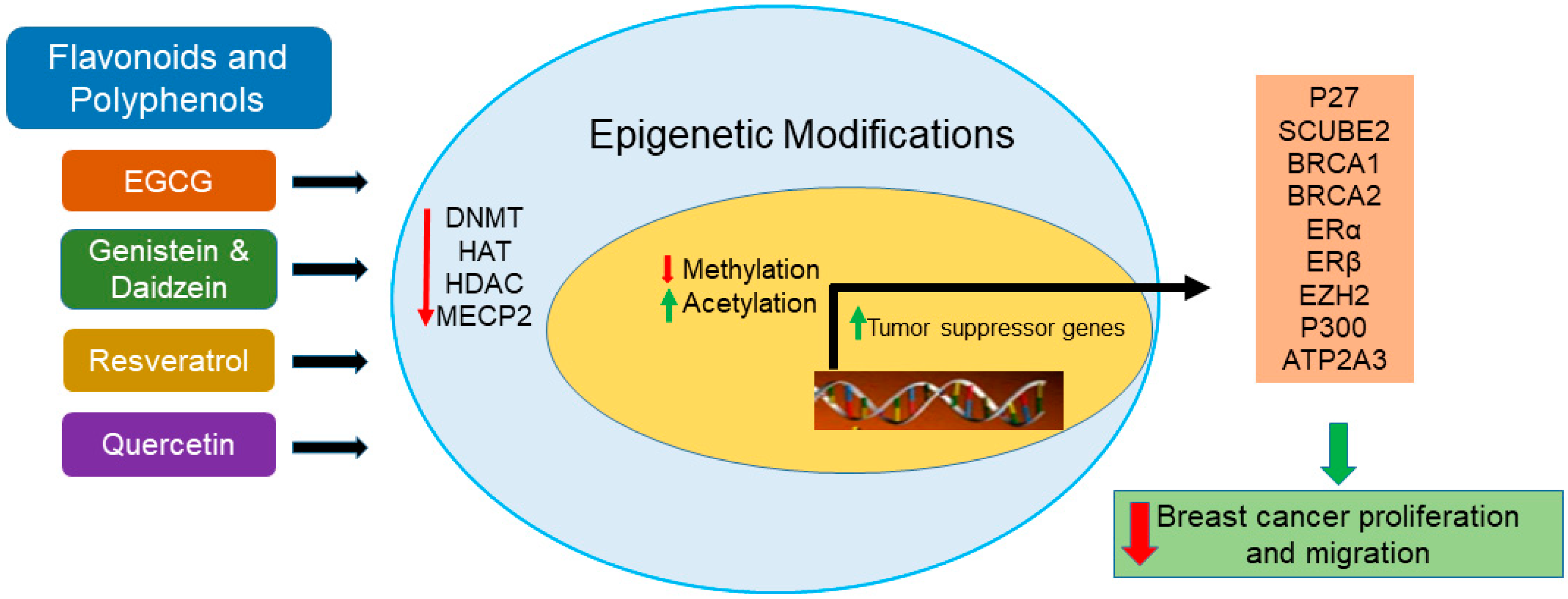
3. Epigenetics and Breast Cancer
3.1. Epigenetic Modifiers in Clinical Use
3.2. Epigallocatechin Regulates Tumor Suppressor Genes by Promoter Demethylation
3.3. Genistein and Daidzein Modulates Histone Modifications
3.4. Resveratrol Upregulates ATP2A3 Expression
3.5. Quercetin Modulates BRCA1 Expression Level
4. Conclusions and Further Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Nakhjavani, M.; Hardingham, J.E.; Palethorpe, H.M.; Price, T.J.; Townsend, A.R. Druggable Molecular Targets for the Treatment of Triple Negative Breast Cancer. J. Breast Cancer. 2019, 22, 341–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.L.; Kuo, Y.C.; Ho, Y.S.; Huang, Y.H. Triple-Negative Breast Cancer: Current Understanding and Future Therapeutic Breakthrough Targeting Cancer Stemness. Cancers (Basel) 2019, 11, 1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agurs-Collins, T.; Ross, S.A.; Dunn, B.K. The Many Faces of Obesity and Its Influence on Breast Cancer Risk. Front. Oncol. 2019, 9, 765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picon-Ruiz, M.; Morata-Tarifa, C.; Valle-Goffin, J.J.; Friedman, E.R.; Slingerland, J.M. Obesity and adverse breast cancer risk and outcome: Mechanistic insights and strategies for intervention. CA Cancer J. Clin. 2017, 67, 378–397. [Google Scholar] [CrossRef]
- American Institute for Cancer Research (AICR’s) Third Expert Report, Diet, Nutrition, Physical Activity, and Cancer: A Global Perspective. 2018. Available online: https://www.wcrf.org/sites/default/files/Breast-cancer-report.pdf (accessed on 3 March 2010).
- Howlader, N.; Noone, A.M.; Krapcho, M.; Miller, D.; Brest, A.; Yu, M.; Ruhl, J.; Tatalovich, Z.; Mariotto, A.; Lewis, D.R.; et al. (Eds.) SEER Cancer Statistics Review, 1975–2016, National Cancer Institute. Bethesda, MD, based on November 2018 SEER Data Submission, Posted to the SEER Web Site. 2019. Available online: https://seer.cancer.gov/csr/1975_2016/ (accessed on 22 January 2020).
