Mechanism and Potential of Egg Consumption and Egg Bioactive Components on Type-2 Diabetes


1
Department of Agricultural, Food and Nutritional Science, University of Alberta, Edmonton, AB T6G 2R3, Canada
2
Cardiovascular Research Centre, University of Alberta, Edmonton, AB T6G 2R3, Canada
*
Author to whom correspondence should be addressed.
Nutrients 2019, 11(2), 357; https://doi.org/10.3390/nu11020357
Submission received: 26 December 2018
/
Revised: 1 February 2019
/
Accepted: 1 February 2019
/
Published: 8 February 2019
(This article belongs to the Special Issue Egg Intake and Human Health)
Abstract
:Type-2 diabetes (T2D) is one of the major global health challenges and a substantial economic burden. Egg and egg-derived components have been indicated to possess antioxidant, anti-inflammatory, anti-hypertensive, immunomodulatory, and anti-cancer activities. However, the scientific evidence about the benefits of egg on T2D is debatable. The relationship between egg consumption and the risk of T2D from observational epidemiological studies is not consistent. Interventional clinical studies, however, provide promising evidence that egg consumption ameliorates the risk of T2D. Current research progress also indicates that some egg components and egg-derived peptides might be beneficial in the context of T2D, in terms of insulin secretion and sensitivity, oxidative stress, and inflammation, suggesting possible application on T2D management. The current review summarizes recent clinical investigations related to the influence of egg consumption on T2D risk and in vivo and in vitro studies on the effect and mechanism of egg components and egg-derived peptides on T2D.
1. Introduction
Diabetes is a rapidly growing public health problem worldwide, which is characterized by prolonged hyperglycemia and impaired insulin secretion together with or without insulin resistance. Type-1 diabetes is caused by the cell-mediated autoimmune destruction of pancreatic β-cells and accounts for 5–10% of diabetic cases. The majority of diabetes victims (90–95%) are affected by type-2 diabetes (T2D), marked by insulin resistance and relative insulin deficiency [1]. T2D is closely associated with life style—especially diet and exercise—and is preventable.
According to the World Health Organization, between 1980 and 2014, the global prevalence of diabetes among adults has been increasing from 4.7% (108 million people) to 8.5% (422 million people) [2], in which the westernized lifestyle with dietary changes and lack of exercise is believed to play a role [3,4,5]. Thus, the identification of effective dietary components that can reduce the risk of T2D or slow down the progression of complications is important to improve the quality of life for diabetic patients and people at risk of T2D. Egg is one of the major protein sources in the diet. Also, egg is composed of a broad range of health beneficial components including amino acids, vitamins, minerals, and carotenoids [6]. To date, there are some evidence showing the beneficial property of some egg components and egg-derived peptides in the context of T2D, which are often associated with their anti-oxidative and anti-inflammatory properties [6,7,8]. In this review, we will provide an update on the mechanism and potential of egg, egg components, and egg-derived peptides on T2D management.
2. Egg Consumption and the Risk of T2D
The in vivo study addressing the effect of whole egg consumption on T2D is very limited. In Zucker diabetic fatty rats, eight-week feeding of dried whole egg-based diet was shown to reduce blood glucose and triglyceride concentrations, decrease the percentage of body fat, suppress weight gain, and increase circulating 25-hydroxycholecalciferol level, compared to diabetic rats fed with casein-based diet [9,10].
In terms of human study, contradicting observational evidence has been gained from prospective cohort [11,12,13,14,15,16,17,18,19,20,21,22,23], case-control [24], and systematic studies [25,26,27,28,29] in different populations on the relationship between the consumption of egg and T2D risk. A positive association between egg intake and the risk of T2D seemed to be only reported in the US population [12,17,23,29,30,31]. For example, Djousse et al. showed in a 20-year cohort study with 22,071 male physicians (≥40 years at entry) and 39,876 female healthcare workers (≥45 years at entry) in the US that the consumption of 7 eggs/week increased the risk of T2D in both men (Hazard Ratio, HR, 1.58, CI 1.25–2.01) and women (HR, 1.77, CI 1.28–2.43), compared to subjects who had egg consumption of <1/week [12]. In another large cohort study, a positive association between the frequency of egg consumption and the incidence of T2D was reported in 4568 African American subjects (average age: 55 ± 13 years, 64% female) [17]. In addition, higher egg intake was suggested to be correlated with higher blood glucose concentration and negative cardiovascular outcomes in T2D patients in a cohort study conducted in the United Kingdom [32]. However, it has been shown in a prospective cohort study performed in eastern Finland that there was a negative association between egg consumption and the risk of T2D in middle-aged and older men [18,33]. An inverse association was also reported between egg intake and fasting plasma glucose and serum C-reactive protein concentrations [18]. In a Korean study that recruited 7002 middle-aged and older individuals (3318 men and 3684 women), egg consumption was found to be negatively associated with T2D risk in men, but not in women, suggesting the existence of gender differences [34]. Taken together, a meta-analysis that included above mentioned five cohort studies conducted in the US and Finland indicated that egg intake is not a risk factor for T2D [35].
The reported discrepancies among observational studies conducted in the US and other countries on the relationship between egg consumption and the risk of T2D could be caused by a number of reasons. In a meta-analysis of prospective cohort studies, Wallin et al. [29] suggested that the various food consumption habits among different countries and cultures could partly explain the heterogeneity in various reports. It has been widely reported that obesity and T2D is correlated with high intake of red meat, fat, and sugar in the diet [36,37], which is characterized as a common dietary pattern in the US population [38]. In fact, Sabate et al. [22] found that there was a significant interaction between meat and egg intake in a cohort study conducted with 55,851 participants in the US. Furthermore, the impact of socioeconomic factors including political environment, culture, income and stress on the prevalence of T2D might contribute to the inconsistency of the results among different countries [39,40]. It was well-established that social determinants such as income, education, housing, and accessibility to nutritious foods are critical players in the development of T2D [41,42]. In addition, alcohol intake and the use of tobacco and drugs are risk factors of T2D [43,44]. In particular, chronic stress, which leads to increases in blood pressure, blood glucose concentration and cortisol level as a consequence of allostatic overload [45], has been closely associated with the increased risk of T2D [46]. Other limitations of the observational studies are the unexcluded environmental factors and selection bias, and the uncontrolled confounding variables [47]. Also, the cooking methods—such as frying, boiling, and steaming—may impact overall nutritional profile and metabolism of egg intake [48,49]. Although cholesterol was once hypothesized as a risk factor for cardiovascular disease, accumulating evidence revoke the hypothesis [27,30,50]; the 2015–2020 Dietary Guidelines for Americans thus removed the limit of cholesterol intake. Currently, there is no consensus on the recommendation of egg intake for T2D patients, however, one egg a day is believed to be safe [51].
