The Effect of Inulin on Lifespan, Related Gene Expression and Gut Microbiota in InRp5545/TM3 Mutant Drosophila melanogaster: A Preliminary Study



Abstract
1. Introduction
2. Materials and Methods
2.1. Fly Husbandry
2.2. Survival Analysis
2.3. 16S Amplicon Analysis
2.4. Quantitative Reverse Transcription-PCR (qRT-PCR) to Assess Drosophila Gene Expression
2.5. Statistics
3. Results
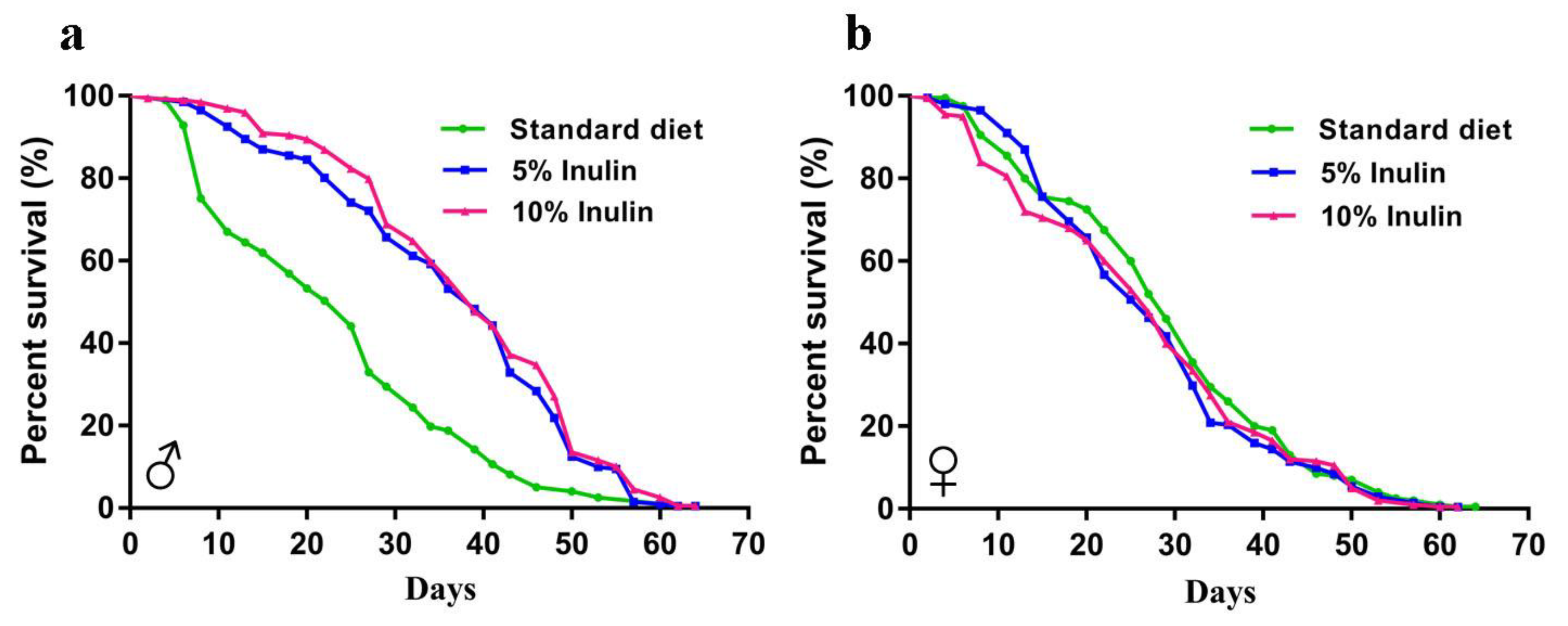
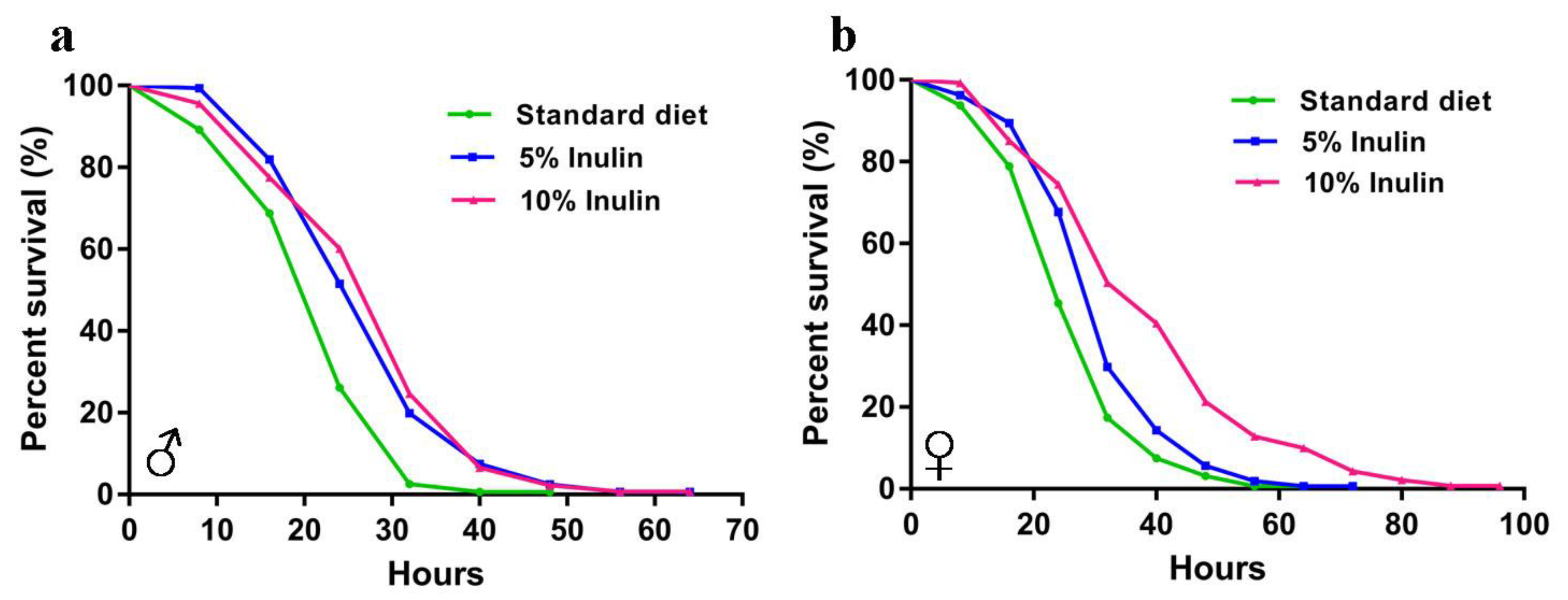
3.1. Inulin Extends the InRp5545/TM3 D. melanogaster Lifespan in Males but Not in Females
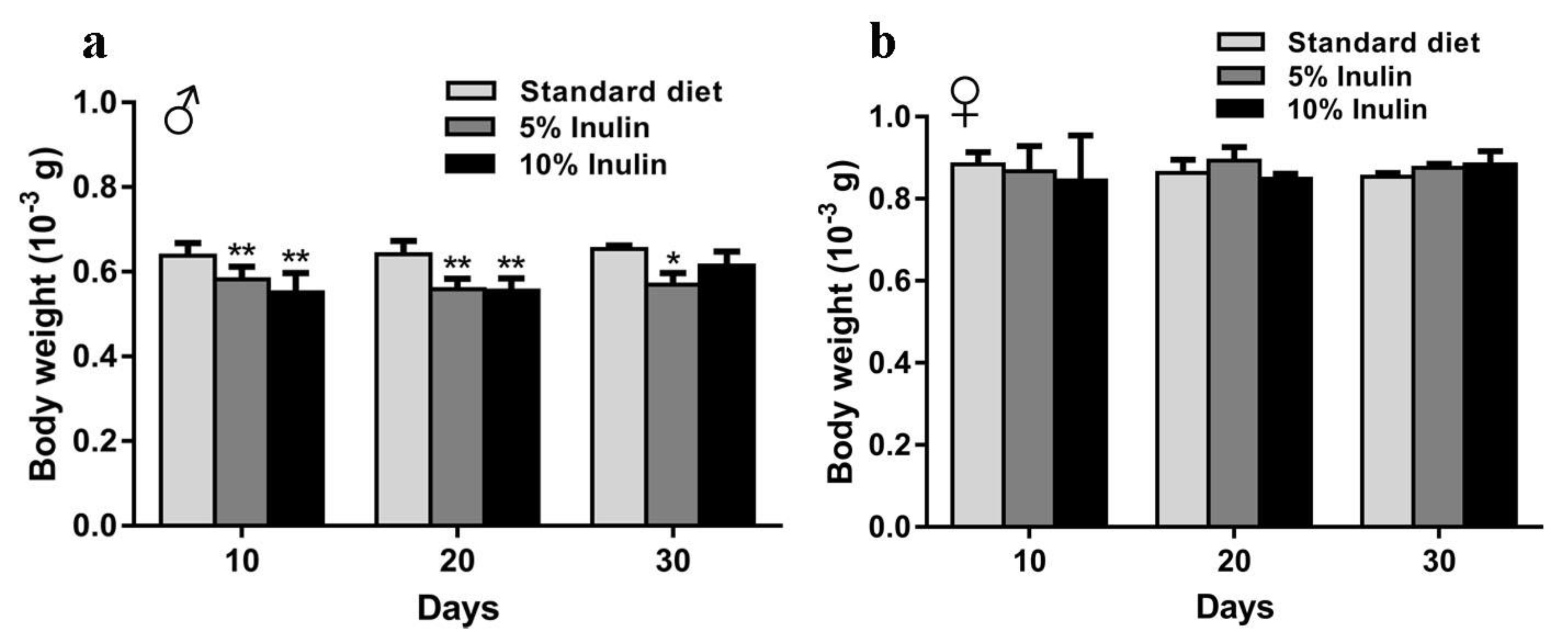
3.2. Inulin Modulates InRp5545/TM3 D. melanogaster Metabolism
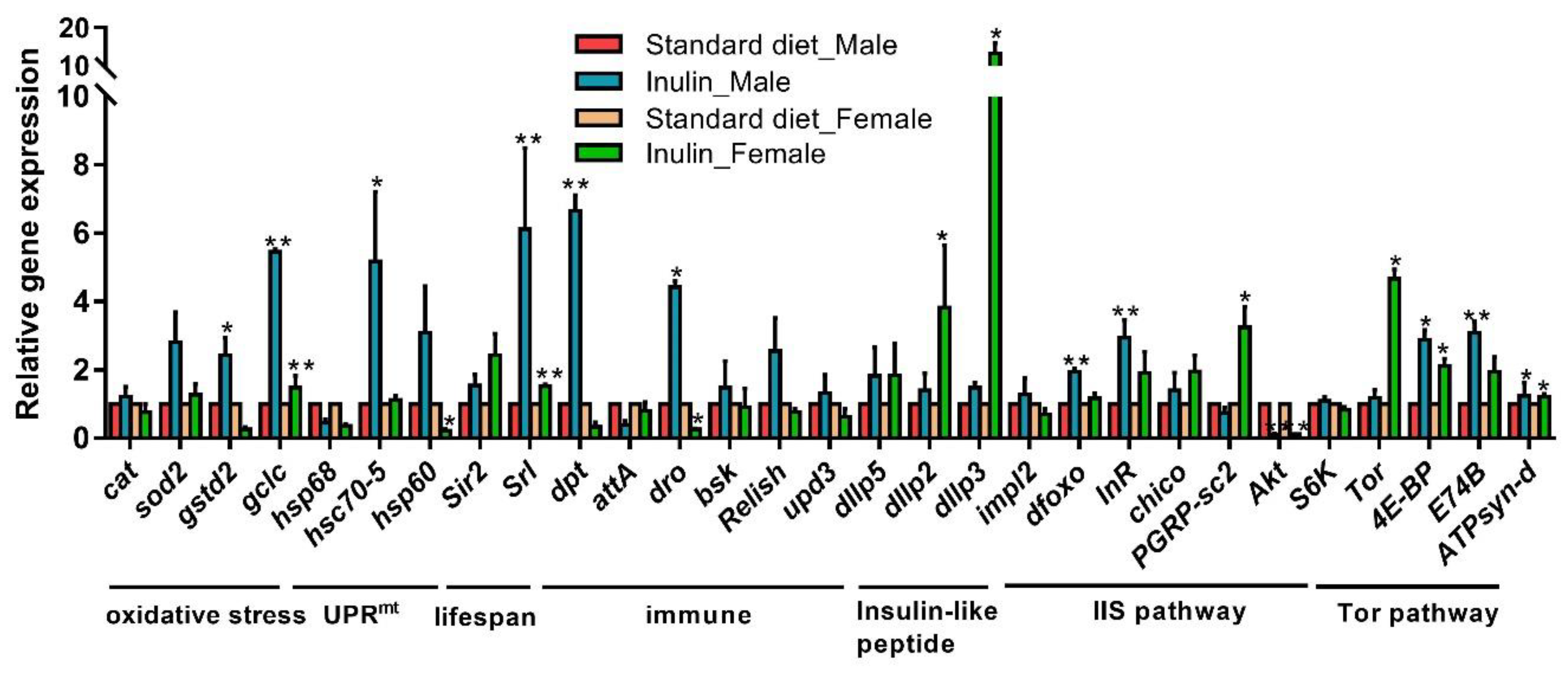
3.3. Different Gene Expression Patterns in InRp5545/TM3 Males and Females Fed with Inulin
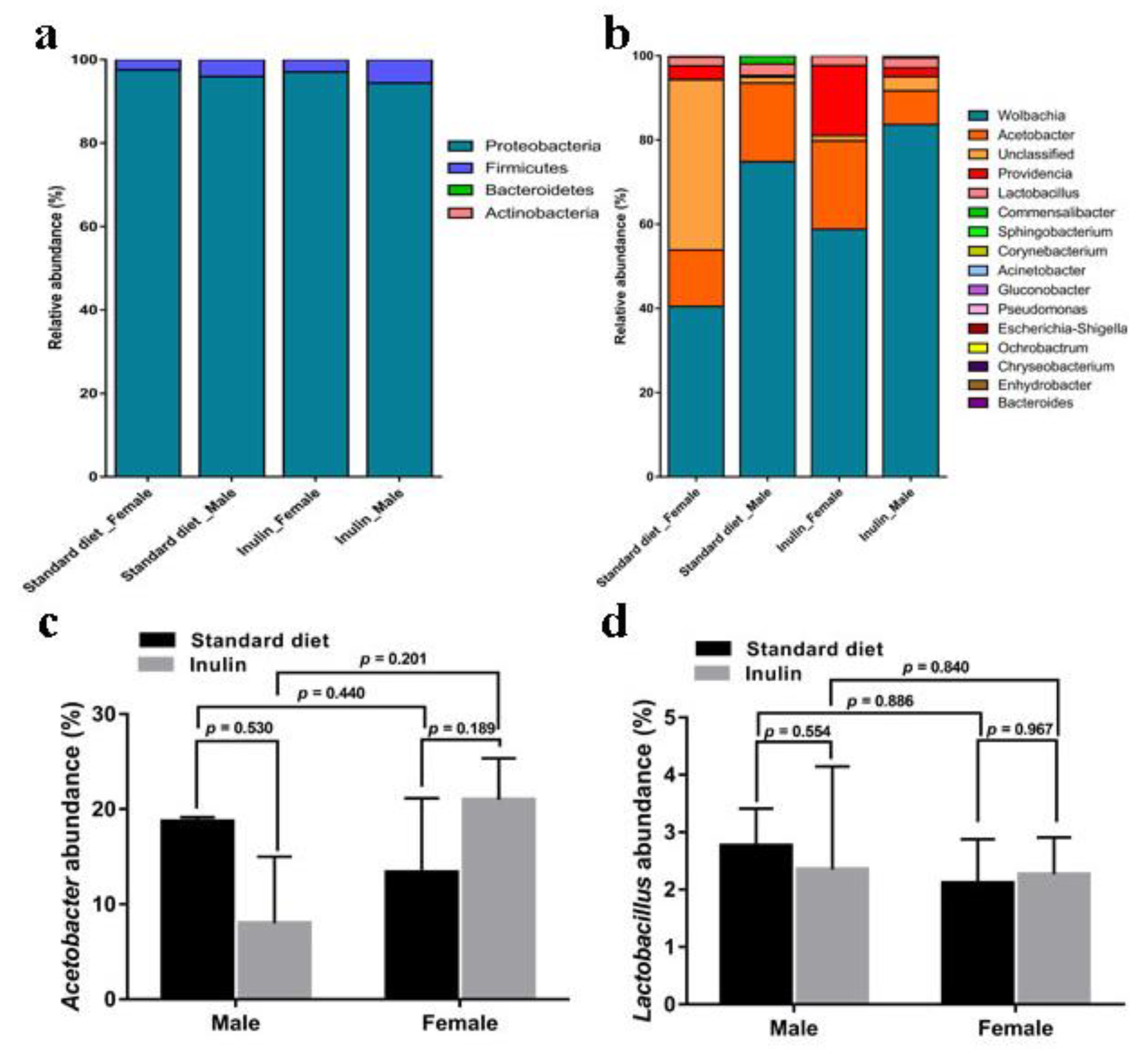
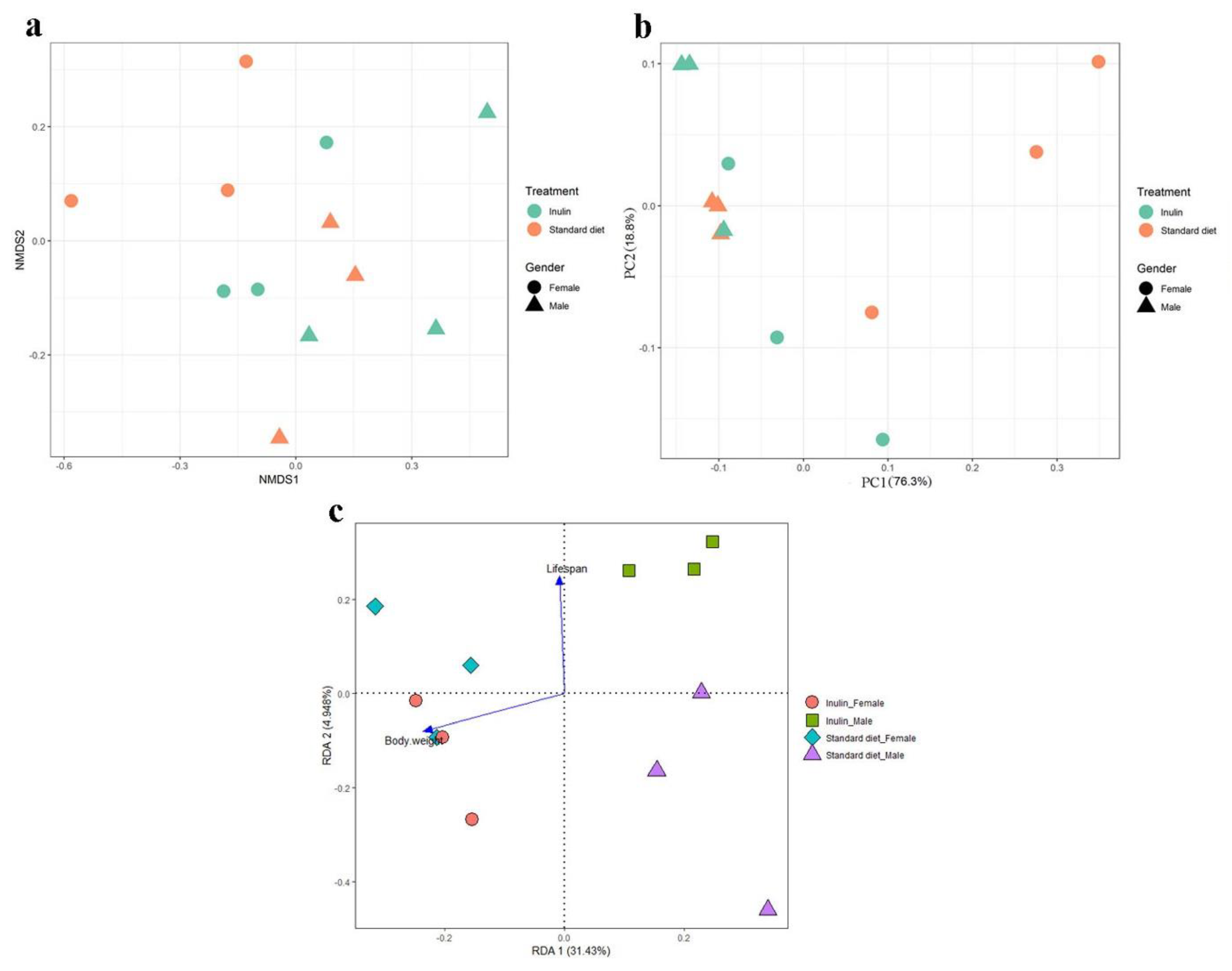
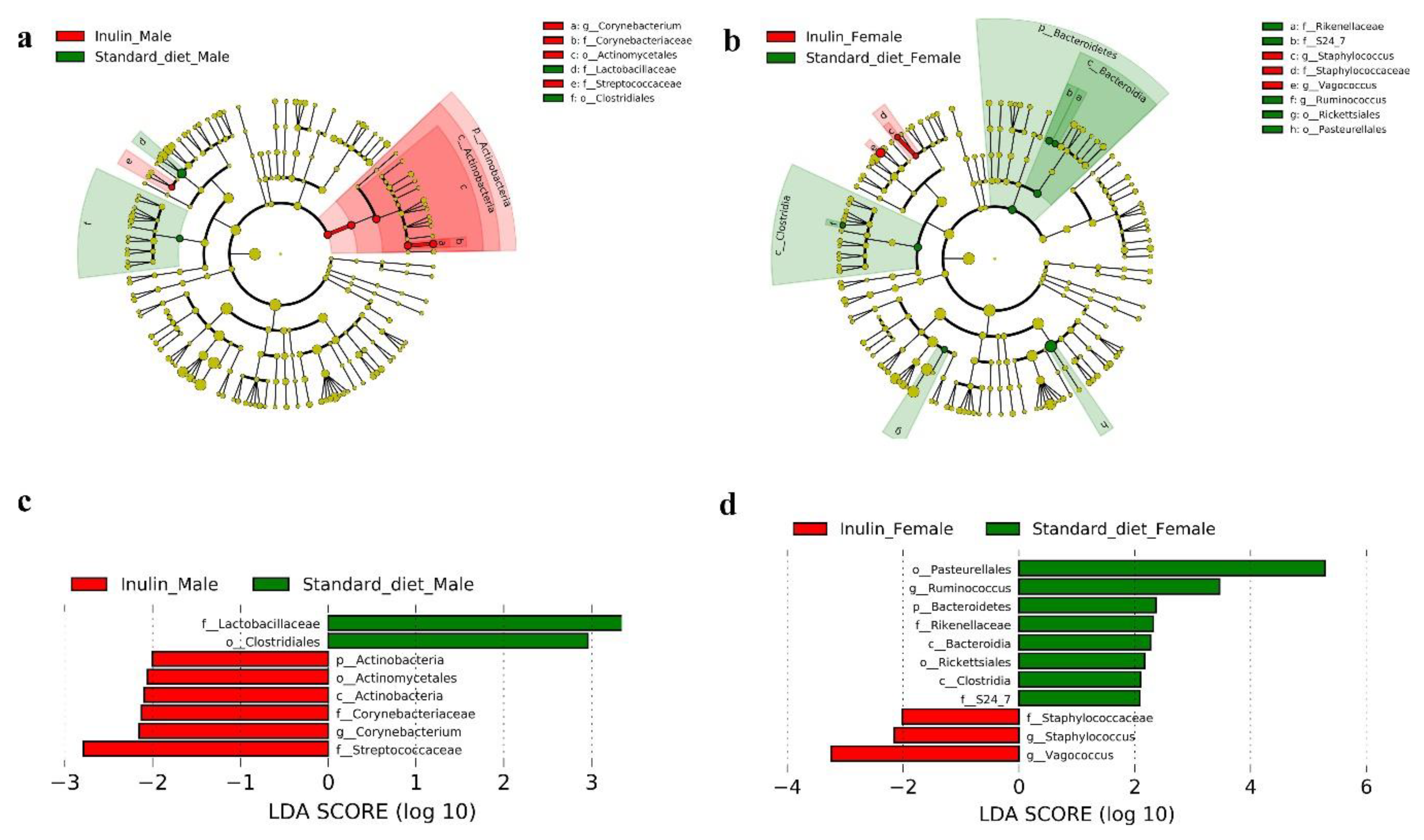
3.4. Gut Microbiota
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lillycrop, K.A. Effect of maternal diet on the epigenome: Implications for human metabolic disease. Proc. Nutr. Soc. 2011, 70, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S. Impact of maternal diet on the epigenome during in utero life and the developmental programming of diseases in childhood and adulthood. Nutrients 2015, 7, 9492–9507. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Zhang, C.; Yang, W.; Wang, Y.; Lin, Y.; Yang, P.; Yu, Q.; Fan, J.; Liu, E. Fat and cholesterol diet induced lipid metabolic disorders and insulin resistance in rabbit. Exp. Clin. Endocrinol. Diabetes 2009, 117, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Feige, J.N.; Lagouge, M.; Canto, C.; Strehle, A.; Houten, S.M.; Milne, J.C.; Lambert, P.D.; Mataki, C.; Elliott, P.J.; Auwerx, J. Specific SIRT1 activation mimics low energy levels and protects against diet-induced metabolic disorders by enhancing fat oxidation. Cell Metab. 