- Dietze, E.C.; Sistrunk, C.; Miranda-Carboni, G.; O’Regan, R.; Seewaldt, V.L. Triple-negative breast cancer in African-American women: Disparities versus biology. Nat. Rev. Cancer. 2015, 15, 248–254. [Google Scholar] [CrossRef] [Green Version]
- Mehrotra, J.; Ganpat, M.M.; Kanaan, Y.; Fackler, M.J.; McVeigh, M.; Lahti-Domenici, J.; Polyak, K.; Argani, P.; Naab, T.; Garrett, E.; et al. Estrogen receptor/progesterone receptor-negative breast cancers of youngAfrican-American women have a higher frequency of methylation of multiple genes than those of Caucasian women. Clin. Cancer Res. 2004, 10, 2052–2057. [Google Scholar] [CrossRef] [Green Version]
- Struewing, J.P.; Hartge, P.; Wacholder, S.; Baker, S.M.; Berlin, M.; McAdams, M.; Timmerman, M.M.; Brody, L.C.; Tucker, M.A. The risk of cancer associated with specific mutations of BRCA1 and BRCA2 among Ashkenazi Jews. N. Engl. J. Med. 1997, 336, 1401–1408. [Google Scholar] [CrossRef]
- Greenup, R.; Buchanan, A.; Lorizio, W.; Rhoads, K.; Chan, S.; Leedom, T.; King, R.; McLennan, J.; Crawford, B.; Kelly Marcom, P.; et al. Prevalence of BRCA mutations among women with triple-negative breast cancer (TNBC) in a genetic counseling cohort. Ann. Surg. Oncol. 2013, 20, 3254–3258. [Google Scholar] [CrossRef]
- López-Camarillo, C.; Gallardo-Rincón, D.; Álvarez-Sánchez, M.E.; Marchat, L.A. Pharmaco-epigenomics: On the Road of Translation Medicine. Adv. Exp. Med. Biol. 2019, 1168, 31–42. [Google Scholar] [PubMed]
- Shukla, S.; Penta, D.; Mondal, P.; Meeran, S.M. Epigenetics of Breast Cancer: Clinical Status of Epi-drugs and Phytochemicals. Adv. Exp. Med. Biol. 2019, 1152, 293–310. [Google Scholar] [PubMed]
- Davalos, V.; Martinez-Cardus, A.; Esteller, M. The Epigenomic Revolution in Breast Cancer: From Single-Gene to Genome-Wide Next-Generation Approaches. Am. J. Pathol. 2017, 187, 2163–2174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reglero, C.; Reglero, G. Precision Nutrition and Cancer Relapse Prevention: A Systematic Literature Review. Nutrients 2019, 11, 2799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermawan, A.; Putri, H. Current report of natural product development against breast cancer stem cells. Int. J. Biochem. Cell Biol. 2018, 104, 114–132. [Google Scholar] [CrossRef]
- Rodríguez-García, C.; Sánchez-Quesada, C.; J Gaforio, J. Dietary Flavonoids as Cancer Chemopreventive Agents: An Updated Review of Human Studies. Antioxidants (Basel) 2019, 8, 137. [Google Scholar] [CrossRef] [Green Version]
- Thilakarathna, S.H.; Rupasinghe, H.P. Flavonoid bioavailability and attempts for bioavailability enhancement. Nutrients 2013, 5, 3367–3387. [Google Scholar] [CrossRef]
- Hollman, P.C.H. Absorption, Bioavailability, and Metabolism of Flavonoids. Pharm. Biol. 2004, 42, 74–83. [Google Scholar] [CrossRef]
- Aherne, S.A.; O’Brien, N.M. Dietary flavonols: Chemistry, food content, and metabolism. Nutrition 2002, 18, 75–81. [Google Scholar] [CrossRef]
- Higdon, J.V.; Frei, B. Tea catechins and polyphenols: Health effects, metabolism, and antioxidant functions. Crit. Rev. Food Sci. Nutr. 2003, 43, 89–143. [Google Scholar] [CrossRef]
- Lopez-Lazaro, M. Distribution and biological activities of the flavonoid luteolin. Mini Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef]
- Kalaiselvan, V.; Kalaivani, M.; Vijayakumar, A.; Sureshkumar, K.; Venkateskumar, K. Current knowledge and future direction of research on soy isoflavones as a therapeutic agents. Pharmacogn. Rev. 2010, 8, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Del Bo’, C.; Bernardi, S.; Marino, M.; Porrini, M.; Tucci, M.; Guglielmetti, S.; Cherubini, A.; Carrieri, B.; Kirkup, B.; Kroon, P.; et al. Systematic Review on Polyphenol Intake and Health Outcomes: Is there Sufficient Evidence to Define a Health-Promoting Polyphenol-Rich Dietary Pattern? Nutrients 2019, 11, 1355. [Google Scholar]
- Hüser, S.; Guth, S.; Joost, H.G.; Soukup, S.T.; Köhrle, J.; Kreienbrock, L.; Diel, P.; Lachenmeier, D.W.; Eisenbrand, G.; Vollmer, G.; et al. Effects of isoflavones on breast tissue and the thyroid hormone system in humans: A comprehensive safety evaluation. Arch. Toxicol. 2018, 92, 2703–2748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.L.; Ho, S.C.; Mo, X.F.; Lin, F.Y.; Zhang, N.Q.; Luo, H.; Zhang, X.; Zhang, C.X. Association between flavonoids, flavonoid subclasses intake and breast cancer risk: A case-control study in China. Eur. J. Cancer Prev. 2019. [Google Scholar] [CrossRef] [PubMed]
- Touvier, M.; Druesne-Pecollo, N.; Kesse-Guyot, E.; Andreeva, V.A.; Fezeu, L.; Galan, P.; Hercberg, S.; Latino-Martel, P. Dual association between polyphenol intake and breast cancer risk according to alcohol consumption level: A prospective cohort study. Breast Cancer Res. Treat. 2013, 137, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Fink, B.N.; Steck, S.E.; Wolff, M.S.; Britton, J.A.; Kabat, G.C.; Schroeder, J.C.; Teitelbaum, S.L.; Neugut, A.I.; Gammon, M.D. Dietary flavonoid intake and breast cancer risk among women on Long Island. Am. J. Epidemiol. 2007, 165, 514–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fink, B.N.; Steck, S.E.; Wolff, M.S.; Britton, J.A.; Kabat, G.C.; Gaudet, M.M.; Abrahamson, P.E.; Bell, P.; Schroeder, J.C.; Teitelbaum, S.L.; et al. Dietary flavonoid intake and breast cancer survival among women on Long Island. Cancer Epidemiol. Biomarkers Prev. 2007, 16, 2285–2292. [Google Scholar] [CrossRef] [Green Version]
