Antifatigue Activity and Exercise Performance of Phenolic-Rich Extracts from Calendula officinalis, Ribes nigrum, and Vaccinium myrtillus

 ,
, 
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extraction and Isolation
2.3. Determination of Total Phenolics
2.4. Antioxidant Activity
2.5. Animals and Experimental Designs
2.6. Exercise Endurance Performance Test
2.7. Forelimb Grip Strength
2.8. Determination of Fatigue-Associated Biochemical Variables
2.9. Clinical Biochemical Profiles
2.10. Body Composition and Glycogen Content Analysis
2.11. Histopathology
2.12. Statistical Analysis
3. Results
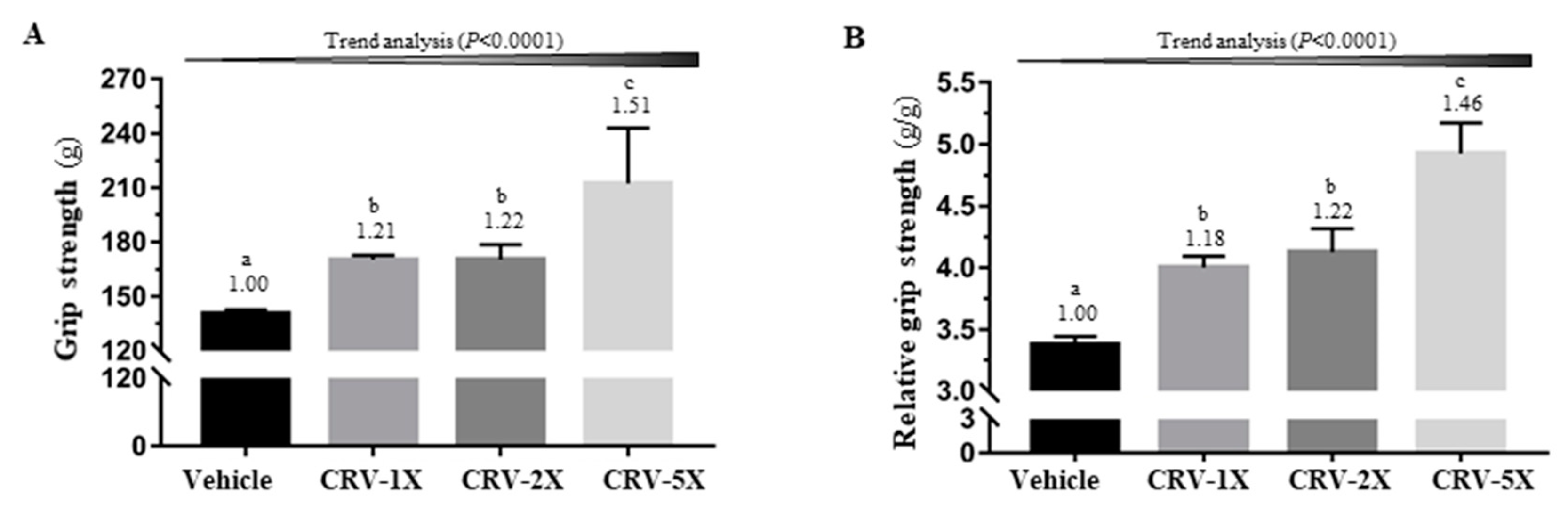
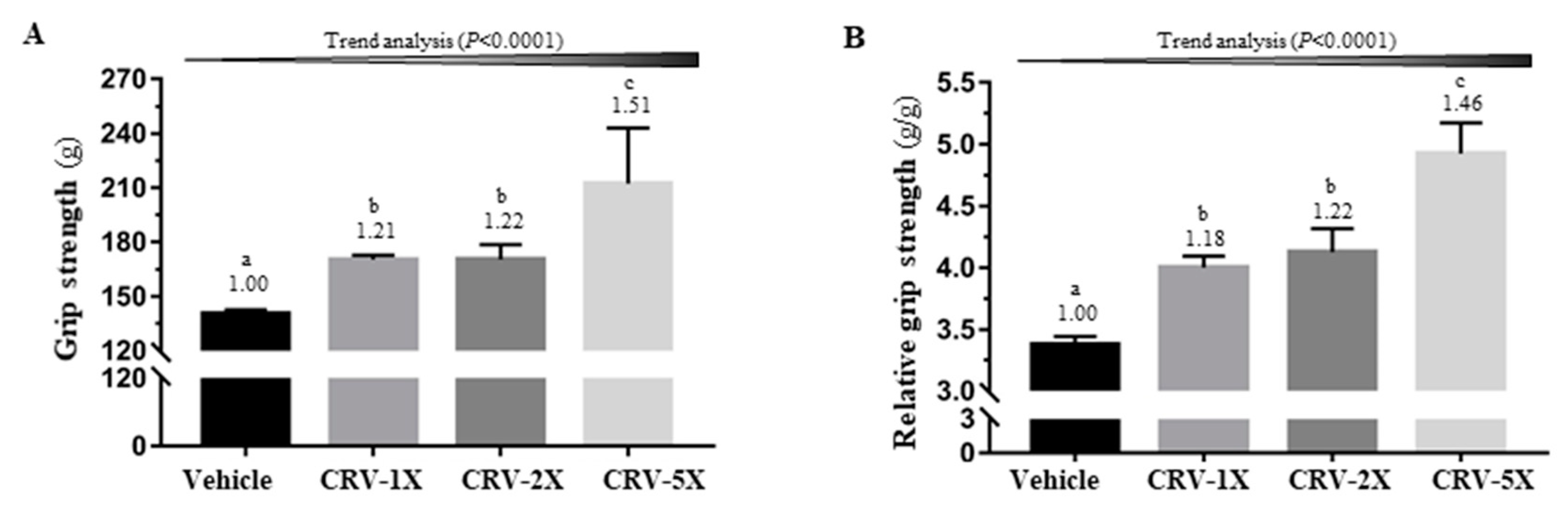
3.1. Effect of CRV Supplementation on Grip Strength
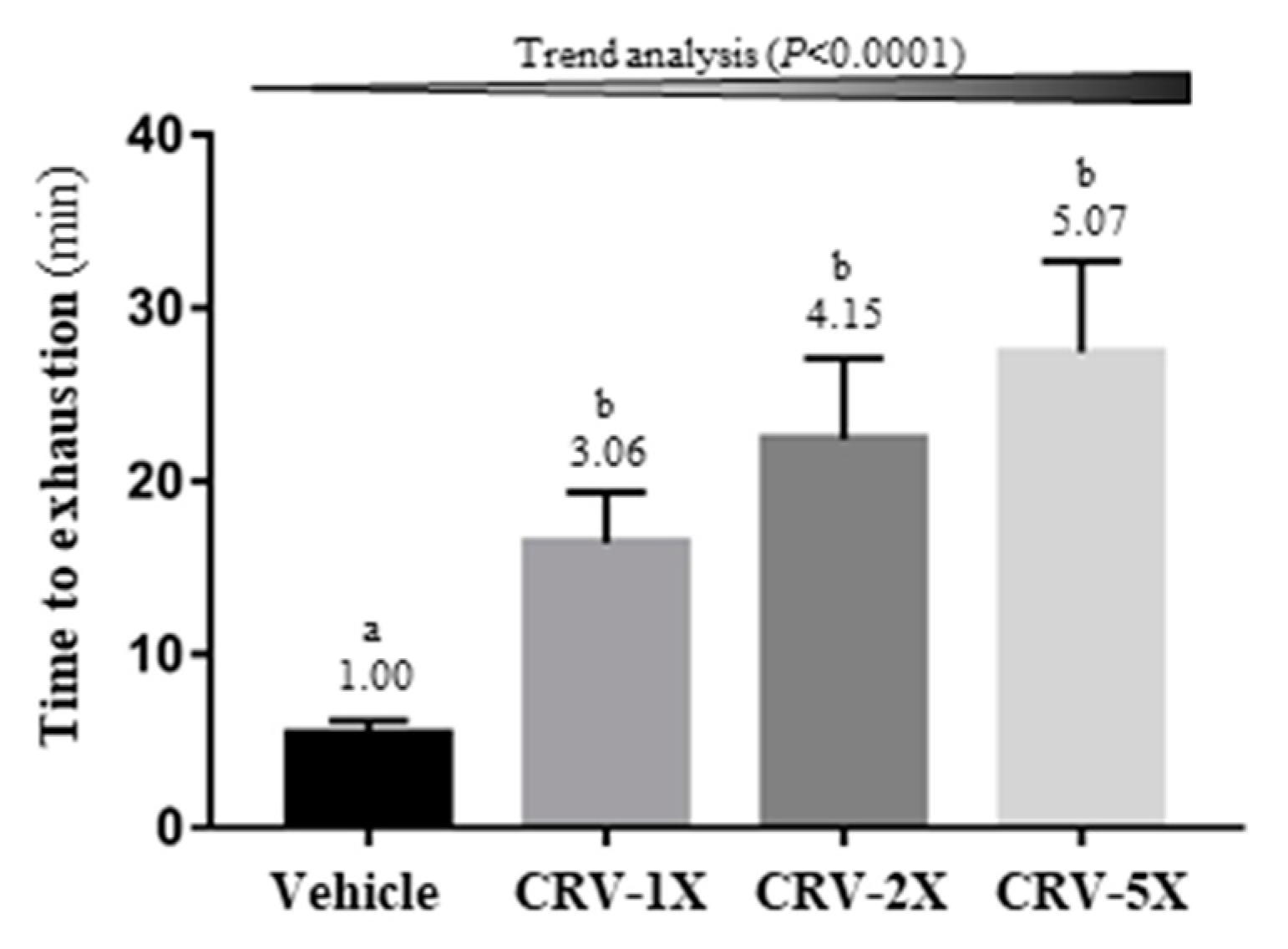
3.2. Effect of CRV Supplementation on Endurance Capacity
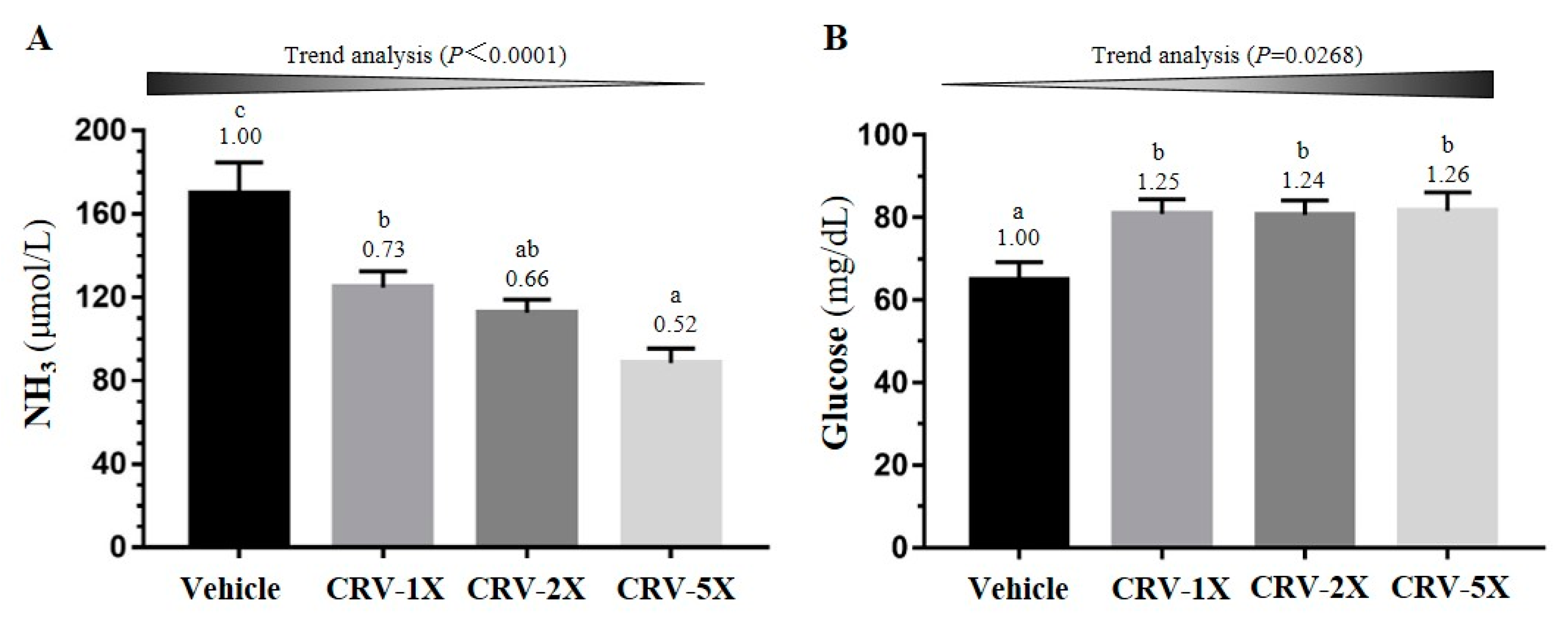
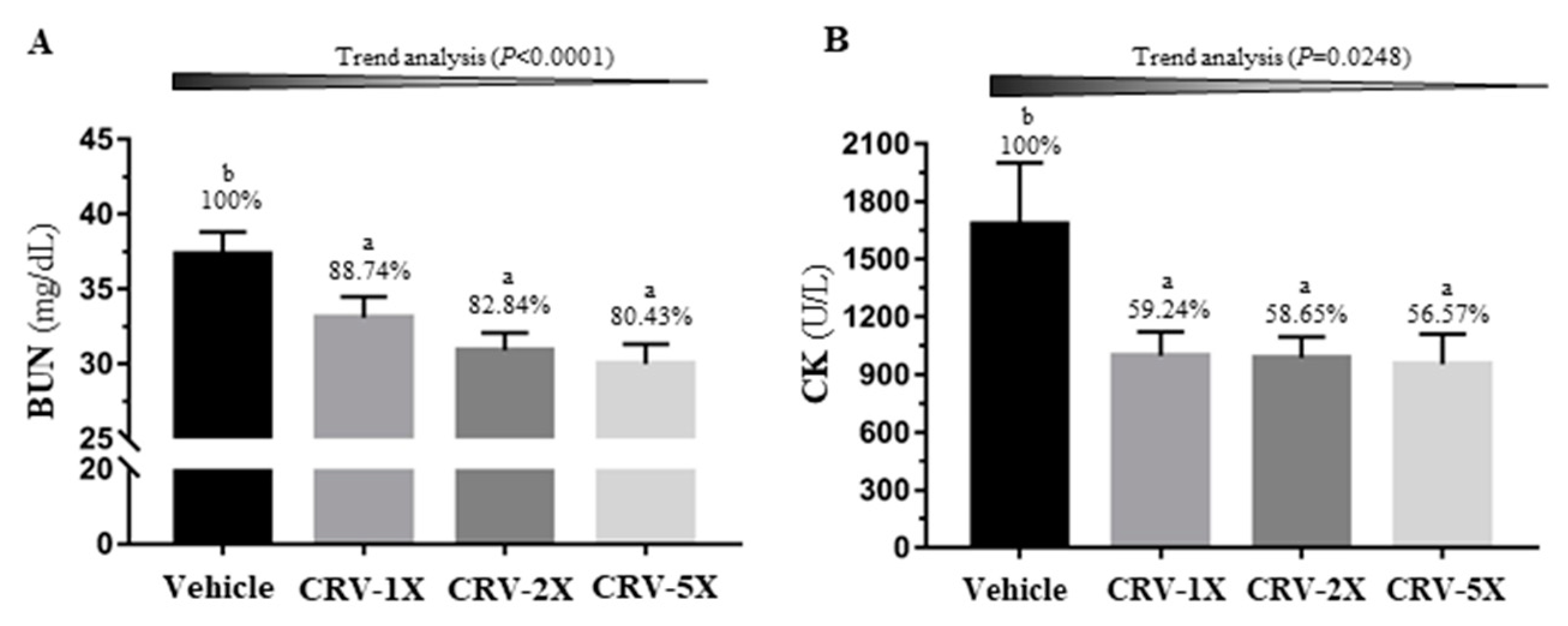
3.3. Effect of CRV Supplementation on Exercise-Related Biochemical Indexes after Exercise Challenge
3.4. Effect of CRV Supplementation on Glycogen Content
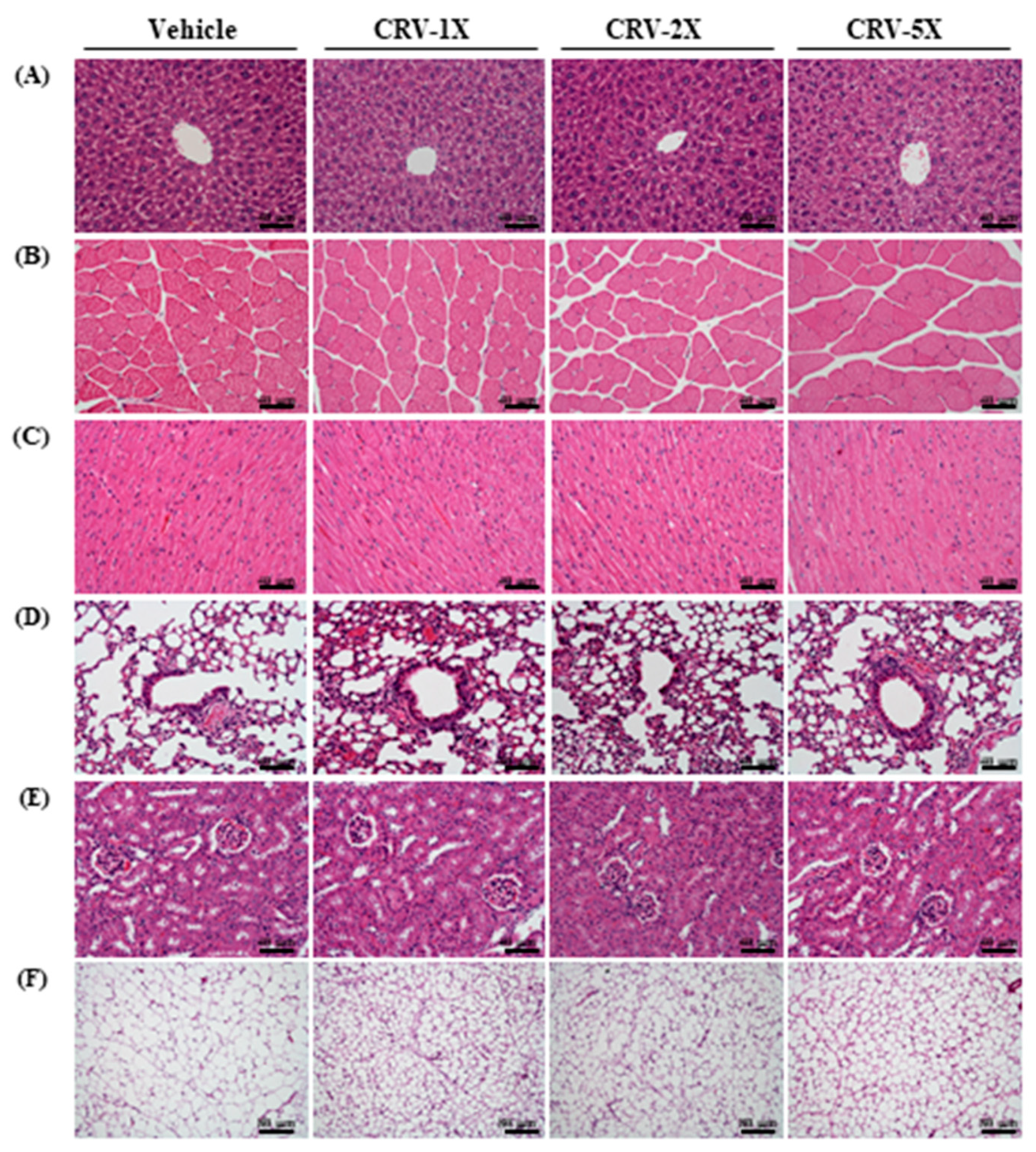
3.5. Subacute Oral Toxicity Evaluation after CRV Supplementation
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Belluardo, N.; Westerblad, H.; Mudó, G.; Casabona, A.; Bruton, J.; Caniglia, G.; Pastoris, O.; Grassi, F.; Ibáñez, C.F. Neuromuscular junction disassembly and muscle fatigue in mice lacking neurotrophin-4. Mol. Cell Neurosci. 2001, 18, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Yu, K.W.; Kang, D.H.; Suh, H.J. Anti-stress and anti-fatigue effect of fermented rice bran. Phytother. Res. 2002, 16, 700–702. [Google Scholar] [CrossRef] [PubMed]
- Preethi, K.C.; Kuttan, R. Hepato and reno protective action of Calendula officinalis L. flower extract. Indian J. Exp. Biol. 2009, 47, 163–168. [Google Scholar] [PubMed]
- Koeppen, B.H.; Herrmann, K. Flavonoid glycosides and hydroxycinnamic acid esters of blackcurrants (Ribes nigrum). Phenolics of fruits 9. Z. Lebensm. Unters. 1977, 164, 263–268. [Google Scholar] [CrossRef]
- Macheix, J.J.; Fleuriet, A.; Billot, J. Fruit Phenolics; CRC Press: Boca Raton, FL, USA, 1990; pp. 1–103. [Google Scholar]
- Schuster, B.; Herrmann, K. Hydroxybenzoic and hydroxycinnamic acid derivatives in soft fruits. Phytochemistry 1985, 24, 2761–2764. [Google Scholar] [CrossRef]
- Anttonen, M.J.; Karjalainen, R.O. High-performance liquid chromatography analysis of black currant (Ribes nigrum L.) fruit phenolics grown either conventionally or organically. J. Agric. Food Chem. 2006, 54, 7530–7538. [Google Scholar] [CrossRef]
- Katsube, N.; Iwashita, K.; Tsushida, T.; Yamaki, K.; Kobori, M. Induction of apoptosis in cancer cells by Bilberry (Vaccinium myrtillus) and the anthocyanins. J. Agric. Food Chem. 2003, 51, 68–75. [Google Scholar] [CrossRef]
- Faria, A.; Oliveira, J.; Neves, P.; Gameiro, P.; Santos-Buelga, C.; de Freitas, V.; Mateus, N. Antioxidant properties of prepared blueberry (Vaccinium myrtillus) extracts. J. Agric. Food Chem. 2005, 53, 6896–6902. [Google Scholar] [CrossRef]