2008, 8, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Zhao, S.; Li, D.; Chang, F.; Tian, X.; Huang, G.; Zhu, Z.; Liu, D.; Dou, X.; Li, S.; et al. Nutrient Intake Is associated with longevity characterization by metabolites and element profiles of healthy centenarians. Nutrients 2016, 8, 564. [Google Scholar] [CrossRef] [PubMed]
- Hur, K.Y.; Lee, M.-S. Gut microbiota and metabolic disorders. Diabetes Metab. J. 2015, 39, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Delzenne, N.M.; Neyrinck, A.M.; Bäckhed, F.; Cani, P.D. Targeting gut microbiota in obesity: Effects of prebiotics and probiotics. Nat. Rev. Endocrinol. 2011, 7, 639. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Beatty, E.R.; Wang, X.; Cummings, J.H. Selective stimulation of bifidobacteria in the human colon by oligofructose and inulin. Gastroenterology 1995, 108, 975–982. [Google Scholar] [CrossRef]
- Niness, K.R. Inulin and Oligofructose: What Are They? J. Nutr. 1999, 129, 1402S–1406S. [Google Scholar] [CrossRef] [PubMed]
- Tarini, J.; Wolever, T.M.S. The fermentable fibre inulin increases postprandial serum short-chain fatty acids and reduces free-fatty acids and ghrelin in healthy subjects. Appl. Physiol. Nutr. Metab. 2010, 35, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Kolida, S.; Tuohy, K.; Gibson, G.R. Prebiotic effects of inulin and oligofructose. Br. J. Nutr. 2007, 87, S193–S197. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed]
- Delzenne, N.M.; Cani, P.D.; Daubioul, C.; Neyrinck, A.M. Impact of inulin and oligofructose on gastrointestinal peptides. Br. J. Nutr. 2007, 93, S157–S161. [Google Scholar] [CrossRef]