- Bosetti, C.; Spertini, L.; Parpinel, M.; Gnagnarella, P.; Lagiou, P.; Negri, E.; Franceschi, S.; Montella, M.; Peterson, J.; Dwyer, J.; et al. Flavonoids and breast cancer risk in Italy. Cancer Epidemiol Biomarkers Prev. 2005, 14, 805–808. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.Y.; Qin, L.Q. Soy isoflavones consumption and risk of breast cancer incidence or recurrence: A meta-analysis of prospective studies. Breast Cancer Res. Treat. 2011, 125, 315–323. [Google Scholar] [CrossRef]
- Xie, Q.; Chen, M.L.; Qin, Y.; Zhang, Q.Y.; Xu, H.X.; Zhou, Y.; Mi, M.T.; Zhu, J.D. Isoflavone consumption and risk of breast cancer: A dose-response meta-analysis of observational studies. Asia Pac. J. Clin. Nutr. 2013, 22, 118–127. [Google Scholar]
- Chen, M.; Rao, Y.; Zheng, Y.; Wei, S.; Li, Y.; Guo, T.; Yin, P. Association between soy isoflavone intake and breast cancer risk for pre- and post-menopausal women: A meta-analysis of epidemiological studies. PLoS ONE 2014, 9, e89288. [Google Scholar] [CrossRef] [Green Version]
- Keinan-Boker, L.; van Der Schouw, Y.T.; Grobbee, D.E.; Peeters, P.H. Dietary phytoestrogens and breast cancer risk. Am. J. Clin. Nutr. 2004, 79, 282–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verheus, M.; van Gils, C.H.; Keinan-Boker, L.; Grace, P.B.; Bingham, S.A.; Peeters, P.H. Plasma phytoestrogens and subsequent breast cancer risk. J. Clin. Oncol. 2007, 25, 648–655. [Google Scholar] [CrossRef]
- Ward, H.; Chapelais, G.; Kuhnle, G.G.; Luben, R.; Khaw, K.T.; Bingham, S.; European Prospective into Cancer-Norfolk cohort. Breast cancer risk in relation to urinary and serum biomarkers of phytoestrogen exposure in the European Prospective into Cancer-Norfolk cohort study. Breast Cancer Res. 2008, 10, R32. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Lv, J.; Guo, Y.; Bian, Z.; Gao, M.; Du, H.; Yang, L.; Chen, Y.; Zhang, X.; Wang, T.; et al. China Kadoorie Biobank Collaborative Group. Soy intake and breast cancer risk: A prospective study of 300,000 Chinese women and a dose-response meta-analysis. Eur. J. Epidemiol. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nachvak, S.M.; Moradi, S.; Anjom-Shoae, J.; Rahmani, J.; Nasiri, M.; Maleki, V.; Sadeghi, O. Soy, Soy Isoflavones, and Protein Intake in Relation to Mortality from All Causes, Cancers, and Cardiovascular Diseases: A Systematic Review and Dose-Response Meta-Analysis of Prospective Cohort Studies. J. Acad. Nutr. Diet. 2019, 119, 1483–1500.e17. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.K.; Sandin, S.; Ström, P.; Löf, M.; Adami, H.O.; Weiderpass, E. Prospective study of breast cancer in relation to coffee, tea and caffeine in Sweden. Int. J. Cancer 2015, 137, 1979–1989. [Google Scholar] [CrossRef]
- Lowcock, E.C.; Cotterchio, M.; Anderson, L.N.; Boucher, B.A.; El-Sohemy, A. High coffee intake, but not caffeine, is associated with reduced estrogen receptor negative and postmenopausal breast cancer risk with no effect modification by CYP1A2 genotype. Nutr. Cancer. 2013, 65, 398–409. [Google Scholar] [CrossRef]
- Boggs, D.A.; Palmer, J.R.; Stampfer, M.J.; Spiegelman, D.; Adams-Campbell, L.L.; Rosenberg, L. Tea and coffee intake in relation to risk of breast cancer in the Black Women’s Health Study. Cancer Causes Control 2010, 21, 1941–1948. [Google Scholar] [CrossRef] [Green Version]
- Bhoo Pathy, N.; Peeters, P.; van Gils, C.; Beulens, J.W.; van der Graaf, Y.; Bueno-de-Mesquita, B.; Bulgiba, A.; Uiterwaal, C.S. Coffee and tea intake and risk of breast cancer. Breast Cancer Res. Treat. 2010, 121, 461–467. [Google Scholar] [CrossRef]
- Tang, N.; Zhou, B.; Wang, B.; Yu, R. Coffee consumption and risk of breast cancer: A meta analysis. Am. J. Obstet. Gynecol. 2009, 200, 290.e1-9. [Google Scholar] [CrossRef] [PubMed]
- Nakachi, K.; Matsuyama, S.; Miyake, S.; Suganuma. M.; Imai. K. Preventive effects of drinking green tea on cancer and cardiovascular disease: Epidemiological evidence for multiple targeting prevention. Biofactors 2000, 13, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Zhu, L.; Wang, K.; Yan, Y.; He, J.; Ren, Y. Green tea consumption and risk of breast cancer: A systematic review and updated meta-analysis of case-control studies. Medicine (Baltimore) 2019, 98, e16147. [Google Scholar] [CrossRef] [PubMed]
- Pasculli, B.; Barbano, R.; Parrella, P. Epigenetics of breast cancer: Biology and clinical implication in the era of precision medicine. Semin. Cancer Biol. 2018, 51, 22–35. [Google Scholar] [CrossRef]
- Teschendorff, A.E.; Gao, Y.; Jones, A.; Ruebner, M.; Beckmann, M.W.; Wachter, D.L.; Fasching, P.A.; Widschwendter, M. DNA methylation outliers in normal breast tissue identify field defects that are enriched in cancer. Nat. Commun. 2016, 7, 10478. [Google Scholar] [CrossRef]
