1. Introduction
Childhood undernutrition is endemic throughout the world. Nutritional deficiency can severely impact neurodevelopment, and several lines of evidence indicate the importance of dietary long-chain polyunsaturated fatty acids (LCPUFA) in proper brain development. Within the brain, docosahexaenoic acid (DHA, 22:6
n-3) and arachidonic acid (AA, 20:4
n-6) are the most abundant LCPUFA. In humans, brain accumulation of LCPUFA begins prenatally and continues until the 2nd postnatal year as reviewed in Janssen & Kiliaan 2014 [
1]. During this period of infant neurodevelopment, LCPUFAs incorporate into cellular membranes, regulate gene expression, and facilitate neurogenesis [
2,
3]. Postnatal availability of LCPUFA is crucial for myelination of nerve fibers and neurite outgrowth [
4,
5,
6,
7]. Childhood deficiencies in LCPUFA are associated with cognitive, behavioral, and visual impairments in children and adolescents [
1].
The rat cerebellum is primarily developed and remodeled postnatally; similarly, human cerebellar development carries into the first postnatal years [
8,
9,
10,
11]. In rat cerebellum, non-neuronal cells dynamically change their numbers by postnatal day 25 and approximately 150 million neurons are added within the first three postnatal months [
9]. The cerebellum has long been recognized for regulating motor coordination but is also involved with higher cognitive function [
12]. Early rat studies demonstrated that the developing cerebellum is particularly susceptible to undernutrition [
13,
14].
DHA is obtained directly from the diet or is produced indirectly through its precursor α-linolenic acid (ALA, 18:3
n-3). Similarly, if AA is not obtained through dietary sources, it may be synthesized from its parent fatty acid linoleic acid (LA, 18:2
n-6). Although less abundant than DHA or AA, brain docosapentaenoic acid (DPA
n-6, 22:5
n-6), resulting from the elongation and desaturation of AA, is linked to roles in brain health and function. Elevated content of DPA
n-6 is considered a biomarker of
n-3 PUFA deficiency in animal models [
15]. The exact physiological role of DPA
n-6 within the brain is not entirely clear, but increased levels of DPA
n-6 are associated with deficits in learning and behavior [
16].
Dietary LCPUFA content alters gene expression throughout the body in a tissue-specific manner [
17]. LCPUFA directly bind and activate transcription factors that modulate expression of target genes [
18]. For example, peroxisome proliferator-activated receptors (PPARs) are ligand-dependent transcription factors that bind specific
n-3 and
n-6 PUFAs and regulate gene expression [
19]. Maternal
n-3 PUFA deficient diets lead to elevated postnatal brain D2 (dopamine) receptors and D2 receptor mRNA levels in rats [
20,
21]. Genes important in the processes of synaptic plasticity and cognition are modified in the brains of rats fed PUFA-enriched diets with the expression of transthyretin (
Ttr) being reduced and α-synuclein (
Snca) and calmodulins (
Calm1-3) being increased [
18,
22].
Previous means of studying global LCPUFA-dependent differences in the transcriptome of the rodent brain have relied on RNA or cDNA microarray technology that analyzed a fraction of the genome [
22,
23,
24,
25]. Microarrays offer high-throughput expression analysis for thousands of genes between experimental conditions; although, the results are largely dependent on the selection/quality of probes used on the chip, along with other limitations [
26]. Next-generation sequencing (NGS) or RNA-sequencing (RNA-seq) is a powerful alternative to microarray technology and is becoming more cost effective. NGS offers a global approach to measure differentially expressed genes (DEGs) that is not dependent on probes, and also displays greater sensitivity and dynamic range of quantification than microarrays [
26,
27].
We have estimated that a three week old rat pup corresponds to an 11 month old child that has reached 70% maximum brain weight, and where myelination is still occurring within the corpus callosum and cerebellum by using an online model (
http://www.translatingtime.net/) to compare brain development across species [
28]. In this study, we sought to identify the degree to which dietary
n-3 PUFA deficiency modifies gene expression within the cerebellum of young rats given the neurodevelopmental importance of LCPUFA in postnatal brain health. We tested the hypothesis that a diet deficient in
n-3 PUFA would result in modifications to the juvenile cerebellar transcriptome by characterizing the fatty acid profile and transcript abundance in the cerebellum of rats with variable levels of
n-3 PUFA in their diets. A subset of transcripts that appeared to be differentially abundant between dietary classes from analysis of the RNAseq data, were validated with Quantitative PCR (qPCR) using RNA from two separate cohorts of animals. Cerebellar FA analysis demonstrated a 4-fold increase in DPA
n-6, and slight increases of AA and docosatetraenoic acid (DTA; 22:4
n-6) in the cerebellum of CO-fed animals.
Nr4a3 is an immediate early gene (IEG) that does not require de novo protein synthesis for its expression, therefore, can be activated and transcribed rapidly.
Nr4a3 encodes a transcriptional activator that plays roles in neuroprotection and learning and memory formation.
Nr4a3 was confirmed in both cohorts of animals to be elevated over 2-fold in the cerebellum from the CO diet.