- Baker, K.D.; Thummel, C.S. Diabetic larvae and obese flies—Emerging studies of metabolism in Drosophila. Cell Metab. 2007, 6, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Tatar, M.; Kopelman, A.; Epstein, D.; Tu, M.-P.; Yin, C.-M.; Garofalo, R.S. A mutant Drosophila insulin receptor homolog that extends life-span and impairs neuroendocrine function. Science 2001, 292, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Patterson, J.K.; Yasuda, K.; Welch, R.M.; Miller, D.D.; Lei, X.G. Supplemental dietary inulin of variable chain lengths alters intestinal bacterial populations in young pigs. J. Nutr. 2010, 140, 2158–2161. [Google Scholar] [CrossRef] [PubMed]
- Obata, F.; Fons, C.O.; Gould, A.P. Early-life exposure to low-dose oxidants can increase longevity via microbiome remodelling in Drosophila. Nat. Commun. 2018, 9, 975. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wheeler, C.T.; Alberico, T.; Sun, X.; Seeberger, J.; Laslo, M.; Spangler, E.; Kern, B.; de Cabo, R.; Zou, S. The effect of resveratrol on lifespan depends on both gender and dietary nutrient composition in Drosophila melanogaster. AGE 2013, 35, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.; Carey, J.R.; Liedo, P.; Ingram, D.K.; Müller, H.-G.; Wang, J.-L.; Yao, F.; Yu, B.; Zhou, A. The prolongevity effect of resveratrol depends on dietary composition and calorie intake in a tephritid fruit fly. Exp. Gerontol. 2009, 44, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Boyd, O.; Weng, P.; Sun, X.; Alberico, T.; Laslo, M.; Obenland, D.M.; Kern, B.; Zou, S. Nectarine promotes longevity in Drosophila melanogaster. Free Radic. Biol. Med. 2011, 50, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Schriner, S.E.; Kuramada, S.; Lopez, T.E.; Truong, S.; Pham, A.; Jafari, M. Extension of Drosophila lifespan by cinnamon through a sex-specific dependence on the insulin receptor substrate chico. Exp. Gerontol. 2014, 60, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, C.J. The genetics of ageing. Nature 2010, 464, 504. [Google Scholar] [CrossRef] [PubMed]