- DeVaux, R.S.; Herschkowitz, J.I. Beyond DNA: The Role of Epigenetics in the Premalignant Progression of Breast Cancer. J. Mammary Gland Biol. Neoplasia 2018, 23, 223–235. [Google Scholar] [CrossRef] [Green Version]
- Sturgeon, S.R.; Balasubramanian, R.; Schairer, C.; Muss, H.B.; Ziegler, R.G.; Arcaro, K.F. Detection of promoter methylation of tumor suppressor genes in serum DNA of breast cancer cases and benign breast disease controls. Epigenetics 2012, 7, 1258–1267. [Google Scholar] [CrossRef] [Green Version]
- Vos, S.; Moelans, C.B.; van Diest, P.J. BRCA promoter methylation in sporadic versus BRCA germline mutation-related breast cancers. Breast Cancer Res. 2017, 19, 64. [Google Scholar] [CrossRef] [Green Version]
- Busch, C.; Burkard, M.; Leischner, C.; Lauer, U.M.; Frank, J.; Venturelli, S. Epigenetic activities of flavonoids in the prevention and treatment of cancer. Clin. Epigenetics 2015, 7, 64. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.; Eom, G.H. HDAC and HDAC Inhibitor: From Cancer to Cardiovascular Diseases. Chonnam Med. J. 2016, 52, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Eom, G.H.; Kook, H. Posttranslational modifications of histone deacetylases: Implications for cardiovascular diseases. Pharmacol. Ther. 2014, 143, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Gatla, H.R.; Zou, Y.; Uddin, M.M.; Singha, B.; Bu, P.; Vancura, A.; Vancurova, I. Histone, Deacetylase (HDAC) Inhibition Induces IκB Kinase (IKK)-dependent Interleukin-8/CXCL8 Expression in Ovarian Cancer Cells. J. Biol. Chem. 2017, 292, 5043–5054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vancurova, I.; Uddin, M.M.; Zou, Y.; Vancura, A. Combination Therapies Targeting HDAC and IKK in Solid Tumors. Trends Pharmacol. Sci. 2018, 39, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Gianfredi, V.; Nucci, D.; Abalsamo, A.; Acito, M.; Villarini, M.; Moretti, M.; Realdon, S. Green Tea Consumption and Risk of Breast Cancer and Recurrence—A Systematic Review and Meta-Analysis of Observational Studies. Nutrients 2018, 10, 1886. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, M.; Inoue, M.; Sasazuki, S.; Sawada, N.; Yamaji, T.; Shimazu, T.; Willett, W.C.; Tsugane, S.; Japan Public Health Center-Based Prospective Study Group. Green tea drinking and subsequent risk of breast cancer in a population-based cohort of Japanese women. Breast Cancer Res. 2010, 12, R88. [Google Scholar] [CrossRef] [Green Version]
- Berletch, J.B.; Liu, C.; Love, W.K.; Andrews, L.G.; Katiyar, S.K.; Tollefsbol, T.O. Epigenetic and genetic mechanisms contribute to telomerase inhibition by EGCG. J. Cell Biochem. 2008, 103, 509–519. [Google Scholar] [CrossRef] [Green Version]
- Meeran, S.M.; Patel, S.N.; Chan, T.H.; Tollefsbol, T.O. A novel prodrug of epigallocatechin-3-gallate: Differential epigenetic hTERT repression in human breast cancer cells. Cancer Prev. Res. (Phila) 2011, 4, 1243–1254. [Google Scholar] [CrossRef] [Green Version]
- Mirza, S.; Sharma, G.; Parshad, R.; Gupta, S.D.; Pandya, P.; Ralhan, R. Expression of DNA methyltransferases in breast cancer patients and to analyze the effect of natural compounds on DNA methyltransferases and associated proteins. J. Breast Cancer. 2013, 16, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.C.; Lee, Y.C.; Li, L.H.; Cheng, C.J.; Yang, R.B. Tumor suppressor SCUBE2 inhibits breast-cancer cell migration and invasion through the reversal of epithelial-mesenchymal transition. J. Cell Sci. 2014, 127, 85–100. [Google Scholar] [CrossRef] [Green Version]
- Sheng, J.; Shi, W.; Guo, H.; Long, W.; Wang, Y.; Qi, J.; Liu, J.; Xu, Y. The Inhibitory Effect of (-)-Epigallocatechin-3-Gallate on Breast Cancer Progression via Reducing SCUBE2 Methylation and DNMT Activity. Molecules 2019, 24, 2899. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Buckhaults, P.; Cui, X.; Tollefsbol, T.O. Combinatorial epigenetic mechanisms and efficacy of early breast cancer inhibition by nutritive botanicals. Epigenomics 2016, 8, 1019–1037. [Google Scholar] [CrossRef] [Green Version]
- Lewis, K.A.; Jordan, H.R.; Tollefsbol, T.O. Effects of SAHA and EGCG on Growth Potentiation of Triple-Negative Breast Cancer Cells. Cancers (Basel) 2018, 11, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Yuan, Y.Y.; Meeran, S.M.; Tollefsbol, T.O. Synergistic epigenetic reactivation of estrogen receptor-α (ERα) by combined green tea polyphenol and histone deacetylase inhibitor in ERα-negative breast cancer cells. Mol. Cancer. 2010, 9, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Meeran, S.M.; Tollefsbol, T.O. Combinatorial bioactive botanicals re-sensitize tamoxifen treatment in ER-negative breast cancer via epigenetic reactivation of ERα expression. Sci. Rep. 2017, 7, 9345. [Google Scholar] [CrossRef] [Green Version]
- Bhagwat, S.; Haytowitz, D.B.; Holden, J.M. (Ret.). 2014, US DA Database for the Flavonoid Content of Selected Foods, Release 3.1. U.S. Department of Agriculture, Agricultural Research Service. Nutrient Data Laboratory Home Page. Available online: http://www.ars.usda.gov/nutrientdata/flav (accessed on 12 December 2019).