Unlike the observational studies, the short-term clinical studies have shown that high egg consumption is correlated with significant improvements on blood lipid level, cholesterol profile, insulin sensitivity, or hyperglycemia [52,53], suggesting higher egg consumption might be beneficial on insulin resistance in T2D. The effect of high-egg consumption on insulin resistance, glucose metabolism, and cholesterol level has been investigated in interventional studies such as randomized clinical trials [52,53,54] and the results were summarized in Table 1. Metabolic syndrome characterized by obesity, hyperlipidemia, hypertension, insulin resistance, and chronic inflammation is believed to be a predictor of T2D [55,56]. To investigate the effects of daily egg intake on lipoprotein metabolism and insulin resistance along with carbohydrate restriction in subjects with metabolic syndrome, Blesso et al. [52] conducted a randomized, single-blind, parallel designed study for 12 weeks to compare the meals containing three whole eggs or yolk-free egg substitute. Although both whole egg group and egg substitute group significantly resulted in improvement on very-low-density lipoprotein (VLDL) particle size, atherogenic lipoprotein subclasses, and oxidized low-density lipoprotein (LDL), whole egg group was shown to have greater increase in high-density lipoprotein cholesterol (HDL-C) and large HDL particles and higher reduction in total VLDL and medium VLDL particles than egg substitute group. Furthermore, plasma insulin concentration and insulin resistance were significantly reduced in whole egg group only [52], Which suggests that egg diet might be effective in managing metabolic syndrome. In addition, the inclusion of whole eggs provided additional benefits in improving insulin resistance compared to yolk-free egg diet [52], indicating the contribution of egg yolk components to this effect. Similarly, 65 subjects with T2D or impaired glucose tolerance were randomized to receive two low-energy diets, one with high protein and high cholesterol content plus two eggs per day, another is high protein-low cholesterol diet containing 100g of lean animal protein. After 12 weeks, diet containing 2 eggs/day did not change the blood lipid profiles of T2D subjects, while HDL-C was improved [53]. In another study conducted by Ratliff et al. in healthy men, one week of daily consumption of eggs for breakfast resulted in reduced energy intake and lower plasma glucose and insulin concentrations compared to a bagel breakfast [57]. A similar result was reported in a randomized, single-blind, crossover trial (n = 34) in adults with T2D, which showed that the intake of 2 eggs/day for 12 weeks significantly reduced body weight, waist circumference, visceral fat rating, and percent body fat [58]. The inclusion of eggs in the diet did not change the glycemic hemoglobin A1c (HbA1c) and homeostasis model assessment-insulin resistance (HOMA-IR). However, the exclusion of eggs significantly increased insulin resistance [58]. Also, the significant changes in weight and lipid metabolism suggest that egg might be beneficial in the context of metabolic syndrome [58]. On the contrary, several other clinical trials indicate that egg consumption may not alter metabolic biomarkers associated with T2D [54,59]. Fuller et al. reported that the intake of ≥12 eggs/week for 3 months did not significantly change glycemic response and blood concentrations of total cholesterol, HDL-C, LDL-C, and total triglycerides compared to low-egg diet group (<2eggs/week) in subjects with prediabetes or T2D [54]. This study was revalidated in a longer term (12 months) by the same research group [59]. There was no significant difference between high-egg (≥12 eggs/week) and low-egg diet (<2 eggs/week) in plasma glucose, glycated hemoglobin, 1,5-anhydroglucitol, serum lipids, inflammatory cytokines, oxidative stress, and adiponectin for 12 months [59]. However, the decrease in plasma glucose concentration and HbA1c were higher in high-egg group compared to low-egg group without statistical significance, which is probably due to the mixed subjects with prediabetes and T2D and the lack of a control group who does not consume an egg diet. Although controversial data exists, the current clinical studies provide promising evidence that egg diets ameliorate the risk of T2D. Since the current studies were mostly conducted for short term duration, further studies investigating prolonged consumption of eggs are warranted in individuals with insulin resistance or T2D.
3. Egg Components and T2D
3.1. Egg White Hydrolysate (EWH)
EWH produced using alcalase, flavourzyme, neutrase, trypsin, pepsin, pancreatin, and peptidase all showed in vitro activities against oxidative stress and inflammation [61], which are closely inter-connected processes involved in the onset and development of T2D and the progression of complications [62]. Oxidative stress is the result of the overproduction of intracellular reactive oxygen species (ROS) and reactive nitrogen species that damage lipids, proteins, and DNA [63]. The exposure of high concentration of glucose (i.e., 30 mM) causes ROS production through mitochondria pathway in pancreatic β-cells, which in turn inhibit glucose-stimulated insulin secretion (GSIS) [64,65]. Glucose also induces ROS generation and apoptosis in podocytes, whose degeneration predicts nephropathy in the context of T2D [66]. ROS was shown to induce insulin resistance in adipose tissue, skeletal muscle, and hepatocytes by impairing insulin signaling [67,68,69]. Furthermore, there is an association between oxidative stress and negative prognosis of diabetic retinopathy in both animal model and human patients [70]. Prolonged oxidative stress leads to chronic inflammation, while inflammation can induce oxidative stress [71]. Inflammation is the body’s protective response in the purpose of fixing an injury, infection, or irritation, which requires adequate balance between a broad spectrum of mediators, including vasoactive amines, complements, cytokines, chemokines, and eicosanoids [72]. The blood levels of pro-inflammatory cytokines and chemokines such as interleukin (IL)-1, IL-6, IL-18, tumor necrosis factor (TNF)-α, and monocyte chemotactic protein (MCP)-1 were reported to be elevated in T2D patients compared to normal subjects [73,74,75]. Adipose tissue inflammation is believed to play a crucial role in the development of impaired insulin secretion and sensitivity in T2D [76]. TNF-α and IL-1β are known to trigger apoptosis of islets [77], suppress GSIS in both islets and pancreatic beta-cells [78,79], and inhibit insulin signaling in adipose, muscle and liver cells and tissues mainly by suppressing the phosphorylation of insulin receptor, insulin receptor substrate (IRS)-1 and Akt (protein kinase B) [80,81,82,83,84]. Higher egg consumption is associated with lower concentrations of blood pro-inflammatory cytokines in T2D patients [53,85], although egg contains both anti- and pro-inflammatory components, such as anti-inflammatory lutein and zeaxanthin, pro-inflammatory cholesterol, and phospholipids being both anti- and pro-inflammatory [6]. Egg proteins, which are mostly in egg white, are generally considered to be anti-inflammatory, unless inducing an allergic reaction [6].
EWH was also reported to have bile acid-binding, angiotensin I-converting enzyme (ACE)-inhibitory and dipeptidyl peptidase 4 (DPP-4)-inhibitory activities in different potency based on in vitro experiments [61]. ACE is an enzyme that hydrolyzes angiotensin I to produce angiotensin II, which elevates blood pressure, promotes inflammation, and plays a role in the development of insulin resistance [86,87]. DPP-4 is an enzyme degrading incretin hormones such as glucagon-like peptide 1 (GLP-1) [88], which is secreted by intestinal L-cells in response to nutrient load and is known to promote insulin secretion from pancreatic β-cells, preserve β-cell proliferation and regeneration, and inhibit glucagon production [89].