- Barzilai, N.; Huffman, D.M.; Muzumdar, R.H.; Bartke, A. The critical role of metabolic pathways in aging. Diabetes 2012, 61, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- Broughton, S.; Partridge, L. Insulin/IGF-like signalling, the central nervous system and aging. Biochem. J. 2009, 418, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Clancy, D.J.; Gems, D.; Harshman, L.G.; Oldham, S.; Stocker, H.; Hafen, E.; Leevers, S.J.; Partridge, L. Extension of life-span by loss of CHICO, a Drosophila insulin receptor substrate protein. Science 2001, 292, 104–106. [Google Scholar] [CrossRef] [PubMed]
- Broughton, S.J.; Piper, M.D.W.; Ikeya, T.; Bass, T.M.; Jacobson, J.; Driege, Y.; Martinez, P.; Hafen, E.; Withers, D.J.; Leevers, S.J. Longer lifespan, altered metabolism, and stress resistance in Drosophila from ablation of cells making insulin-like ligands. Proc. Natl. Acad. Sci. USA 2005, 102, 3105–3110. [Google Scholar] [CrossRef] [PubMed]
- Giannakou, M.E.; Goss, M.; Junger, M.A.; Hafen, E.; Leevers, S.J.; Partridge, L. Long-lived Drosophila with overexpressed dFOXO in adult fat body. Sci. Aging Knowl. Environ. 2004, 305, 361. [Google Scholar] [CrossRef] [PubMed]
- Kapahi, P.; Zid, B.M.; Harper, T.; Koslover, D.; Sapin, V.; Benzer, S. Regulation of lifespan in Drosophila by modulation of genes in the TOR signaling pathway. Curr. Biol. 2004, 14, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.C.; Bohmann, D.; Jasper, H. JNK extends life span and limits growth by antagonizing cellular and organism-wide responses to insulin signaling. Cell 2005, 121, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Karpac, J.; Jasper, H. Promoting longevity by maintaining metabolic and proliferative homeostasis. J. Exp. Biol. 2014, 217, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Katewa, S.D.; Kapahi, P. Role of TOR signaling in aging and related biological processes in Drosophila melanogaster. Exp. Gerontol. 