- Mirjalili, M.H.; Moyano, E.; Bonfill, M.; Cusido, R.M.; Palazón, J. Steroidal lactones from Withania somnifera, an ancient plant for novel medicine. Molecules 2009, 14, 2373–2393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, R. Therapeutic effects of guggul and its constituent guggulsterone:cardiovascular benefits. Cardiovasc Drug Rev. 2007, 25, 375–390. [Google Scholar] [PubMed]
- Janle, E.M.; Morré, D.M.; Morré, D.J.; Zhou, Q.; Zhu, Y. Pharmacokinetics of green tea catechins in extract and sustained-release preparations. J. Diet. Suppl. 2008, 5, 248–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.S. Inhibition of carcinogenesis by tea. Nat. Clin. Proc. Cardiovasc. Med. 1997, 389, 134–135. [Google Scholar] [CrossRef]
- Nakagawa, K.; Okuda, S.; Miyazawa, T. Dose-dependent incorporation of tea catechins, (-)-epigallocatechin-3-gallate and (-)-epigallocatechin, into human plasma. Biosci. Biotechnol. Biochem. 1997, 61, 1981–1985. [Google Scholar] [CrossRef]
- Yang, C.S.; Chen, L.; Lee, M.J.; Balentine, D.; Kuo, M.C.; Schantz, S.P. Blood and urine levels of tea catechins after ingestion of different amounts of green tea by human volunteers. Cancer Epidemiol. Biomarkers Prev. 1998, 7, 351–354. [Google Scholar]
- Iino, C.; Shimoyama, T.; Iino, K.; Yokoyama, Y.; Chinda, D.; Sakuraba, H.; Fukuda, S.; Nakaji, S. Daidzein Intake Is Associated with Equol Producing Status through an Increase in the Intestinal Bacteria Responsible for Equol Production. Nutrients 2019, 11, 433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, K.B.; Atkinson, C.; Frankenfeld, C.L.; Jokela, T.; Wähälä, K.; Thomas, W.K.; Lampe, J.W. Prevalence of daidzein-metabolizing phenotypes differs between Caucasian and Korean American women and girls. J. Nutr. 2006, 136, 1347–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziaei, S.; Halaby, R. Dietary Isoflavones and Breast Cancer Risk. Medicines (Basel) 2017, 4, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trock, B.J.; Hilakivi-Clarke, L.; Clarke, R. Meta-analysis of soy intake and breast cancer risk. J. Natl. Cancer Inst. 2006, 98, 459–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, X.O.; Zheng, Y.; Cai, H.; Gu, K.; Chen, Z.; Zheng, W.; Lu, W. Soy food intake and breast cancer survival. JAMA 2009, 302, 2437–2443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messina, M.; Hilakivi-Clarke, L. Early intake appears to be the key to the proposed protective effects of soy intake against breast cancer. Nutr. Cancer. 2009, 61, 792–798. [Google Scholar] [CrossRef] [PubMed]
- “Soy Isoflavones” Linus Pauling Institute. Available online: https://lpi.oregonstate.edu/mic/dietary-factors/phytochemicals/soy-isoflavones#source (accessed on 3 March 2020).