- Sanchez-Moreno, C.; Cao, G.; Ou, B.; Prior, R. Anthocyanin and proanthocyanidin content in selected white and red wines. Oxygen radical absorbance capacity comparison with nontraditional wines obtained from highbush blueberry. J. Agric. Food Chem. 2003, 51, 4889–4896. [Google Scholar] [CrossRef]
- Santos-Buelga, C.; Scalbert, A. Proanthocyanidins and tannin-like compounds snature, occurrence, dietary intake, and effects on nutrition and health. J. Sci. Food Agric. 2000, 80, 1094–1117. [Google Scholar] [CrossRef]
- Salta, J.; Martins, A.; Santos, R.S.; Neng, N.R.; Nogueira, J.M.F.; Justino, J.; Rauter, A.P. Phenolic composition and antioxidant activity of Rocha pear and other pear cultivars—A comparative study. J. Funct. Foods 2010, 2, 153–157. [Google Scholar] [CrossRef]
- Huang, S.C.; Lee, F.T.; Kuo, T.Y.; Yang, J.H.; Chien, C.T. Attenuation of long-term Rhodiola rosea supplementation on exhaustive swimming-evoked oxidative stress in the rat tissues. Chin. J. Physiol. 2009, 52, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Abbey, E.L.; Rankin, J.W. Effect of quercetin supplementation on repeated-sprint performance, xanthine oxidase activity, and inflammation. Int. J. Sport Nutr. Exerc. Metab. 2011, 21, 91–96. [Google Scholar] [CrossRef] [PubMed]
- König, D.; Wagner, K.H.; Elmadfa, I.; Berg, A. Exercise and oxidative stress: Significance of antioxidants with reference to inflammatory, muscular, and systemic stress. Exerc. Immunol. Rev. 2001, 7, 108–133. [Google Scholar] [PubMed]
- Malaguti, M.; Angeloni, C.; Hrelia, S. Polyphenols in exercise performance and prevention of exercise-induced muscle damage. Oxid. Med. Cell Longev. 2013, 2013, 825928. [Google Scholar] [CrossRef] [PubMed]
- Tung, Y.T.; Lin, L.C.; Liu, Y.L.; Ho, S.T.; Lin, C.Y.; Chuang, H.L.; Chiu, C.C.; Huang, C.C.; Wu, J.H. Antioxidative phytochemicals from Rhododendron oldhamii Maxim. leaf extracts reduce serum uric acid levels in potassium oxonate-induced hyperuricemic mice. BMC Complement. Altern. Med. 2015, 15, 423. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Hsu, Y.J.; Wei, L.; Chen, Y.J.; Huang, C.C. Association of physical performance and biochemical profile of mice with intrinsic endurance swimming. Int. J. Med. Sci. 2016, 13, 892–901. [Google Scholar] [CrossRef] [Green Version]
- Ho, C.S.; Tung, Y.T.; Kung, W.M.; Huang, W.C.; Leung, W.K.; Huang, C.C.; Wu, J.H. Effect of Coriolus versicolor mycelia extract on exercise performance and physical fatigue in mice. Int. J. Med. Sci. 2017, 14, 1110–1117. [Google Scholar] [CrossRef]
- Hsiao, C.Y.; Hsu, Y.J.; Tung, Y.T.; Lee, M.C.; Huang, C.C.; Hsieh, C.C. Effects of Antrodia camphorata and Panax ginseng supplementation on anti-fatigue properties in mice. J. Vet. Med. Sci. 2018, 80, 284–291. [Google Scholar] [CrossRef]
- Hsu, Y.J.; Huang, W.C.; Chiu, C.C.; Liu, Y.L.; Chiu, W.C.; Chiu, C.H.; Chiu, Y.S.; Huang, C.C. Capsaicin supplementation reduces physical fatigue and improves exercise performance in mice. Nutrients 2016, 8, 648. [Google Scholar] [CrossRef]