There are some in vivo studies indicating that EWH might be beneficial in the context of T2D (Table 2). In spontaneous T2D mice with moderate obesity, eight weeks feeding of protease-produced EWH resulted in an improved glucose tolerance as shown in oral glucose tolerance test and intraperitoneal glucose tolerance test, which was accompanied with a lower plasma insulin concentration, suggesting both insulin secretion and sensitivity could be altered [90]. In high-fat diet (HFD)-fed rats, oral administration of pepsin-prepared EWH for 6 weeks was shown to decrease fat pad mass, increase lean mass, and alleviate glucose intolerance and insulin resistance, which were accompanied with enhanced Akt phosphorylation in liver, muscle, and fat tissues [91]. The plasma concentrations of IL-1α, IL-β, and MCP-1 in HFD-fed rats were also reduced by EWH supplementation [91]. EWH obtained from protease digestion significantly improved the fasting blood glucose concentration and HOMA-IR in non-obese spontaneous diabetic rats, without altering serum levels of insulin and adiponectin, DPP-4 activity, and homeostasis model assessments-insulin secretion [92]. However, in rats fed with a high-fat and high-sucrose diet, the same EWH had no effect on serum glucose and insulin concentrations [93]. Feeding with alcalase-produced EWH for 15 weeks was reported to protect against renovascular damage in obese T2D rats, which was accompanied with suppressed renal mRNA expression of TNF-α, without altering blood GLP-1 and glucose concentrations [94], suggesting the effect may not be dependent on its DPP-4 inhibiting activity. These results indicate that the effect of EWH on glucose and insulin metabolism and T2D-associated inflammation may be dependent on the preparation methods and the obese condition associated with diabetes. However, it should be noted that there is big variation in EWH dose (highest/lowest = ~10) in the current available studies [90,91,92,93,94].
The mechanism of EWH’s benefit on T2D is largely unknown. However, there are some in vitro studies supporting the insulin mimetic or sensitizing effects of EWH. Adipose tissue dysfunction plays a critical role in the development of insulin resistance and impaired metabolic homeostasis in T2D [95]. The differentiation of 3T3-F442A mouse pre-adipocytes was shown to be promoted by EWH (prepared by thermoase and pepsin) as evidenced by upregulated lipid accumulation and adiponectin production, possibly by upregulating the protein expression of peroxisome proliferator associated receptor gamma (PPAR)-γ and CCAAT/enhancer-binding protein alpha [96]. EWH treatment also increased the phosphorylation of extracellular signal regulated kinase 1/2 and attenuated c-Jun phosphorylation in 3T3-F442A cells, which was associated with decreased COX-2 expression, a critical regulator of inflammatory pathway. In addition, EWH treatment potentiated Akt phosphorylation induced by insulin [96]. In rat skeletal muscle cells L6, EWH were reported to improve TNF-α-impaired glucose uptake in response to insulin by promoting insulin signaling [97].
3.2. Lutein and Zeaxanthin
Lutein and zeaxanthin are carotenoid with similar structures to pre-vitamin A (β-carotene), which are concentrated at the macula as the primary pigment molecules [98]. Carotenoids are known for their potent antioxidant activity [99]. Egg yolk contains high level of lutein and zeaxanthin (~143 and 94 μg/yolk, respectively) [100], higher than most fruit and vegetables [101]. The serum concentrations of lutein and zeaxanthin increased 26 and 38% respectively after consuming 1 egg/day for 5 weeks in individuals aged >60y, without changes on concentrations of total cholesterol, LDL-C, HDL-C, and triglyceride in serum [100].
Lutein and zeaxanthin function as the major pigment molecules in retina [98]. The macular pigment optical density has been shown to be significantly lower in T2D patients than both type-1 diabetes subjects and normal individuals, although the level was comparable between T2D patients with or without retinopathy [102]. The lutein and zeaxanthin blood concentration in T2D patients with retinopathy was significantly lower than normal subjects [103]. In addition, in a cross-sectional study that involved 111 T2D patients, the plasma concentration of non-pro-vitamin A carotenoids, including lycopene, lutein, and zeaxanthin, was found to be significantly lower in patients with retinopathy than subjects without retinopathy. Furthermore, the ratio of carotenoids that are not pro-vitamin A to the ones that are pro-vitamin A in plasma was negatively associated with the risk of diabetic retinopathy [104].
There are some in vivo studies indicating possible benefits of lutein on insulin resistance and secretion. In rats fed with high-fat diet, lutein administration by gavage for 45 days attenuated hepatic insulin resistance, possibly by increasing the expression of IRS-2, phosphatidylinositol 3-kinase (PI3K), and glucose transporter (GLUT)-4 in liver at both transcription and translation levels [105]. The expression of PPAR-α and sirtuin 1 were also increased, which are players in insulin signaling as well [105]. Lutein and zeaxanthin treatment for eight weeks significantly reduced serum insulin concentration in high-fat diet-fed rats [106], indicating the modulating effect on insulin secretion.
The possible protecting role of lutein and zeaxanthin against diabetic retinopathy has been suggested by both animal and human studies. Lutein and zeaxanthin were shown to attenuate the oxidative damage of retina in high-fat diet-fed rats which develop obesity and insulin resistance, as evidenced by reduced malondialdehyde concentration and increased activity of antioxidant enzymes in retina [106]. Lutein treatment improved the result of electroretinogram test, reduced oxidative stress, and suppressed nuclear factor kappa B (NF-κB) activity in retina of diabetic rats induced by alloxan [107], which is a toxic glucose analogues causing β-cell damage through generating free radicals [108]. Zeaxanthin was reported to have similar protective property on the development of diabetic retinopathy as well. In diabetic rats induced by streptozotocin—which is another β-cell damaging glucose analogue mainly acting by inducing DNA damage [108]—zeaxanthin supplementation inhibited the levels of lipid peroxide, oxidatively modified DNA, electron transport complex III, nitrotyrosine, and mitochondrial superoxide dismutase in the retina, which were accompanied with reduced retinal expression of vascular endothelial growth factor and intercellular adhesion molecule-1 [109]. In db/db diabetic mice, dietary supplementation of wolfberry which has high lutein and zeaxanthin content was reported to restore the thinned retina, especially the inner nuclear and photoreceptor layers, and protect the integrity of the retinal pigment epithelia by improving mitochondria function and attenuating hypoxia, oxidative stress, and ER dysfunction [110,111]. The protecting effect of lutein on retina in the context of T2D was shown to be comparable with docosahexaenoic acid in terms of oxidative stress, apoptosis, thickness of the outer and inner nuclear layers, and electroretinogram in diabetic rats [112]. In T2D patients with diabetic retinopathy, three-month supplementation of lutein and zeaxanthin significantly improved visual acuity, increased contrast sensitivity, and decreased fovea thickness [103]. These evidence suggests that dietary lutein and zeaxanthin supplementation might be a promising strategy to alleviate the development of diabetic retinopathy, which requires further investigation.
3.3. Choline
Choline is a serine-derived water-soluble amine and is the build block of phosphatidylcholine and sphingomyelin, which are essential membrane phospholipid and precursors of second messengers diacylglycerol and ceramide. Choline is also a precursor of an crucial neurotransmitter acetylcholine, which is involved in voluntary muscle movement and cognitive function [113]. Another important physiological function of choline is that it is required for the transport of triglyceride in lipoprotein from liver [114].