- Khan, S.A.; Chatterton, R.T.; Michel, N.; Bryk, M.; Lee, O.; Ivancic, D.; Heinz, R.; Zalles, C.M.; Helenowski, I.B.; Jovanovic, B.D.; et al. Soy isoflavone supplementation for breast cancer risk reduction: A randomized phase II trial. Cancer Prev Res. (Phila) 2012, 5, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Meeran, S.M.; Patel, S.N.; Chen, H.; Hardy, T.M.; Tollefsbol, T.O. Epigenetic reactivation of estrogen receptor-α (ERα) by genistein enhances hormonal therapy sensitivity in ERα-negative breast cancer. Mol. Cancer 2013, 12, 9. [Google Scholar] [CrossRef] [Green Version]
- Bosviel, R.; Dumollard, E.; Déchelotte, P.; Bignon, Y.J.; Bernard-Gallon, D. Can soy phytoestrogens decrease DNA methylation in BRCA1 and BRCA2 oncosuppressor genes in breast cancer? OMICS 2012, 16, 235–244. [Google Scholar] [CrossRef]
- Romagnolo, D.F.; Donovan, M.G.; Papoutsis, A.J.; Doetschman, T.C.; Selmin, O.I. Genistein Prevents BRCA1 CpG Methylation and Proliferation in Human Breast Cancer Cells with Activated Aromatic Hydrocarbon Receptor. Curr. Dev. Nutr. 2017, 1, e000562. [Google Scholar] [CrossRef] [Green Version]
- Donovan, M.G.; Selmin, O.I.; Doetschman, T.C.; Romagnolo, D.F. Epigenetic Activation of BRCA1 by Genistein In Vivo and Triple Negative Breast Cancer Cells Linked to Antagonism toward Aryl Hydrocarbon Receptor. Nutrients 2019, 11, 2559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hockings, J.K.; Thorne, P.A.; Kemp, M.Q.; Morgan, S.S.; Selmin, O.; Romagnolo, D.F. The ligand status of the aromatic hydrocarbon receptor modulates transcriptional activation of BRCA-1 promoter by estrogen. Cancer Res. 2006, 66, 2224–2232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jadhav, R.R.; Santucci-Pereira, J.; Wang, Y.V.; Liu, J.; Nguyen, T.D.; Wang, J.; Jenkins, S.; Russo, J.; Huang, T.H.; Jin, V.X.; et al. DNA Methylation Targets Influenced by Bisphenol A and/or Genistein Are Associated with Survival Outcomes in Breast Cancer Patients. Genes (Basel) 2017, 8, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dagdemir, A.; Durif, J.; Ngollo, M.; Bignon, Y.J.; Bernard-Gallon, D. Histone lysine trimethylation or acetylation can be modulated by phytoestrogen, estrogen or anti-HDAC in breast cancer cell lines. Epigenomics 2013, 5, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Paul, B.; Li, Y.; Tollefsbol, T.O. The Effects of Combinatorial Genistein and Sulforaphane in Breast Tumor Inhibition: Role in Epigenetic Regulation. Int. J. Mol. Sci. 2018, 19, 1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassidy, A.; Brown, J.E.; Hawdon, A.; Faughnan, M.S.; King, L.J.; Millward, J.; Zimmer-Nechemias, L.; Wolfe, B.; Setchell, K.D. Factors affecting the bioavailabilityof soy isoflavones in humans after ingestion of physiologically relevant levelsfrom different soy foods. J. Nutr. 2006, 136, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Anupongsanugool, E.; Teekachunhatean, S.; Rojanasthien, N.; Pongsatha, S.; Sangdee, C. Pharmacokinetics of isoflavones, daidzein and genistein, after ingestion of soybeverage compared with soy extract capsules in postmenopausal Thai women. BMC Clin. Pharmacol. 2005, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Setchell, K.D.; Brown, N.M.; Desai, P.; Zimmer-Nechemias, L.; Wolfe, B.E.; Brashear, W.T.; Kirschner, A.S.; Cassidy, A.; Heubi, J.E. Bioavailability of pure isoflavones in healthy humans and analysis of commercial soy isoflavone supplements. J. Nutr. 2001, 131, 1362S–1375S. [Google Scholar] [CrossRef] [Green Version]
- Salehi, B.; Mishra, A.P.; Nigam, M.; Sener, B.; Kilic, M.; Sharifi-Rad, M.; Fokou, P.V.T.; Martins, N.; Sharifi-Rad, J. Resveratrol: A Double-Edged Sword in Health Benefits. Biomedicines 2018, 6, 91. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo-Torres, E.; Hernández-Oliveras, A.; Meneses-Morales, I.; Rodríguez, G.; Fuentes-García, G.; Zarain-Herzberg, Á. Resveratrol up-regulates ATP2A3 gene expression in breast cancer cell lines through epigenetic mechanisms. Int. J. Biochem. Cell Biol. 2019, 113, 37–47. [Google Scholar] [CrossRef]