- Daviglus, M.L.; Lloyd-Jones, D.M.; Pirzada, A. Preventing cardiovascular disease in the 21st century: Therapeutic and preventive implications of current evidence. Am. J. Cardiovasc. Drugs 2006, 6, 87–101. [Google Scholar] [CrossRef]
- Bertram, M.Y.; Lim, S.S.; Barendregt, J.J.; Vos, T. Assessing the cost-effectiveness of drug and lifestyle intervention following opportunistic screening for pre-diabetes in primary care. Diabetologia 2010, 53, 875–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Church, T. Exercise in obesity, metabolic syndrome, and diabetes. Prog. Cardiovasc. Dis. 2011, 53, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Deweerdt, S. Prevention: Activity is the best medicine. Nature 2011, 475, S16–S17. [Google Scholar] [CrossRef] [PubMed]
- Hurley, B.F.; Hanson, E.D.; Sheaff, A.K. Strength training as a countermeasure to aging muscle and chronic disease. Sports Med. 2011, 41, 289–306. [Google Scholar] [CrossRef] [PubMed]
- Mastaloudis, A.; Leonard, S.W.; Traber, M.G. Oxidative stress in athletes during extreme endurance exercise. Free Radic Biol. Med. 2001, 31, 911–922. [Google Scholar] [CrossRef]
- Steensberg, A.; Morrow, J.; Toft, A.D.; Bruunsgaard, H.; Pedersen, B.K. Prolonged exercise, lymphocyte apoptosis and F2-isoprostanes. Eur. J. Appl. Physiol. 2002, 87, 38–42. [Google Scholar] [CrossRef]
- Malaguti, M.; Angeloni, C.; Garatachea, N.; Baldini, M.; Leoncini, E.; Collado, P.S.; Teti, G.; Falconi, M.; Gonzalez-Gallego, J.; Hrelia, S. Sulforaphane treatment protects skeletal muscle against damage induced by exhaustive exercise in rats. J. Appl. Physiol. 2009, 107, 1028–1036. [Google Scholar] [CrossRef] [Green Version]
- Navalta, J.W.; McFarlin, B.K.; Lyons, T.S. Does exercise really induce lymphocyte apoptosis? Front. Biosci. 2010, 2, 478–488. [Google Scholar] [CrossRef]
- Sastre, J.; Asensi, M.; Gascó, E.; Pallardó, F.V.; Ferrero, J.A.; Furukawa, T.; Viña, J. Exhaustive physical exercise causes oxidation of glutathione status in blood: Prevention by antioxidant administration. Am. J. Physiol. 1992, 263, 992–995. [Google Scholar] [CrossRef]
- Viña, J.; Gimeno, A.; Sastre, J.; Desco, C.; Asensi, M.; Pallardó, F.V.; Cuesta, A.; Ferrero, J.A.; Terada, L.S.; Repine, J.E. Mechanism of free radical production in exhaustive exercise in humans and rats; role of xanthine oxidase and protection by allopurinol. Iubmb Life 2000, 49, 539–544. [Google Scholar] [PubMed]
- Armstrong, R.B.; Ogilvie, R.W.; Schwane, J.A. Eccentric exercise-induced injury to rat skeletal muscle. J. Appl. Physiol. Respir. Env. Exerc. Physiol. 1983, 54, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Popovic, L.M.; Mitic, N.R.; Radic, I.; Miric, D.; Kisic, B.; Krdzic, B.; Djokic, T. The effect of exhaustive exercise on oxidative stress generation and antioxidant defense in guinea pigs. Adv. Clin. Exp. Med. 2012, 21, 313–320. [Google Scholar] [PubMed]
- McGinley, C.; Shafat, A.; Donnelly, A.E. Does antioxidant vitamin supplementation protect against muscle damage? Sports Med. 2009, 39, 1011–1032. [Google Scholar] [CrossRef] [PubMed]