Eggs, liver, and peanuts are good food sources of choline. The choline content in whole egg is ~100 mg/egg, mainly in egg yolk [115,116]. There are some evidence showing the beneficial effect of choline on glucose and insulin metabolism. In mice with metabolic syndrome, muscle synthesis of fatty acid and triglyceride was reduced by choline dietary supplementation, whereas glycogen generation was increased. In addition, phosphorylation of IRS-1 and Akt in muscle was enhanced by choline [117]. In a study that involved 2394 adults from Newfoundland, the dietary choline intake was negatively associated with blood concentrations of fasting glucose and insulin and HOMA-IR in both males and females with age, total calorie intake, and physical activity level being controlled [118]. Choline was also reported to aid in the delivery of insulin ingested orally. An ionic liquid consist of choline and geranate was shown to improve oral insulin delivery and result in a significantly reduced blood glucose level for up to 12 hours in rats [119], which were attributed to enhanced paracellular transport and reduced enzymatic degradation of insulin.
However, there are also studies showing contradictory results. Phosphatidylethanolamine N-methyltransferase-deficient mice, which display impaired choline de novo synthesis, were protected from high-fat diet-induced obesity and insulin resistance [120]. In addition, a choline-deficient diet led to an improved glucose tolerance and less weight gain in wild-type mice [120]. Similar attenuation on weight gain and improvement on glucose tolerance and insulin resistance by a choline-deficient diet were seen in both ob/ob obese mice and high-fat diet-fed wild-type mice [121]. In a human study that involved three prospective cohorts with 203,308 subjects (both male and female) without diabetes, cardiovascular disease, and cancer [122], a valid food-frequency questionnaire was used to assess the dietary intake of phosphatidylcholine, which can be degraded to choline by gut microbes. The results showed that higher phosphatidylcholine intake was associated with increased risk of T2D [122].
4. Egg-Derived Peptides and T2D
4.1. Alpha-Glucosidase Inhibitory Peptides
Although individual variation and abdominal discomfort have been reported, synthetic α-glucosidase inhibitors, such as acarbose and voglibose, were shown to be beneficial against postprandial hyperglycemia in diabetic individuals by inhibiting α-glucosidase, which hydrolyzes carbohydrate and release monosaccharides for absorption in the small intestine [123,124]. Both acarbose and voglibose were effective on improving glucose turnover in T2D patients who were taking insulin and metformin and blood glucose levels were not adequately controlled [125], indicating α-glucosidase inhibitors could be valuable supplements for T2D patients. It was also reported that voglibose diet supplementation augmented GLP-1 secretion in both healthy people and mice with T2D, which is attributed to the delaying effect on carbohydrate absorption [126,127]. However, acarbose was not shown to increase GLP-1 secretion in patients with T2D [128], indicating α-glucosidase inhibitors may have distinct potency on promoting GLP-1 secretion in different health conditions. Thus, food-derived α-glucosidase inhibitors might be attractive options replacing synthetic ones for the management of T2D. Eight peptides with α-glucosidase inhibitory activity have been identified in egg albumin with peptide KLPGF being the most potent one, which was shown to have comparable potency as acarbose [129]. Another egg white derived peptide RVPSLM is ~3-fold more potent than acarbose in glucosidase inhibition [130]. Peptide VTGRFAGHPAAQ with high α-glucosidase inhibitory activity was identified from egg yolk protein [131]. However, effect of α-glucosidase inhibitory peptides from egg on glucose and insulin metabolism in the context of T2D requires further study.
4.2. ACE Inhibitory Peptides
In the context of T2D, angiotensin II, produced from angiotensin I by ACE, has been implicated to play a role in the development of insulin resistance. Angiotensin II was reported to inhibit insulin/ PI3K/Akt signaling, induce oxidative stress by activating NADPH oxidases, and upregulate inflammation by activation of NF-κB [86], which is known to initiate the transcription of pro-inflammatory genes including cytokines, chemokines, and adhesion molecules [132]. The action of angiotensin II is believed to be mainly through a G-protein coupled receptor, angiotensin II type 1 receptor [133]. ACE inhibitors, traditionally used for anti-hypertension purpose, have been reported to exert favorable effects on kidney, heart and eye functions in T2D patients [134].
Milk derived ACE inhibitory tripeptides, IPP (Ile-Pro-Pro) and VPP (Val-Pro-Pro), induced similar adipogenic differentiation to insulin, which was accompanied with restored adipokine levels and reduced activation of NF-κB [135]. An ACE inhibitory tripeptide, IRW (Ile-Arg-Trp), which is derived from egg white ovotransferrin, has been indicated to be beneficial against insulin resistance. In fully differentiated L6 myoblasts, IRW were reported to restore TNF-α-impaired insulin- stimulated glucose uptake by promoting phosphorylation of IRS-1 tyrosine residue and Akt, which were accompanied with decreased phosphorylation of p38 and c-Jun N-terminal kinases [97]. IRW in vitro treatment also reversed angiotensin II-impaired insulin-stimulated glucose uptake in L6 myoblasts, by reducing serine phosphorylation of IRS-1, increasing Akt phosphorylation, upregulating GLUT-4 translocation, decreasing expression of angiotensin II type 1 receptor, and inhibiting ROS generation [136]. The effect and mechanisms of ACE inhibitory peptides from eggs on diabetes largely remain to be elucidated.
4.3. DPP-4 Inhibitory Peptides
Several DPP-4 inhibitors (also known as gliptins) have been approved in the US, Europe, Japan, and South Korea to treat T2D, which are supposed to augment the bioavailability of incretin hormones, prolong the action of insulin, and thus provide benefits on postprandial glucose response [137]. Peptide LPQNIPPL originated from water-soluble extract of a gouda-type cheese was reported to have DPP-4 inhibitory activities in vitro, and improve glucose tolerance as shown in oral glucose tolerance test in healthy rats when administrated together with glucose by intraperitoneal injection [138]. In addition, oral administration of peptides with DPP-4 inhibitory activity derived from the porcine skin gelatin hydrolysates were demonstrated to improve glucose tolerance in diabetic rats in 21 and 42 days after streptozotocin injection [139]. Three peptides, YINQMPQKSREA, VTGRFAGHPAAQ, and YINQMPQKSRE, with DPP-4 inhibitory activity have been identified in egg yolk protein, with YINQMPQKSRE being the most active one (IC50 = 222.8 µg/mL) [131]. However, further studies elucidating the effect of egg-derived DPP-4 inhibitory peptides in the context of T2D are needed.
5. Concluding Remark
To date, the observational epidemiological evidence about the egg consumption and the risk of T2D is not consistent, which might be the result of different dietary pattern and socioeconomic factors. However, it has been indicated that there is association between higher egg consumption and improved blood lipid profile, insulin sensitivity, and glucose response in interventional clinical trials.