- Contreras-Leal, E.; Hernández-Oliveras, A.; Flores-Peredo, L.; Zarain-Herzberg, Á.; Santiago-García, J. Histone deacetylase inhibitors promote the expression of ATP2A3 gene in breast cancer cell lines. Mol. Carcinog. 2016, 55, 1477–1485. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Tollefsbol, T.O. Combinational Proanthocyanidins and Resveratrol Synergistically Inhibit Human Breast Cancer Cells and Impact Epigenetic⁻Mediating Machinery. Int. J. Mol. Sci. 2018, 19, 2204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papoutsis, A.J.; Selmin, O.I.; Borg, J.L.; Romagnolo, D.F. Gestational exposure to the AhR agonist 2,3,7,8-tetrachlorodibenzo-p-dioxin induces BRCA-1 promoter hypermethylation and reduces BRCA-1 expression in mammary tissue of rat offspring: Preventive effects of resveratrol. Mol. Carcinog. 2015, 54, 261–269. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, M.R.; Nabavi, S.M.; Braidy, N.; Setzer, W.N.; Ahmed, T.; Nabavi, S.F. Quercetin and the mitochondria: A mechanistic view. Biotechnol. Adv. 2016, 5, 532–549. [Google Scholar] [CrossRef]
- Srinivasan, A.; Thangavel, C.; Liu, Y.; Shoyele, S.; Den, R.B.; Selvakumar, P.; Lakshmikuttyamma, A. Quercetin regulates β-catenin signaling and reduces the migration of triple negative breast cancer. Mol. Carcinog. 2016, 55, 743–756. [Google Scholar] [CrossRef]
- Shang, S.; Hua, F.; Hu, Z.W. The regulation of β-catenin activity and function in cancer: Therapeutic opportunities. Oncotarget 2017, 8, 33972–33989. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.H.; An, J.Y.; Kwon, Y.T.; Rhee, J.G.; Lee, Y.J. Effects of low dose quercetin: Cancer cell-specific inhibition of cell cycle progression. J. Cell Biochem. 2009, 106, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Kundur, S.; Prayag, A.; Selvakumar, P.; Nguyen, H.; McKee, L.; Cruz, C.; Srinivasan, A.; Shoyele, S.; Lakshmikuttyamma, A. Synergistic anticancer action of quercetin and curcumin against triple-negative breast cancer cell lines. J. Cell Physiol. 2019, 234, 11103–11108. [Google Scholar] [CrossRef]
- Ferry, D.R.; Smith, A.; Malkhandi, J.; Fyfe, D.W.; deTakats, P.G.; Anderson, D.; Baker, J.; Kerr, D.J. Phase I clinical trial of the flavonoid quercetin: Pharmacokinetics and evidence for in vivo tyrosine kinase inhibition. Clin. Cancer Res. 1996, 2, 659–668. [Google Scholar]
- Egert, S.; Wolffram, S.; Bosy-Westphal, A.; Boesch-Saadatmandi, C.; Wagner, A.E.; Frank, J.; Rimbach, G.; Mueller, M.J. Daily quercetin supplementation dose-dependentlyincreases plasma quercetin concentrations in healthy humans. J. Nutr. 2008, 138, 1615–1621. [Google Scholar] [CrossRef] [Green Version]
- Jin, F.; Nieman, D.C.; Shanely, R.A.; Knab, A.M.; Austin, M.D.; Sha, W. The variable plasmaquercetin response to 12-week quercetin supplementation in humans. Eur. J. Clin. Nutr. 2010, 64, 692–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zander, S.A.; Kersbergen, A.; Sol, W.; Gonggrijp, M.; van de Wetering, K.; Jonkers, J.; Borst, P.; Rottenberg, S. Lack of ABCG2 shortens latency of BRCA1-deficient mammary tumors and this is not affected by genistein or resveratrol. Cancer Prev Res. (Phila) 2012, 8, 1053–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
| Dietary Source | mg/100 g Source |
|---|---|
| Green tea, brewed | 70.20 |
| Oolong tea, brewed | 34.48 |
| Green tea, brewed (decaffeinated) | 26.05 |
| Black tea, brewed | 9.36 |
| Pecannut | 2.30 |
| Fuji Apple, with skin | 1.93 |
| Hazelnut | 1.06 |
| Black tea, brewed (decaffeinated) | 1.01 |
| Cranberry | 0.97 |
| Blackberry | 0.68 |
| Raspberry | 0.54 |
| Plum | 0.48 |
| Red Delicious Apple, without skin | 0.46 |
| Pistachio nuts | 0.40 |
| Granny Smith Apple, with skin | 0.24 |
| Golden Delicious Apple, with skin | 0.19 |
| Avocado | 0.15 |
| Red Delicious Apple, with skin | 0.13 |
| Gala Apple, with skin | 0.11 |
| Strawberry | 0.11 |
| Dietary Source | Serving | Genistein (mg) | Daidzein (mg) |
|---|---|---|---|
| Soybeans, mature seeds (boiled) | ½ cup | 26.9 | 26.5 |
| Soy protein concentrate, aqueous washed | 3.5 ounces | 52.8 | 38.2 |
| Soy protein concentrate, alcohol washed | 3.5 ounces | 5.8 | 5.3 |
| Soybeans, dry roasted | 1 ounce | 21.2 | 17.4 |
| Soybeans, green, boiled (edamame) | ½ cup | 6.3 | 6.7 |
| Miso | ½ cup | 32 | 22.6 |
| Soy milk, low-fat | 1 cup | 3.7 | 2.4 |
| Tofu, soft | 3 ounces | 10.1 | 8.1 |
| Tofu, yogurt | ½ cup | 12.3 | 7.5 |
| Tempeh | 3 ounces | 30.7 | 19.3 |
| Tempeh, cooked | 3 ounces | 18 | 11.1 |
| Soy burger, unprepared | 1 patty | 3.5 | 1.6 |
| Soy sausage | 3 links | 6.9 | 3.3 |
| Soy cheese, cheddar | 1 ounces | 0.6 | 0.5 |
| Dietary Source | mg/100 g Source | Dietary Source | mg/100 g Source |
|---|---|---|---|
| Elderberry juice | 108.16 | Mustard greens, raw | 8.80 |
| Radish leaves, raw | 70.37 | Mizuna (Japanese mustard) | 8.55 |
| Chokeberry juice | 68.17 | Arugula, raw | 7.92 |