2011, 46, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Partridge, L.; Alic, N.; Bjedov, I.; Piper, M.D.W. Ageing in Drosophila: The role of the insulin/Igf and TOR signalling network. Exp. Gerontol. 2011, 46, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Kockel, L.; Kerr, K.S.; Melnick, M.; Brückner, K.; Hebrok, M.; Perrimon, N. Dynamic switch of negative feedback regulation in Drosophila Akt–Tor signaling. PLOS Genet. 2010, 6, e1000990. [Google Scholar] [CrossRef] [PubMed]
- Zid, B.M.; Rogers, A.N.; Katewa, S.D.; Vargas, M.A.; Kolipinski, M.C.; Lu, T.A.; Benzer, S.; Kapahi, P. 4E-BP extends lifespan upon dietary restriction by enhancing mitochondrial activity in Drosophila. Cell 2009, 139, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.; Gravel, S.-P.; Chénard, V.; Sikström, K.; Zheng, L.; Alain, T.; Gandin, V.; Avizonis, D.; Arguello, M.; Zakaria, C.; et al. mTORC1 controls mitochondrial activity and biogenesis through 4E-BP-dependent translational regulation. Cell Metab. 2013, 18, 698–711. [Google Scholar] [CrossRef] [PubMed]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239. [Google Scholar] [CrossRef] [PubMed]
- Le Bourg, É. Oxidative stress, aging and longevity in Drosophila melanogaster. FEBS Lett. 2001, 498, 183–186. [Google Scholar] [CrossRef]
- Seifert, S.; Watzl, B. Inulin and oligofructose: Review of experimental data on immune modulation. J. Nutr. 2007, 137, 2563S–2567S. [Google Scholar] [CrossRef] [PubMed]
- Roller, M.; Rechkemmer, G.; Watzl, B. Prebiotic inulin enriched with oligofructose in combination with the probiotics Lactobacillus rhamnosus and Bifidobacterium lactis modulates intestinal immune functions in rats. J. Nutr. 2004, 134, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Kelly-Quagliana, K.A.; Nelson, P.D.; Buddington, R.K. Dietary oligofructose and inulin modulate immune functions in mice. Nutr. Res. 2003, 23, 257–267. [Google Scholar] [CrossRef]