4. Discussion
Undernutrition during early brain development can result in serious deficits in cognition, behavior, and motor skills that may have lasting consequences [
41]. LCPUFA deficiency during brain development displays similar impairments [
1]. Still, data is limited in describing what genes are altered within the transcriptome of the juvenile cerebellum in response to dietary
n-3 PUFA deprivation.
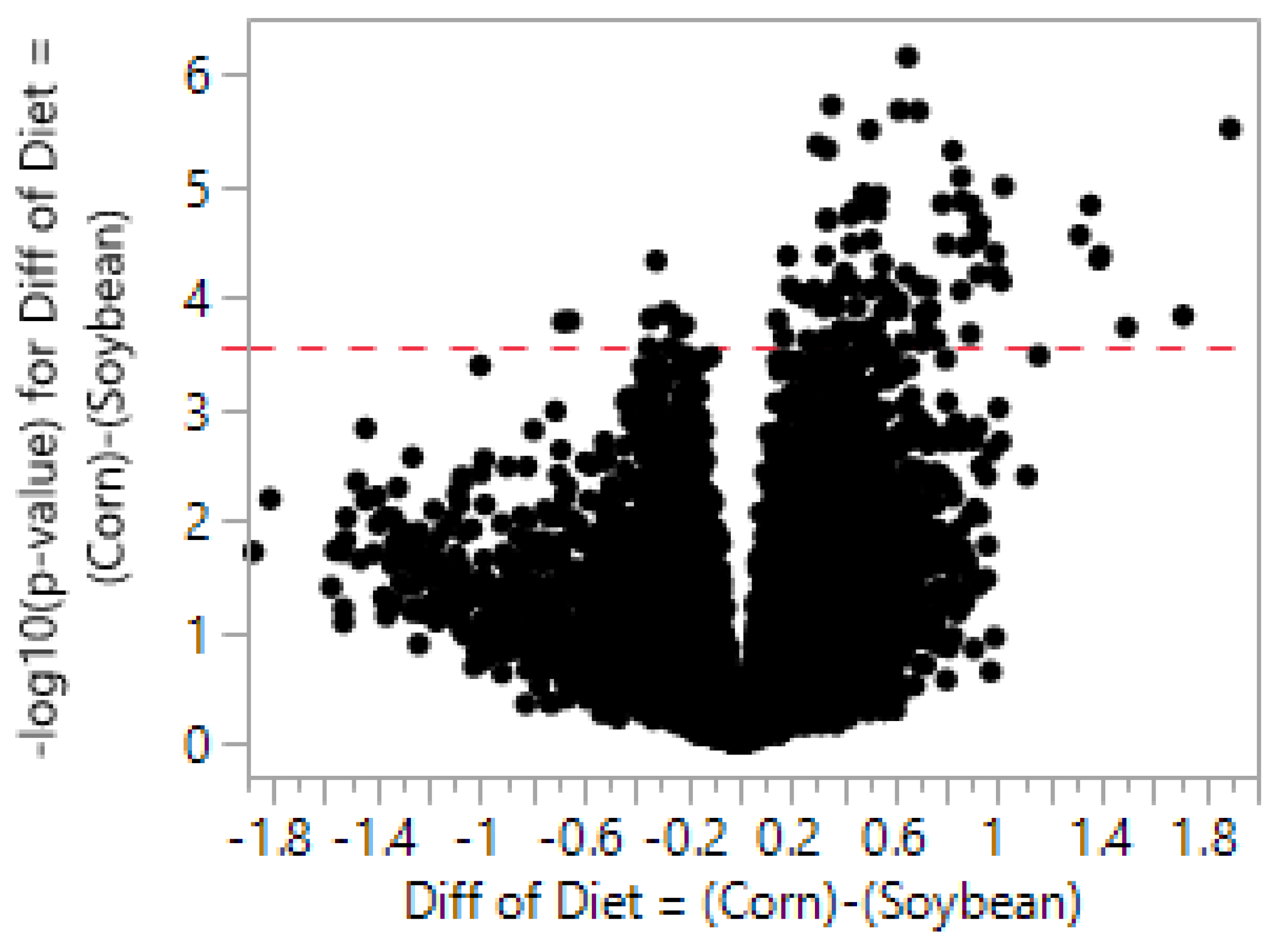
In this study we identified 137 potential dietary
n-3 PUFA-responsive genes within the cerebellar transcriptome of developing rats using NGS. In comparison, data obtained from the earlier cDNA microarray studies identified 102 DEGs [
22], 23 DEGs [
25], 24 DEGs [
24], and 6 DEGs [
23] in rat brains by modifying dietary
n-3 and
n-6 PUFA content. This correlates to roughly 0.50–0.70% (dependent on normalization) DEGs identified with NGS and the earlier studies ranged 0.20–3.40% (dependent on study). By using qPCR, we were only able to confirm that 6 of the 39 genes changed in the same direction (qualitatively consistent) with NGS data, correlating to roughly a 15% validation rate in the 1st cohort of animals. Of the 6 genes we were able to confirm in the 1st cohort, only 1 gene (
Nr4a3) could be confirmed in the repeated study (2nd cohort). To our knowledge, brain expression of
Nr4a3 has not previously been reported to be responsive to dietary
n-3 or
n-6 PUFA content and was not identified as a DEG in the microarray studies listed above. It is important to note, however, that the earlier studies used different rat strains, age of rats, duration on diets, oil composition and content of
n-3 and
n-6 PUFA within diets, whole brain or hippocampal tissue, and RNA samples were pooled 8–12 animals/group. So, a fair comparison between the earlier studies and the current study cannot be made.