| Wild rocket, raw | 66.19 | Blueberry | 7.67 |
| Cilantro leaves, raw | 52.90 | Red leaf lettuce, raw | 7.61 |
| Yellow wax hot pepper, raw | 50.63 | Red swiss chard, raw | 7.50 |
| Juniper berry, ripe | 46.61 | Rowanberry, raw | 7.40 |
| Green juniper berry, unripe | 42.81 | Chokeberry juice | 6.49 |
| Red onion, raw | 39.21 | Fig, raw | 5.47 |
| Radicchio, raw | 31.51 | Crowberry, raw | 5.45 |
| Watercress, raw | 29.99 | Chives, raw | 4.77 |
| Hartwort leaves | 29.30 | Acerola, west Indian cherry | 4.74 |
| Ancho pepper | 27.60 | Blueberry, frozen | 4.64 |
| White onion, pan fried | 26.90 | Cranberry, dried | 4.50 |
| Corn poppy leaves | 26.30 | Kale, canned | 4.50 |
| Onion, cooked | 24.36 | Bayberry, raw | 4.36 |
| Hawthorn leaves, raw | 24.10 | Brussels sprout, cooked | 4.33 |
| Nalta jute, raw | 23.53 | Green leaf lettuce, raw | 4.16 |
| Currant | 22.85 | Red Delicious Apple, with skin | 3.86 |
| Okra, raw | 20.97 | Gala Apple, with skin | 3.80 |
| Apple, skin only | 19.36 | Golden Delicious Apple, with skin | 3.69 |
| Wild bog whortleberry, frozen | 17.70 | Blackberry, raw | 3.58 |
| Sour cherry, powder | 17.44 | Bay leaves | 4.33 |
| Sweet potato leaves, raw | 16.94 | Green leaf lettuce, raw | 3.19 |
| Juneberry or Saskatoon berry | 16.64 | Concord grape | 3.11 |
| Cranberry juice | 16.41 | Bilberry, raw | 3.04 |
| Buckwheat | 15.38 | Yellow snap bean, cooked | 3.03 |
| Asparagus, cooked | 15.16 | Granny Smith Apple, with skin | 2.54 |
| Cranberry, raw | 14.84 | Cranberry sauce, canned | 2.40 |
| Asparagus, raw | 13.98 | Fuji Apple, with skin | 2.35 |
| Goji berry (wolfberry), dried | 13.60 | Broccoli raab, raw | 2.25 |
| Lingonberry (cowberry) | 13.30 | Purple plum, raw | 2.19 |
| Spring onion leaves | 12.60 | Tomato juice, canned | 1.19 |
| Black diamond plum, with peel | 12.45 | Red grape | 1.04 |
| Cranberry bush berry, raw | 10.73 | Watercress, steamed | 0.63 |
| Sweet potato leaves, cooked | 9.84 | Golden Delicious Apple, without skin | 0.51 |
| Arctic brambleberry | 9.10 | Red Delicious Apple, without skin | 0.41 |
| Flavonoid | Epigenetic Modification | Breast Cancer Cell Line | Regulated Genes | Reference |
|---|---|---|---|---|
| EGCG | ↓ Promotermethylation ↓ H3Lys9 Acetylation | MCF-7 | ↓ hTERT | Berletch et al., 2008 [58] |
| EGCG | ↓ DNMT ↓ HAT | MCF-7 & MDA-MB-231 | ↓ hTERT | Meeran et al., 2011 [59] |
| EGCG | ↓ Promoter methylation ↓ DNMT | MDA-MB-231 | Signal peptide-CUB-EGF domain-containing protein 2 (SCUBE2) | Lin et al., 2014 [61] Sheng et al., 2019 [62] |
| EGCG & SAHA | ↓ DNMT ↑ Histone Acetylation | MCF-7, MDA-MB-231 & MDA-MB-157 | ↑ ERα, p27, & E-cadherin | Lewis et al., 2018 [64] |
| EGCG & Sulforaphane | ↓ DNMT& HDAC | HMECs MDA-MB-231 & MDA-MB-157 | ↑ DCBLD2, ↓ SPET9↑ ERα | Li et al., 2016 [63] Li et al., 2017 [66] |
| EGCG & Trichostatin A | ↑ Histone Acetylation ↑ H3K4Me2 ↓ H3K9 | MDA-MB-231 | ↑ ERα | Liet al., 2010 [65] |
| Genistein & Trichostatin A | ↑ Histone Acetylation ↓ DNMT | MDA-MB-231 | ↑ ERα | Li et al., 2013 [82] |
| Genistein and Daidzein | ↓ Promoter methylation ↓ MeCP2 | MDA-MB-231 | ↑ BRCA1 & BRCA2 | Bosviel et al., 2012 [83] |
| Genistein | ↓ DNMT1 | UACC-3199 HCC38 | ↑ BRCA1 & ERα | Romagnolo et al., 2017 [84] Donovan et al., 2019 [85] |
| Genistein | ↓ H3K27Me3 & H3K9Me3, ↑ H4K8Ac and H3K4Ac | MDA-MB-231 | ↑ EZH2, BRCA1, ERα, ERβ, SRC3 and P300 | Dagdemir et al., 2013 [88] |
| Genistein and Sulforaphane | ↓ HDAC & HMT | MCF-7 & MDA-MB-231 | ↓ KLF4 and hTERT | Paul et al., 2018 [89] |
| Resveratrol | ↑ H3K27Ac ↓ DNMT, MeCP2 & MBD2 | MDA-MB-231 | ↑ ATP2A3 | Izquierdo-Torres et al., 2019 [94] |
| Resveratrol | ↓ DNMT1 | ↑ BRCA1 | Papoutsis et al., 2015 [97] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selvakumar, P.; Badgeley, A.; Murphy, P.; Anwar, H.; Sharma, U.; Lawrence, K.; Lakshmikuttyamma, A. Flavonoids and Other Polyphenols Act as Epigenetic Modifiers in Breast Cancer. Nutrients 2020, 12, 761. https://doi.org/10.3390/nu12030761
Selvakumar P, Badgeley A, Murphy P, Anwar H, Sharma U, Lawrence K, Lakshmikuttyamma A. Flavonoids and Other Polyphenols Act as Epigenetic Modifiers in Breast Cancer. Nutrients. 2020; 12(3):761. https://doi.org/10.3390/nu12030761
Chicago/Turabian StyleSelvakumar, Priyanga, Aja Badgeley, Paige Murphy, Hina Anwar, Urvashi Sharma, Katharine Lawrence, and Ashakumary Lakshmikuttyamma. 2020. "Flavonoids and Other Polyphenols Act as Epigenetic Modifiers in Breast Cancer" Nutrients 12, no. 3: 761. https://doi.org/10.3390/nu12030761