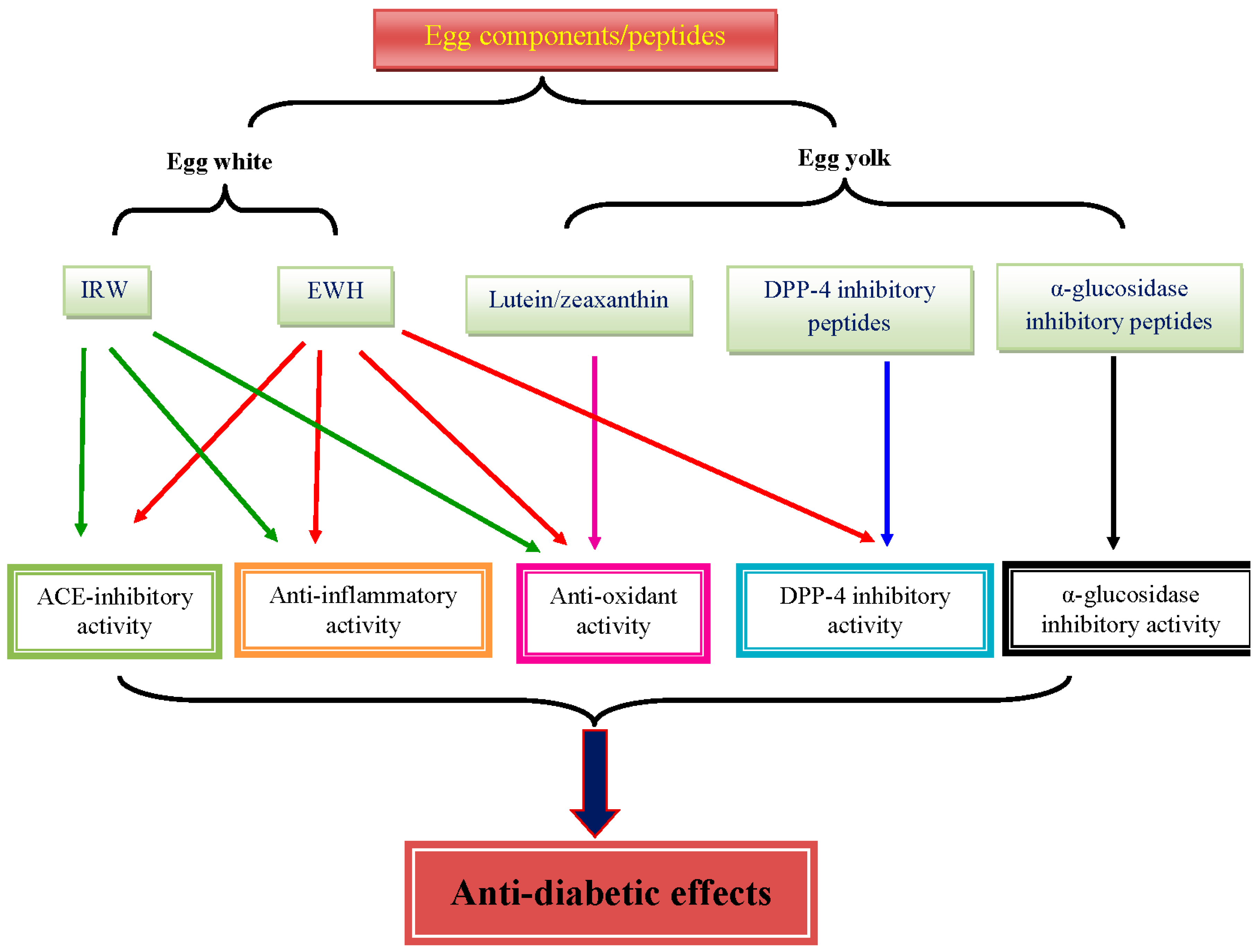
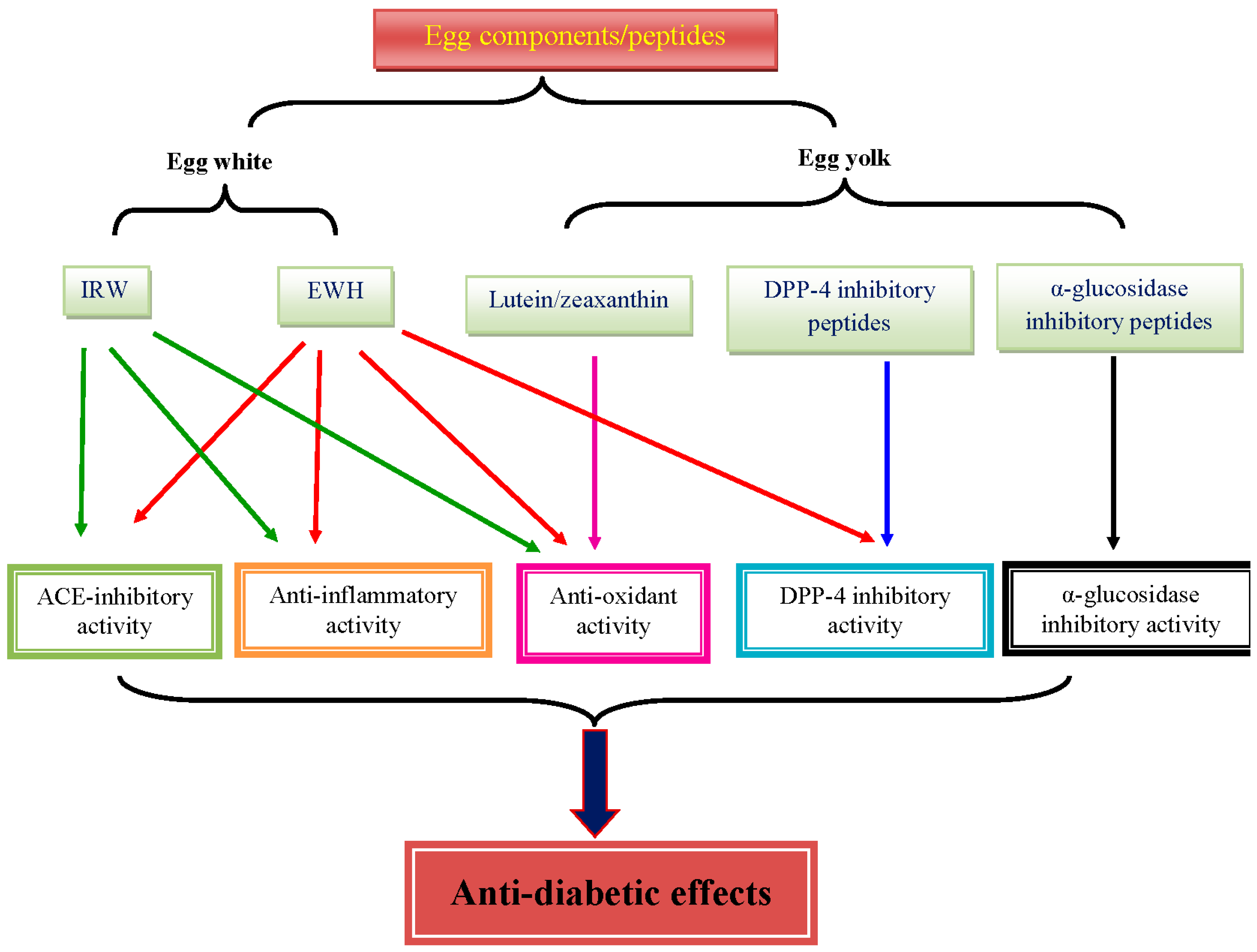
EWH, lutein, zeaxanthin, and ACE inhibitory tripeptides from egg have been shown to have some benefits against glucose and insulin intolerance, oxidative stress, and inflammation in the context of T2D (Figure 1). The effect of EWH seems to be related to the production process, which needs optimization. In addition, the role of α-glucosidase inhibitory peptides, DPP-4 inhibitory peptides, and choline from egg in T2D is poorly understood. Thus, more mechanistic studies are warranted to elucidate the role of each components of egg on T2D. Furthermore, the absorption and bioavailability of the egg components and egg-derived peptides largely remain to be addressed.