- Davison, G.; Gleeson, M. The effect of 2 weeks vitamin C supplementation on immunoendocrine responses to 2.5 h cycling exercise in man. Eur. J. Appl. Physiol. 2006, 97, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.; Williams, C.; McGregor, S.J.; Nicholas, C.W.; McArdle, F.; Jackson, M.J.; Powell, J.R. Prolonged vitamin C supplementation and recovery from demanding exercise. Int. J. Sport Nutr. Exerc. Metab. 2001, 11, 466–481. [Google Scholar] [CrossRef]
- Beaton, L.J.; Allan, D.A.; Tarnopolsky, M.A.; Tiidus, P.M.; Phillips, S.M. Contraction-induced muscle damage is unaffected by vitamin E supplementation. Med. Sci. Sports Exerc. 2002, 34, 798–805. [Google Scholar] [CrossRef] [PubMed]
- Filaire, E.; Massart, A.; Rouveix, M.; Portier, H.; Rosado, F.; Durand, D. Effects of 6 weeks of n-3 fatty acids and antioxidant mixture on lipid peroxidation at rest and postexercise. Eur. J. Appl. Physiol. 2011, 111, 1829–1839. [Google Scholar] [CrossRef]
- Malaguti, M.; Baldini, M.; Angeloni, C.; Biagi, P.; Hrelia, S. High-protein-PUFA supplementation, red blood cell membranes, and plasma antioxidant activity in volleyball athletes. Int. J. Sport Nutr. Exerc. Metab. 2008, 18, 301–312. [Google Scholar] [CrossRef]
- Phillips, T.; Childs, A.C.; Dreon, D.M.; Phinney, S.; Leeuwenburgh, C. A dietary supplement attenuates IL-6 and CRP after eccentric exercise in untrained males. Med. Sci. Sports Exerc. 2003, 35, 2032–2037. [Google Scholar] [CrossRef]
- Zitterl-Eglseer, K.; Sosa, S.; Jurenitsch, J.; Schubert-Zsilavecz, M.; Della Loggia, R.; Tubaro, A.; Bertoldi, M.; Franz, C. Anti-oedematous activities of the main triterpendiol esters of marigold (Calendula officinalis L.). J. Ethnopharmacol. 1997, 57, 139–144. [Google Scholar] [CrossRef]
- Cordova, C.A.; Siqueira, I.R.; Netto, C.A.; Yunes, R.A.; Volpato, A.M.; Cechinel Filho, V.; Curi-Pedrosa, R.; Creczynski-Pasa, T.B. Protective properties of butanolic extract of the Calendula officinalis L. (marigold) against lipid peroxidation of rat liver microsomes and action as free radical scavenger. Redox Rep. 2002, 7, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Bruneton, J. In Pharmacognosy, Phytochemistry, Medical Plants; Intercept Limited: Andover, MA, USA, 1995; pp. 309–310. [Google Scholar]
- Kalt, W.; McDonald, J.E.; Ricker, R.D.; Lu, X. Anthocyanin content and profile within and among blueberry species. Can. J. Plant Sci. 1999, 79, 617–623. [Google Scholar] [CrossRef]
- Bettini, V.; Aragno, R.; Bettini, M.; Braggion, G.; Calore, L.; Concolato, M. Vasodilator and inhibitory effects of Vaccinium myrtillus anthocyanosides on the contractile responses of coronary artery segments to acetylcholine: Role of the prostacyclins and of the endothelium-derived relaxing factor. Fitoterapia 1991, 62, 15–28. [Google Scholar]
- Laplaud, P.M.; Lelubre, A.; Chapman, M.J. Antioxidant action of Vaccinium myrtillus extract on human low density lipoproteins in vitro: Initial observations. Fundam. Clin. Pharm. 1997, 11, 35–40. [Google Scholar] [CrossRef]
- Morazzoni, P.; Livio, S.; Scilingo, A.; Malandrino, S. Vaccinium myrtillus anthocyanosides pharmacokinetics in rats. Arzneimittelforschung 1991, 41, 128–131. [Google Scholar] [PubMed]