- Watzl, B.; Girrbach, S.; Roller, M. Inulin, oligofructose and immunomodulation. Br. J. Nutr. 2007, 93, S49–S55. [Google Scholar] [CrossRef]
- Gourbeyre, P.; Desbuards, N.; Grémy, G.; Le Gall, S.; Champ, M.; Denery-Papini, S.; Bodinier, M. Exposure to a galactooligosaccharides/inulin prebiotic mix at different developmental time points differentially modulates immune responses in mice. J. Agric. Food Chem. 2012, 60, 11942–11951. [Google Scholar] [CrossRef] [PubMed]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lopez-Lluch, G.; Lewis, K.; et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006, 444, 337. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.E.; Cook, K.R.; Arvanitakis, A.V.; Hawley, R.S. Third chromosome balancer inversions disrupt protein-coding genes and influence distal recombination events in Drosophila melanogaster. G3 2016, 6, 1959–1967. [Google Scholar] [CrossRef] [PubMed]
- Broderick, N.A.; Buchon, N.; Lemaitre, B. Microbiota-induced changes in Drosophila melanogaster host gene expression and gut morphology. mBio 2014, 5, e01117-14. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.C.-N.; Dobson, A.J.; Douglas, A.E. Gut microbiota dictates the metabolic response of Drosophila to diet. J. Exp. Biol. 2014, 217, 1894–1901. [Google Scholar] [CrossRef] [PubMed]
- Chaston, J.M.; Newell, P.D.; Douglas, A.E. Metagenome-wide association of microbial determinants of host phenotype in Drosophila melanogaster. mBio 2014, 5, e01631-14. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-H.; Douglas Angela, E. Consumption of dietary sugar by gut bacteria determines Drosophila lipid content. Biol. Lett. 2015, 11, 20150469. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.C.; Kim, S.-H.; You, H.; Kim, B.; Kim, A.C.; Lee, K.-A.; Yoon, J.-H.; Ryu, J.-H.; Lee, W.-J. Drosophila microbiome modulates host developmental and metabolic homeostasis via insulin signaling. Science 2011, 334, 670–674. [Google Scholar] [CrossRef] [PubMed]