The majority of the cerebellar DEG fold-changes in this study were <2-fold. This finding is consistent with a previous RNA microarray study examining DEGs within the cerebral cortex of young baboons that were fed diets that differed in amounts of DHA and AA for 12 weeks [
42]. Furthermore, a more recent microarray study that analyzed modifications in the mouse brain transcriptome by altering dietary
n-3 to
n-6 levels for 20 weeks, identified roughly 1000 genes of interest but less than 8% were >2-fold changes [
43]. These microarray studies identified more potential
n-3 and
n-6 PUFA responsive genes within the brain; however, the extent to which the DEGs were modified was low. While there are several important experimental design differences compared to our study, namely dietary
n-3 and
n-6 LCPUFA content and duration on diets, the LCPUFA-dependent modifications within the brain transcriptome was relatively small, which is consistent with our results. These results are quite different from a recent study analyzing effects of dietary
n-3 and
n-6 PUFA within the pig liver transcriptome that report over 3500 DEGs with average fold-changes of 9.4 [
44]. These previous data, in addition to our data, indicate that the brain transcriptome is tightly regulated with regard to dietary
n-3 PUFA.
The most prominent finding in this study was identifying and confirming the elevated expression of
Nr4a3 from CO in both cohorts of animals.
Nr4a3 (gene product—NOR1) is a transcriptional activator known for playing important roles in cell survival and apoptosis [
45,
46]. The
Nr4a gene family regulates the neuroendocrine system which is responsible for energy utilization and metabolism [
47]. In the brain,
Nr4a3 is capable of providing neuroprotection and promoting neuronal survival upon excitotoxic and oxidative stress generated from kainic acid, a glutamate analog, in the hippocampus of mice [
48,
49]. Moreover,
Nr4a3 and other related
Nr4a gene members are elevated after learning and memory formation in mice and their activation is associated with increased synaptic plasticity [
50,
51]. Previous studies using mouse skeletal muscle cells and tissue have shown that
Nr4a3 plays a role in regulating genes involved with fatty acid utilization and is inducible by the beta-adrenergic receptor agonist isoprenaline [
52,
53]. In beta cells of the pancreas,
Nr4a3 is inducible by FAs (palmitate) and can modulate secretion of insulin and expression of the
Ins1 and
Ins2 genes [
54]. From our results, it is not clear if the modified level of
Nr4a3 within the cerebellum is an adaptive response to the stress imposed by dietary
n-3 PUFA deprivation, or if dietary
n-3 PUFA deprivation is causing cognitive disturbances. More work is needed to answer these questions.
Reductions in DHA are observed in the rat brain from trans-generational dietary
n-3 PUFA deprivation [
55,
56]. Under natural conditions (non-trans-generationally deprived of dietary
n-3 PUFA), the postnatal rat brain is effective in accumulating DHA required for growth and is effective in limiting DHA loss during times of dietary
n-3 deprivation [
57]. The juvenile
n-3 PUFA deficient rat model used in this study is efficient in depleting
n-3 PUFA from livers and altering the brain FA content [
29,
30]. In prior studies, we have observed small (~10%) reductions in cerebellar levels of DHA and increases in DPA
n-6, AA, and DTA using the CO diet in postnatal rats [
29,
30]. The cerebellar FA content reported here is in good agreement with our earlier studies, with the exception of the DHA reduction in the brains of young rats fed CO (~0.8 mol% ALA) for four weeks. This finding is not unexpected, since the DHA reduction demonstrated previously was minor and ~15 weeks is needed to have more prominent reductions in brain DHA when rats are fed diets utilizing this percentage of ALA [
58].
A longer postnatal duration of dietary n-3 PUFA deprivation or initiating deprivation prenatally may have resulted in a more pronounced effect to the LCPUFA content within the cerebellum and reduced DHA more effectively. More prominent alterations to the cerebellar LCPUFA composition could have resulted in greater fold changes to the DEGs identified in this study or generated an entirely different list of DEGs. Additional studies are needed to assess the possible health consequences associated with the cerebellar LCPUFA-dependent modification of Nr4a3 expression during development and whether these effects are reversible.
This study has limitations. The cellular heterogeneity of the postnatal cerebellum coupled to its dynamic growth and remodeling during this period of time add complexity in measuring DEGs within this important developing tissue. A majority of the DEGS identified displayed only minor changes (<2-fold) and from a statistical standpoint, there is an abundance of NGS data (n = 10/group). This results in increased precision around the estimated means and potentially Type II statistical errors. However, we performed qPCR validations on a 2nd cohort of animals to provide biological replication and reduce the impacts of statistical anomalies.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