- Morazzoni, P.; Bombardelli, E. Vaccinium myrtillus. Fitoterapia 1996, 67, 3–30. [Google Scholar]
- Morazzoni, P.; Magistretti, M.J. Activity of myrtocyan, anthocyanoside complex from Vaccinium myrtillus (VMA), on platelet aggregation and adhesiveness. Fitoterapia 1990, 61, 13–21. [Google Scholar]
- Preethi, K.C.; Kuttan, G.; Kuttan, R. Antioxidant potential of an extract of Calendula officinalis flowers in vitro. and in vivo. Pharm. Biol. 2006, 44, 691–697. [Google Scholar] [CrossRef]
- Olennikov, D.N.; Kashchenko, N.I.; Chirikova, N.K.; Akobirshoeva, A.; Zilfikarov, I.N.; Vennos, C. Isorhamnetin and quercetin derivatives as anti-acetylcholinesterase principles of marigold (Calendula officinalis) flowers and preparations. Int. J. Mol. Sci. 2017, 18, 1685. [Google Scholar] [CrossRef]
- Bryan-Thomas, J. A comparative study of the antioxidant activity (DPPH), total flavonoid, total tannin, total polyphenol levels in plant extracts of the Annona muricata, Ribes nigrum and Manilkara zapota. Int. J. Sci. Res. Public. 2016, 6, 490–494. [Google Scholar]
- Saral, Ö.; Ölmez, Z.; Şahin, H.; Turkish, J. Comparison of antioxidant properties of wild blueberries (Vaccinium arctostaphylos L. and Vaccinium myrtillus L.) with cultivated blueberry varieties (Vaccinium corymbosum L.) in Artvin region of Turkey. Turk. J. Agric. Food Sci. Technol. 2014, 3, 40–44. [Google Scholar] [CrossRef]
- Matsunaga, N.; Chikaraishi, Y.; Shimazawa, M.; Yokota, S.; Hara, H. Vaccinium myrtillus (bilberry) extracts reduce angiogenesis in vitro and in vivo. Evid. Based Complement. Altern. Med. 2010, 7, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Rigane, G.; Ben Younes, S.; Ghazghazi, H.; Ben Salem, R. Investigation into the biological activities and chemical composition of Calendula officinalis L. growing in Tunisia. Int. Food Res. J. 2013, 20, 3001–3007. [Google Scholar]
- Bonarska-Kujawa, D.; Cyboran, S.; Żyłka, R.; Oszmiański, J.; Kleszczyńska, H. Biological activity of blackcurrant extracts (Ribes nigrum L.) in relation to erythrocyte membranes. Biomed. Res. Int. 2014, 2014, 783059. [Google Scholar] [CrossRef]
- Colak, N.; Primetta, A.K.; Riihinen, K.R.; Jaakola, L.; Grúz, J.; Strnad, M.; Torun, H.; Ayaza, F.A. Phenolic compounds and antioxidant capacity in different-colored and non-pigmented berries of bilberry (Vaccinium myrtillus L.). Food Biosci. 2017, 20, 67–78. [Google Scholar] [CrossRef]
- Song, E.K.; Kim, J.H.; Kim, J.S.; Cho, H.; Nan, J.X.; Sohn, D.H.; Ko, G.I.; Oh, H.; Kim, Y.C. Hepatoprotective phenolic constituents of Rhodiola sachalinensis on tacrine-induced cytotoxicity in Hep G2 cells. Phytother. Res. 2003, 17, 563–565. [Google Scholar] [CrossRef]
- Zhao, H.L.; Lin, S.X.; Jia, B.; Li, J.; Zhang, L.; Zhang, S. Inhibitory effects of salidroside on hypoxia-induced proliferation of rabbit pulmonary arterial smooth muscle cells. J. Fourth Mil. Med Univ. 2000, 21, 186–189. [Google Scholar]
- Zhang, W.S.; Zhu, L.Q.; Niu, F.L.; Deng, R.C.; Ma, C.X. Protective effects salidroside on injury induced by hypoxia/hypoglycemia in cultured neurons. Zhongguo Zhong Yao Za Zhi 2004, 29, 462–465. [Google Scholar]