- Storelli, G.; Defaye, A.; Erkosar, B.; Hols, P.; Royet, J.; Leulier, F. Lactobacillus plantarum promotes Drosophila systemic growth by modulating hormonal signals through tor-dependent nutrient sensing. Cell Metab. 2011, 14, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.-H.; Kim, S.-H.; Lee, H.-Y.; Bai, J.Y.; Nam, Y.-D.; Bae, J.-W.; Lee, D.G.; Shin, S.C.; Ha, E.-M.; Lee, W.-J. Innate immune homeostasis by the homeobox gene Caudal and commensal-gut mutualism in Drosophila. Science 2008, 319, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Erkosar, B.; Leulier, F. Transient adult microbiota, gut homeostasis and longevity: Novel insights from the Drosophila model. FEBS Lett. 2014, 588, 4250–4257. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total n. of Flies | Mean (% Change) | Median (% Change) | Log-Rank (vs. Standard Diet) | |
|---|---|---|---|---|
| Male | ||||
| Standard diet | 196 | 34.91 | 22.08 | - |
| 5% Inulin | 200 | 53.70 (+53.83%) | 60.20 (+172.63%) | p < 0.0001 |
| 10% Inulin | 198 | 56.42 (+61.62%) | 62.31 (+182.19%) | p < 0.0001 |
| Female | ||||
| Standard diet | 199 | 42.60 | 32.50 | - |
| 5% Inulin | 200 | 40.73 (−4.39%) | 29.85 (−8.15%) | p = 0.3001 |
| 10% Inulin | 199 | 39.75 (−6.69%) | 33.50 (+3.08%) | p = 0.2570 |
| Total n. of Flies | Mean (% Change) | Median (% Change) | Log-Rank (vs. Standard Diet) | |
|---|---|---|---|---|
| Male | ||||
| Standard diet | 156 | 28.79 | 14.33 | - |
| 5% Inulin | 160 | 36.40 (+26.43%) | 13.66 (−4.65%) | p < 0.0001 |
| 10% Inulin | 137 | 36.81 (+27.87%) | 15.58 (+8.71%) | p < 0.0001 |