Author Contributions
Writing—original draft preparation, X.W., M.S., and C.M.; writing—review and editing, X.W. and J.W.; supervision, J.W.; funding acquisition, J.W.
Funding
This research was funded by Alberta Livestock and Meat Agency (ALMA); Egg Farmers of Canada; and Natural Sciences and Engineering Research Council (NSERC) of Canada.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kharroubi, A.T.; Darwish, H.M. Diabetes mellitus: The epidemic of the century. World J. Diabetes 2015, 6, 850–867. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Report on Diabetes; World Health Organization: Geneva, Switzerland, 2016; pp. 1–86. [Google Scholar]
- Abate, N.; Chandalia, M. Ethnicity and type 2 diabetes: Focus on Asian Indians. J. Diabetes Complicat. 2001, 15, 320–327. [Google Scholar] [CrossRef]
- Kubota, M.; Yoneda, M.; Maeda, N.; Ohno, H.; Oki, K.; Funahashi, T.; Shimomura, I.; Hattori, N. Westernization of lifestyle affects quantitative and qualitative changes in adiponectin. Cardiovasc. Diabetol. 2017, 16, 83. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, S.; Okubo, M.; Yoneda, M.; Jitsuiki, K.; Yamane, K.; Kohno, N. A comparison between Japanese-Americans living in Hawaii and Los Angeles and native Japanese: The impact of lifestyle westernization on diabetes mellitus. Biomed. Pharmacother. 2004, 58, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.J. Bioactive Egg Components and Inflammation. Nutrients 2015, 7, 7889–7913. [Google Scholar] [CrossRef] [PubMed]
- Zani, S.C.D.; Wu, J.P.; Chan, C.B. Egg and Soy-Derived Peptides and Hydrolysates: A Review of Their Physiological Actions against Diabetes and Obesity. Nutrients 2018, 10, 549. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Jahandideh, F.; Wu, J. Food-derived bioactive peptides on inflammation and oxidative stress. Biomed. Res. Int. 2014, 2014, 608979. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.K.; Koh, G.Y.; Rowling, M.J.; Schalinske, K.L. Whole Egg Consumption Prevents Diminished Serum 25-Hydroxycholecalciferol Concentrations in Type 2 Diabetic Rats. J. Agric. Food. Chem. 2016, 64, 120–124. [Google Scholar] [CrossRef]
- Saande, C.J.; Jones, S.K.; Hahn, K.E.; Reed, C.H.; Rowling, M.J.; Schalinske, K.L. Dietary Whole Egg Consumption Attenuates Body Weight Gain and Is More Effective than Supplemental Cholecalciferol in Maintaining Vitamin D Balance in Type 2 Diabetic Rats. J. Nutr. 2017, 147, 1715–1721. [Google Scholar] [CrossRef]
- Vang, A.; Singh, P.N.; Lee, J.W.; Haddad, E.H.; Brinegar, C.H. Meats, processed meats, obesity, weight gain and occurrence of diabetes among adults: Findings from Adventist Health Studies. Ann. Nutr. Metab. 2008, 52, 96–104. [Google Scholar] [CrossRef]
- Djousse, L.; Gaziano, J.M.; Buring, J.E.; Lee, I.M. Egg consumption and risk of type 2 diabetes in men and women. Diabetes Care 2009, 32, 295–300. [Google Scholar] [CrossRef]
- Djousse, L.; Kamineni, A.; Nelson, T.L.; Carnethon, M.; Mozaffarian, D.; Siscovick, D.; Mukamal, K.J. Egg consumption and risk of type 2 diabetes in older adults. Am. J. Clin. Nutr. 2010, 92, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Zazpe, I.; Beunza, J.J.; Bes-Rastrollo, M.; Basterra-Gortari, F.J.; Mari-Sanchis, A.; Martinez-Gonzalez, M.A.; Investigators, S.U.N.P. Egg consumption and risk of type 2 diabetes in a Mediterranean cohort; the sun project. Nutr. Hosp. 2013, 28, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Lajous, M.; Bijon, A.; Fagherazzi, G.; Balkau, B.; Boutron-Ruault, M.C.; Clavel-Chapelon, F. Egg and cholesterol intake and incident type 2 diabetes among French women. Br. J. Nutr. 2015, 114, 1667–1673. [Google Scholar] [CrossRef]
- Kurotani, K.; Nanri, A.; Goto, A.; Mizoue, T.; Noda, M.; Oba, S.; Sawada, N.; Tsugane, S.; Japan Public Health Center-based Prospective Study Group. Cholesterol and egg intakes and the risk of type 2 diabetes: The Japan Public Health Center-based Prospective Study. Br. J. Nutr. 2014, 112, 1636–1643. [Google Scholar] [CrossRef] [PubMed]
- Djousse, L.; Petrone, A.B.; Hickson, D.A.; Talegawkar, S.A.; Dubbert, P.M.; Taylor, H.; Tucker, K.L. Egg consumption and risk of type 2 diabetes among African Americans: The Jackson Heart Study. Clin. Nutr. 2016, 35, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, J.K.; Mursu, J.; Tuomainen, T.P.; Virtanen, H.E.; Voutilainen, S. Egg consumption and risk of incident type 2 diabetes in men: The Kuopio Ischaemic Heart Disease Risk Factor Study. Am. J. Clin Nutr. 2015, 101, 1088–1096. [Google Scholar] [CrossRef]
- Montonen, J.; Jarvinen, R.; Heliovaara, M.; Reunanen, A.; Aromaa, A.; Knekt, P. Food consumption and the incidence of type II diabetes mellitus. Eur. J. Clin. Nutr. 2005, 59, 441–448. [Google Scholar] [CrossRef]
- Ericson, U.; Hellstrand, S.; Brunkwall, L.; Schulz, C.A.; Sonestedt, E.; Wallstrom, P.; Gullberg, B.; Wirfalt, E.; Orho-Melander, M. Food sources of fat may clarify the inconsistent role of dietary fat intake for incidence of type 2 diabetes. Am. J. Clin. Nutr. 2015, 101, 1065–1080. [Google Scholar] [CrossRef]
- Shi, Z.; Yuan, B.; Zhang, C.; Zhou, M.; Holmboe-Ottesen, G. Egg consumption and the risk of diabetes in adults, Jiangsu, China. Nutrition 2011, 27, 194–198. [Google Scholar] [CrossRef]
- Sabate, J.; Burkholder-Cooley, N.M.; Segovia-Siapco, G.; Oda, K.; Wells, B.; Orlich, M.J.; Fraser, G.E. Unscrambling the relations of egg and meat consumption with type 2 diabetes risk. Am. J. Clin. Nutr. 2018, 108, 1121–1128. [Google Scholar] [CrossRef] [PubMed]
- Tamez, M.; Virtanen, J.K.; Lajous, M. Egg consumption and risk of incident type 2 diabetes: A dose-response meta-analysis of prospective cohort studies. Br. J. Nutr. 2016, 115, 2212–2218. [Google Scholar] [CrossRef] [PubMed]
- Radzeviciene, L.; Ostrauskas, R. Egg consumption and the risk of type 2 diabetes mellitus: A case-control study. Publ. Health Nutr. 2012, 15, 1437–1441. [Google Scholar] [CrossRef] [PubMed]