- Kucinskaite, A.; Briedis, V.; Savickkas, A. Experimental analysis of therapeutic properties of Rhodiola rosea L. and its possible application in medicine. Medicina 2004, 40, 614–619. [Google Scholar]
- Wang, S.H.; Wang, W.J.; Wang, X.F.; Chen, W.H. Effects of salidroside on carbohydrate metabolism and differentiation 3T3-L1 adipocytes. Zhong Xi Yi Jie He Za Zhi 2004, 2, 193–195. [Google Scholar] [CrossRef]
- Li, H.B.; Ge, Y.K.; Zheng, X.X.; Zhang, L. Salidroside stimulated glucose uptake in skeletal muscle cells by activating AMP-activated protein kinase. Eur. J. Pharm. 2008, 588, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Vater, C.; Jacobi, A.; Liebers, C.; Zou, X.; Stiehler, M. Salidroside exerts angiogenic and cytoprotective effects on human bone marrow-derived endothelial progenitor cells via Akt/mTOR/p70S6K and MAPK signalling pathways. Br. J. Pharm. 2014, 171, 2440–2456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Donglian, C.; Huaixing, L.; Bende, T.; Lihua, S.; Ying, W. Anti-fatigue effects of salidroside in mice. J. Med Coll. PLA 2008, 23, 88–93. [Google Scholar] [CrossRef]
- Ma, C.; Hu, L.; Tao, G.; Lv, W.; Wang, H. An UPLC-MS-based metabolomics investigation on the anti-fatigue effect of salidroside in mice. J. Pharm. Biomed. Anal. 2015, 105, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Cai, D.L.; Li, H.X.; Tong, B.D.; Wang, Y.; Pei, S.P. Protective effects of salidroside on oxidative damage in fatigue mice. Zhong Xi Yi Jie He Xue Bao 2009, 7, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Deyama, T.; Nishibe, S.; Nakazawa, Y. Constituents and pharmacological effects of Eucommia and Siberian ginseng. Acta Pharm. Sin. 2001, 12, 1057–1070. [Google Scholar]
- Askari, G.; Ghiasvand, R.; Paknahad, Z.; Karimian, J.; Rabiee, K.; Sharifirad, G.; Feizi, A. The effects of quercetin supplementation on body composition, exercise performance and muscle damage indices in athletes. Int. J. Prev. Med. 2013, 4, 21–26. [Google Scholar]
- Askari, G.; Ghiasvand, R.; Karimian, J.; Feizi, A.; Paknahad, Z.; Sharifirad, G.; Hajishafiei, M. Does quercetin and vitamin C improve exercise performance, muscle damage, and body composition in male athletes? J. Res. Med. Sci. 2012, 17, 328–331. [Google Scholar]
- Chen, X.; Wu, Y.; Yang, T.; Wei, M.; Wang, Y.; Deng, X.; Shen, C.; Li, W.; Zhang, H.; Xu, W.; et al. Salidroside alleviates cachexia symptoms in mouse models of cancer cachexia via activating mTOR signalling. J. Cachexia Sarcopenia Muscle 2016, 7, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.C.; Hsu, M.C.; Huang, W.C.; Yang, H.R.; Hou, C.C. Triterpenoid-rich extract from Antrodia camphorata improves physical fatigue and exercise performance in mice. Evid. Based Complement. Altern. Med. 2012, 2012, 364741. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.L.; Wu, Q.P.; Huang, J.M.; Chen, R.; Cai, M.; Tan, J.B. Effects of L-malate on physical stamina and activities of enzymes related to the malate-aspartate shuttle in liver of mice. Physiol. Res. 2007, 56, 213–220. [Google Scholar] [PubMed]