| Female | ||||
| Standard diet | 160 | 24.80 | 12.42 | - |
| 5% Inulin | 160 | 29.02 (+16.99%) | 14.28 (+15.00%) | p = 0.0003 |
| 10% Inulin | 140 | 35.82 (+44.41%) | 17.02 (+37.02%) | p < 0.0001 |
| Standard Diet Male | Standard Diet Female | Inulin Male | Inulin Female | |
|---|---|---|---|---|
| ACE | 25.74 ± 0.13 | 25.59 ± 4.68 (p = 0.966) | 29.13 ± 2.74 (p = 0.155) | 31.43 ± 9.24 (p = 0.753, 0.470) |
| Chao1 | 24.36 ± 0.69 | 24.17 ± 3.97 (p = 0.949) | 33.67 ± 8.18 (p = 0.184) | 25.72 ± 2.44 (p = 0.258, 0.661) |
| Shannon | 1.40 ± 0.06 | 1.86 ± 0.29 (p = 0.094) | 0.92 ± 0.36 (p = 0.135) | 1.78 ± 0.40 (p = 0.087, 0.819) |
| Simpson | 0.43 ± 0.02 | 0.64 ± 0.07 (p = 0.013) | 0.28 ± 0.13 (p = 0.198) | 0.57 ± 0.15 (p = 0.112, 0.601) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Y.; Sun, H.; Yang, W.; Ma, S.; Du, B.; Xu, H. The Effect of Inulin on Lifespan, Related Gene Expression and Gut Microbiota in InRp5545/TM3 Mutant Drosophila melanogaster: A Preliminary Study. Nutrients 2019, 11, 636. https://doi.org/10.3390/nu11030636
Dong Y, Sun H, Yang W, Ma S, Du B, Xu H. The Effect of Inulin on Lifespan, Related Gene Expression and Gut Microbiota in InRp5545/TM3 Mutant Drosophila melanogaster: A Preliminary Study. Nutrients. 2019; 11(3):636. https://doi.org/10.3390/nu11030636
Chicago/Turabian StyleDong, Yuling, Hao Sun, Weichao Yang, Shuang Ma, Beibei Du, and Hui Xu. 2019. "The Effect of Inulin on Lifespan, Related Gene Expression and Gut Microbiota in InRp5545/TM3 Mutant Drosophila melanogaster: A Preliminary Study" Nutrients 11, no. 3: 636. https://doi.org/10.3390/nu11030636
APA StyleDong, Y., Sun, H., Yang, W., Ma, S., Du, B., & Xu, H. (2019). The Effect of Inulin on Lifespan, Related Gene Expression and Gut Microbiota in InRp5545/TM3 Mutant Drosophila melanogaster: A Preliminary Study. Nutrients, 11(3), 636. https://doi.org/10.3390/nu11030636