- Weggemans, R.M.; Zock, P.L.; Katan, M.B. Dietary cholesterol from eggs increases the ratio of total cholesterol to high-density lipoprotein cholesterol in humans: A meta-analysis. Am. J. Clin. Nutr. 2001, 73, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Richard, C.; Cristall, L.; Fleming, E.; Lewis, E.D.; Ricupero, M.; Jacobs, R.L.; Field, C.J. Impact of Egg Consumption on Cardiovascular Risk Factors in Individuals with Type 2 Diabetes and at Risk for Developing Diabetes: A Systematic Review of Randomized Nutritional Intervention Studies. Can. J. Diabetes 2017, 41, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.Y.; Xun, P.; Nakamura, Y.; He, K. Egg consumption in relation to risk of cardiovascular disease and diabetes: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2013, 98, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, C.; Zhou, X.; Li, L. Egg consumption and risk of cardiovascular diseases and diabetes: A meta-analysis. Atherosclerosis 2013, 229, 524–530. [Google Scholar] [CrossRef]
- Wallin, A.; Forouhi, N.G.; Wolk, A.; Larsson, S.C. Egg consumption and risk of type 2 diabetes: A prospective study and dose-response meta-analysis. Diabetologia 2016, 59, 1204–1213. [Google Scholar] [CrossRef]
- Djousse, L.; Gaziano, J.M. Egg consumption in relation to cardiovascular disease and mortality: The Physicians’ Health Study. Am. J. Clin. Nutr. 2008, 87, 964–969. [Google Scholar] [CrossRef]
- Djousse, L.; Khawaja, O.A.; Gaziano, J.M. Egg consumption and risk of type 2 diabetes: A meta-analysis of prospective studies. Am. J. Clin. Nutr. 2016, 103, 474–480. [Google Scholar] [CrossRef]
- Guo, J.; Hobbs, D.A.; Cockcroft, J.R.; Elwood, P.C.; Pickering, J.E.; Lovegrove, J.A.; Givens, D.I. Association between egg consumption and cardiovascular disease events, diabetes and all-cause mortality. Eur. J. Nutr. 2018, 57, 2943–2952. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, H.E.K.; Koskinen, T.T.; Voutilainen, S.; Mursu, J.; Tuomainen, T.P.; Kokko, P.; Virtanen, J.K. Intake of different dietary proteins and risk of type 2 diabetes in men: The Kuopio Ischaemic Heart Disease Risk Factor Study. Br. J. Nutr. 2017, 117, 882–893. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, J. Egg consumption is associated with a lower risk of type 2 diabetes in middle-aged and older men. Nutr. Res. Pract. 2018, 12, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Xu, Q.; Jiang, R.; Han, T.; Sun, C.; Na, L. Dietary Protein Consumption and the Risk of Type 2 Diabetes: A Systematic Review and Meta-Analysis of Cohort Studies. Nutrients 2017, 9, 982. [Google Scholar] [CrossRef] [PubMed]
- Pan, A.; Sun, Q.; Bernstein, A.M.; Schulze, M.B.; Manson, J.E.; Willett, W.C.; Hu, F.B. Red meat consumption and risk of type 2 diabetes: 3 cohorts of US adults and an updated meta-analysis. Am. J. Clin. Nutr. 2011, 94, 1088–1096. [Google Scholar] [CrossRef] [PubMed]
- Erber, E.; Hopping, B.N.; Grandinetti, A.; Park, S.Y.; Kolonel, L.N.; Maskarinec, G. Dietary patterns and risk for diabetes: The multiethnic cohort. Diabetes Care 2010, 33, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Kenfield, S.A.; Van Blarigan, E.L.; Batista, J.L.; Sesso, H.D.; Ma, J.; Stampfer, M.J.; Chavarro, J.E. Dietary patterns after prostate cancer diagnosis in relation to disease-specific and total mortality. Cancer Prev. Res. 2015, 8, 545–551. [Google Scholar] [CrossRef]
- Grzywacz, J.G.; Arcury, T.A.; Ip, E.H.; Nguyen, H.T.; Saldana, S.; Reynolds, T.; Bell, R.A.; Kirk, J.K.; Quandt, S.A. Cultural basis for diabetes-related beliefs among low- and high-education African American, American Indian, and white older adults. Ethn. Dis. 2012, 22, 466–472. [Google Scholar]
- Dendup, T.; Feng, X.; Clingan, S.; Astell-Burt, T. Environmental Risk Factors for Developing Type 2 Diabetes Mellitus: A Systematic Review. Int. J. Environ. Res. Public Health 2018, 15, 78. [Google Scholar] [CrossRef]
- Clark, M.L.; Utz, S.W. Social determinants of type 2 diabetes and health in the United States. World J. Diabetes 2014, 5, 296–304. [Google Scholar] [CrossRef]
- Walker, R.J.; Smalls, B.L.; Egede, L.E. Social determinants of health in adults with type 2 diabetes—Contribution of mutable and immutable factors. Diabetes Res. Clin. Pract. 2015, 110, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.; Nielsen, M.; Fox, M.H. Understanding the social factors that contribute to diabetes: A means to informing health care and social policies for the chronically ill. Perm. J. 2013, 17, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Shu, X.O.; Li, H.; Cai, H.; Liu, Q.; Zheng, W.; Xiang, Y.B.; Villegas, R. Physical activity, smoking, and alcohol consumption in association with incidence of type 2 diabetes among middle-aged and elderly Chinese men. PLoS ONE 2013, 8, e77919. [Google Scholar] [CrossRef]
- Juster, R.P.; McEwen, B.S.; Lupien, S.J. Allostatic load biomarkers of chronic stress and impact on health and cognition. Neurosci. Biobehav. Rev. 2010, 35, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Mooy, J.M.; de Vries, H.; Grootenhuis, P.A.; Bouter, L.M.; Heine, R.J. Major stressful life events in relation to prevalence of undetected type 2 diabetes: The Hoorn Study. Diabetes Care 2000, 23, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Boyko, E.J. Observational research–opportunities and limitations. J. Diabetes Complicat. 2013, 27, 642–648. [Google Scholar] [CrossRef] [PubMed]
- Schlatterer, J.; Breithaupt, D.E. Xanthophylls in commercial egg yolks: Quantification and identification by HPLC and LC-(APCI)MS using a C30 phase. J. Agric. Food. Chem. 2006, 54, 2267–2273. [Google Scholar] [CrossRef] [PubMed]
- Remanan, M.K.; Wu, J. Antioxidant activity in cooked and simulated digested eggs. Food Funct. 2014, 5, 1464–1474. [Google Scholar] [CrossRef]
- Rong, Y.; Chen, L.; Zhu, T.; Song, Y.; Yu, M.; Shan, Z.; Sands, A.; Hu, F.B.; Liu, L. Egg consumption and risk of coronary heart disease and stroke: Dose-response meta-analysis of prospective cohort studies. BMJ 2013, 346, e8539. [Google Scholar] [CrossRef]