- Shang, H.P.; Cao, S.H.; Wang, J.H.; Zheng, H.; Putheti, R. Glabridin from Chinese herb licorice inhibits fatigue in mice. Afr. J. Trad. Complementary Altern. Med. 2010, 7, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.N.; Lee, J.K.; Hong, S.H.; Kim, H.M. Herbkines increases physical stamina in mice. Biol. Pharm. Bull. 2004, 27, 117–119. [Google Scholar] [CrossRef] [PubMed]
- Tsopanakis, A. Stress hormonal factors, fatigue, and antioxidant responses to prolonged speed driving. Pharm. Biochem. Behav. 1998, 60, 747–751. [Google Scholar] [CrossRef]
- Sahlin, K.; Tonkonogi, M.; Söderlund, K. Energy supply and muscle fatigue in humans. Acta Physiol Scand. 1998, 162, 261–266. [Google Scholar] [CrossRef]
- Qian, E.W.; Ge, D.T.; Kong, S.K. Salidroside protects human erythrocytes against hydrogen peroxide-induced apoptosis. J. Nat. Prod. 2012, 75, 531–537. [Google Scholar] [CrossRef]
- Suh, S.H.; Paik, I.Y.; Jacobs, K. Regulation of blood glucose homeostasis during prolonged exercise. Mol. Cells 2007, 23, 272–279. [Google Scholar]
- Gonzalez, J.T.; Fuchs, C.J.; Betts, J.A.; van Loon, L.J. Liver glycogen metabolism during and after prolonged endurance-type exercise. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E543–E553. [Google Scholar] [CrossRef]
- Kim, J.H.; Pan, J.H.; Lee, E.S.; Kim, Y.J. L-Carnitine enhances exercise endurance capacity by promoting muscle oxidative metabolism in mice. Biochem. Biophys. Res. Commun. 2015, 464, 568–573. [Google Scholar] [CrossRef]
- Huang, W.C.; Huang, H.Y.; Hsu, Y.J.; Su, W.H.; Shen, S.Y.; Lee, M.C.; Lin, C.L.; Huang, C.C. The effects of thiamine tetrahydrofurfuryl disulfide on physiological adaption and exercise performance improvement. Nutrients 2018, 10, 851. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chen, Y.J.; Hsu, Y.J.; Wu, M.F.; Chiu, C.C.; Tung, Y.T.; Tsai, W.J.; Huang, W.C.; Huang, C.C. Effects of Ganoderma lucidum and ‘essence of chicken’ on physical fatigue recovery and exercise performance improvement. Chin. J. Physiol. 2018, 61, 372–383. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Point | Vehicle | CRV-1X | CRV-2X | CRV-5X |
|---|---|---|---|---|
| Lactate (mmol/L) | ||||
| Before swimming (A) | 2.9 ± 0.2 a | 2.9 ± 0.2 a | 2.9 ± 0.2 a | 2.9 ± 0.1 a |
| After swimming (B) | 8.5 ± 0.3 b | 5.7 ± 0.2 a | 5.5 ± 0.2 a | 5.4 ± 0.2 a |
| After a 20-min rest (C) | 6.4 ± 0.2 b | 4.0 ± 0.2 a | 3.7 ± 0.2 a | 3.5 ± 0.2 a |
| Rate of lactate production and clearance | ||||
| Production rate = B/A | 2.96 ± 0.1 b | 2.03 ± 0.1 a | 1.92 ± 0.07 a | 1.85 ± 0.04 a |
| Clearance rate = (B − C)/B | 0.25 ± 0.01 b | 0.31 ± 0.02 a | 0.32 ± 0.01 a | 0.34 ± 0.02 a |
| Time Point | Vehicle | CRV-1X | CRV-2X | CRV-5X |
|---|---|---|---|---|
| Initial BW (g) | 38.2 ± 0.4 | 38.1 ± 0.8 | 28.1 ± 0.7 | 38.2 ± 0.5 |
| 1st wk BW (g) | 40.0 ± 0.7 | 40.0 ± 1.0 | 40.0 ± 0.8 | 40.0 ± 0.3 |
| 2nd wk BW (g) | 40.5 ± 0.6 | 40.8 ± 0.8 | 40.5 ± 0.8 | 40.8 ± 0.4 |
| 3rd wk BW (g) | 41.0 ± 0.7 | 41.0 ± 1.0 | 41.0 ± 0.8 | 41.0 ± 0.3 |
| 4th wk BW (g) | 41.7 ± 0.6 | 41.7 ± 1.1 | 41.7 ± 0.8 | 41.9 ± 0.2 |
| Final BW (g) | 42.7 ± 0.7 | 42.7 ± 1.1 | 42.7 ± 1.0 | 42.7 ± 0.3 |
| Water intake (mL/mouse/day) | 9.7 ± 0.2 | 9.6 ± 0.2 | 9.7 ± 0.2 | 9.6 ± 0.1 |
| Chow 5001 (g/mouse/day) | 7.1 ± 0.1 | 7.0 ± 0.1 | 7.0 ± 0.1 | 7.1 ± 0.1 |
| Energy intake (Kcal/mouse/day) | 23.8 ± 0.3 | 23.6 ± 0.3 | 23.5 ± 0.3 | 23.8 ± 0.5 |
| Characteristic | Vehicle | CRV-1X | CRV-2X | CRV-5X |
|---|---|---|---|---|
| Liver (g) | 2.27 ± 0.07 | 2.29 ± 0.10 | 2.26 ± 0.08 | 2.25 ± 0.05 |
| Muscle (g) | 0.41 ± 0.01 | 0.41 ± 0.01 | 0.41 ± 0.01 | 0.41 ± 0.01 |
| Kidney (g) | 0.62 ± 0.02 | 0.62 ± 0.02 | 0.62 ± 0.02 | 0.63 ± 0.02 |
| Heart (g) | 0.25 ± 0.01 | 0.24 ± 0.01 | 0.24 ± 0.01 | 0.23 ± 0.01 |
| Lung (g) | 0.23 ± 0.01 | 0.23 ± 0.01 | 0.23 ± 0.01 | 0.23 ± 0.01 |
| EFP (g) | 0.29 ± 0.04 | 0.29 ± 0.03 | 0.29 ± 0.04 | 0.29 ± 0.02 |
| BAT (g) | 0.12 ± 0.003 | 0.12 ± 0.002 | 0.12 ± 0.01 | 0.12 ± 0.01 |
| Relative liver weight (%) | 5.30 ± 0.08 | 5.34 ± 0.12 | 5.27 ± 0.09 | 5.28 ± 0.07 |
| Relative muscle weight (%) | 0.95 ± 0.01 | 0.97 ± 0.01 | 0.97 ± 0.01 | 0.95 ± 0.01 |
| Relative kidney weight (%) | 1.44 ± 0.02 | 1.45 ± 0.03 | 1.46 ± 0.02 | 1.47 ± 0.03 |
| Relative heart weight (%) | 0.58 ± 0.01 | 0.57 ± 0.01 | 0.56 ± 0.02 | 0.52 ± 0.01 |
| Relative lung weight (%) | 0.54 ± 0.003 | 0.54 ± 0.01 | 0.54 ± 0.003 | 0.53 ± 0.01 |
| Relative EFP weight (%) | 0.67 ± 0.08 | 0.67 ± 0.06 | 0.67 ± 0.08 | 0.67 ± 0.05 |
| Relative BAT weight (%) | 0.28 ± 0.003 | 0.29 ± 0.003 | 0.28 ± 0.01 | 0.28 ± 0.01 |
| Parameter | Vehicle | CRV-1X | CRV-2X | CRV-5X |
|---|---|---|---|---|
| AST (U/L) | 73 ± 4 | 68 ± 3 | 69 ± 5 | 69 ± 2 |
| ALT (U/L) | 49 ± 3 | 41 ± 2 | 41 ± 2 | 42 ± 3 |
| CK (U/L) | 199 ± 25 | 173 ± 23 | 176 ± 26 | 176 ± 26 |
| GLU (mg/dL) | 149 ± 4 | 144 ± 3 | 145 ± 3 | 144 ± 4 |
| CREA (mg/dL) | 0.25 ± 0.01 | 0.24 ± 0.01 | 0.23 ± 0.01 | 0.24 ± 0.01 |
| BUN (mg/dL) | 20.5 ± 0.6 | 20.0 ± 0.5 | 20.4 ± 0.7 | 20.1 ± 0.7 |
| UA (mg/dL) | 1.2 ± 0.1 b | 0.8 ± 0.04 a | 0.8 ± 0.1 a | 0.7 ± 0.03 a |
| TC (mg/dL) | 145 ± 4 | 141 ± 5 | 142 ± 6 | 135 ± 4 |
| TG (mg/dL) | 174 ± 6 | 157 ± 5 | 157 ± 8 | 158 ± 6 |
| ALB (g/dL) | 2.9 ± 0.03 | 2.9 ± 0.03 | 2.9 ± 0.04 | 3.0 ± 0.05 |
| TP (g/dL) | 4.9 ± 0.1 | 4.9 ± 0.05 | 4.9 ± 0.04 | 4.9 ± 0.04 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tung, Y.-T.; Wu, M.-F.; Lee, M.-C.; Wu, J.-H.; Huang, C.-C.; Huang, W.-C. Antifatigue Activity and Exercise Performance of Phenolic-Rich Extracts from Calendula officinalis, Ribes nigrum, and Vaccinium myrtillus. Nutrients 2019, 11, 1715. https://doi.org/10.3390/nu11081715
Tung Y-T, Wu M-F, Lee M-C, Wu J-H, Huang C-C, Huang W-C. Antifatigue Activity and Exercise Performance of Phenolic-Rich Extracts from Calendula officinalis, Ribes nigrum, and Vaccinium myrtillus. Nutrients. 2019; 11(8):1715. https://doi.org/10.3390/nu11081715
Chicago/Turabian StyleTung, Yu-Tang, Ming-Fang Wu, Mon-Chien Lee, Jyh-Horng Wu, Chi-Chang Huang, and Wen-Ching Huang. 2019. "Antifatigue Activity and Exercise Performance of Phenolic-Rich Extracts from Calendula officinalis, Ribes nigrum, and Vaccinium myrtillus" Nutrients 11, no. 8: 1715. https://doi.org/10.3390/nu11081715