- Geiker, N.R.W.; Larsen, M.L.; Dyerberg, J.; Stender, S.; Astrup, A. Egg consumption, cardiovascular diseases and type 2 diabetes. Eur. J. Clin. Nutr. 2018, 72, 44–56. [Google Scholar] [CrossRef]
- Blesso, C.N.; Andersen, C.J.; Barona, J.; Volek, J.S.; Fernandez, M.L. Whole egg consumption improves lipoprotein profiles and insulin sensitivity to a greater extent than yolk-free egg substitute in individuals with metabolic syndrome. Metab. Clin. Exp. 2013, 62, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Pearce, K.L.; Clifton, P.M.; Noakes, M. Egg consumption as part of an energy-restricted high-protein diet improves blood lipid and blood glucose profiles in individuals with type 2 diabetes. Br. J. Nutr. 2011, 105, 584–592. [Google Scholar] [CrossRef]
- Fuller, N.R.; Caterson, I.D.; Sainsbury, A.; Denyer, G.; Fong, M.; Gerofi, J.; Baqleh, K.; Williams, K.H.; Lau, N.S.; Markovic, T.P. The effect of a high-egg diet on cardiovascular risk factors in people with type 2 diabetes: The Diabetes and Egg (DIABEGG) study-a 3-mo randomized controlled trial. Am. J. Clin. Nutr. 2015, 101, 705–713. [Google Scholar] [CrossRef]
- Tenenbaum, A.; Fisman, E.Z.; Motro, M. Metabolic syndrome and type 2 diabetes mellitus: Focus on peroxisome proliferator activated receptors (PPAR). Cardiovasc. Diabetol. 2003, 2, 4. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.A.; Lee, J.H.; Lim, S.Y.; Ha, H.S.; Kwon, H.S.; Park, Y.M.; Lee, W.C.; Kang, M.I.; Yim, H.W.; Yoon, K.H.; et al. Metabolic syndrome as a predictor of type 2 diabetes, and its clinical interpretations and usefulness. J. Diabetes Investig. 2013, 4, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Ratliff, J.; Leite, J.O.; de Ogburn, R.; Puglisi, M.J.; VanHeest, J.; Fernandez, M.L. Consuming eggs for breakfast influences plasma glucose and ghrelin, while reducing energy intake during the next 24 hours in adult men. Nutr. Res. 2010, 30, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Njike, V.Y.; Ayettey, R.G.; Rajebi, H.; Treu, J.A.; Katz, D.L. Egg ingestion in adults with type 2 diabetes: Effects on glycemic control, anthropometry, and diet quality-a randomized, controlled, crossover trial. BMJ Open Diabetes Res. Care 2016, 4, e000281. [Google Scholar] [CrossRef]
- Fuller, N.R.; Sainsbury, A.; Caterson, I.D.; Denyer, G.; Fong, M.; Gerofi, J.; Leung, C.; Lau, N.S.; Williams, K.H.; Januszewski, A.S.; et al. Effect of a high-egg diet on cardiometabolic risk factors in people with type 2 diabetes: The Diabetes and Egg (DIABEGG) Study-randomized weight-loss and follow-up phase. Am. J. Clin. Nutr. 2018, 107, 921–931. [Google Scholar] [CrossRef]
- Grundy, S.M.; Brewer, H.B., Jr.; Cleeman, J.I.; Smith, S.C., Jr.; Lenfant, C.; American Heart, A.; National Heart, L.; Blood, I. Definition of metabolic syndrome: Report of the National Heart, Lung, and Blood Institute/American Heart Association conference on scientific issues related to definition. Circulation 2004, 109, 433–438. [Google Scholar] [CrossRef]
- Garcés-Rimón, M.; López-Expósito, I.; López-Fandiño, R.; Miguel, M. Egg white hydrolysates with in vitro biological multiactivities to control complications associated with the metabolic syndrome. Eur. Food Res. Technol. 2015, 242, 61–69. [Google Scholar] [CrossRef]
- Ma, X.; Chen, Z.; Wang, L.; Wang, G.; Wang, Z.; Dong, X.; Wen, B.; Zhang, Z. The Pathogenesis of Diabetes Mellitus by Oxidative Stress and Inflammation: Its Inhibition by Berberine. Front. Pharmacol. 2018, 9, 782. [Google Scholar] [CrossRef]
- Lichtenberg, D.; Pinchuk, I. Oxidative stress, the term and the concept. Biochem. Biophys. Res. Commun. 2015, 461, 441–444. [Google Scholar] [CrossRef]
- Hou, N.; Torii, S.; Saito, N.; Hosaka, M.; Takeuchi, T. Reactive oxygen species-mediated pancreatic beta-cell death is regulated by interactions between stress-activated protein kinases, p38 and c-Jun N-terminal kinase, and mitogen-activated protein kinase phosphatases. Endocrinology 2008, 149, 1654–1665. [Google Scholar] [CrossRef]
- Sakai, K.; Matsumoto, K.; Nishikawa, T.; Suefuji, M.; Nakamaru, K.; Hirashima, Y.; Kawashima, J.; Shirotani, T.; Ichinose, K.; Brownlee, M.; et al. Mitochondrial reactive oxygen species reduce insulin secretion by pancreatic beta-cells. Biochem. Biophy. Res. Commun. 2003, 300, 216–222. [Google Scholar] [CrossRef]
- Susztak, K.; Raff, A.C.; Schiffer, M.; Bottinger, E.P. Glucose-induced reactive oxygen species cause apoptosis of podocytes and podocyte depletion at the onset of diabetic nephropathy. Diabetes 2006, 55, 225–233. [Google Scholar] [CrossRef]
- Paglialunga, S.; Ludzki, A.; Root-McCaig, J.; Holloway, G.P. In adipose tissue, increased mitochondrial emission of reactive oxygen species is important for short-term high-fat diet-induced insulin resistance in mice. Diabetologia 2015, 58, 1071–1080. [Google Scholar] [CrossRef]
- Di Meo, S.; Iossa, S.; Venditti, P. Skeletal muscle insulin resistance: Role of mitochondria and other ROS sources. J. Endocrinol. 2017, 233, R15–R42. [Google Scholar] [CrossRef]
- Nakamura, S.; Takamura, T.; Matsuzawa-Nagata, N.; Takayama, H.; Misu, H.; Noda, H.; Nabemoto, S.; Kurita, S.; Ota, T.; Ando, H.; et al. Palmitate induces insulin resistance in H4IIEC3 hepatocytes through reactive oxygen species produced by mitochondria. J. Biol. Chem. 2009, 284, 14809–14818. [Google Scholar] [CrossRef]
- Arden, G.B.; Sivaprasad, S. Hypoxia and oxidative stress in the causation of diabetic retinopathy. Curr. Diabetes Rev. 2011, 7, 291–304. [Google Scholar] [CrossRef]
- Khansari, N.; Shakiba, Y.; Mahmoudi, M. Chronic inflammation and oxidative stress as a major cause of age-related diseases and cancer. Recent Pat. Inflamm. Allergy Drug Discov. 2009, 3, 73–80. [Google Scholar] [CrossRef]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Chen, F.Q.; Wang, J.; Liu, X.B.; Ma, X.Y.; Zhang, X.B.; Huang, T.; Ma, D.W.; Wang, Q.Y. Levels of inflammatory cytokines in type 2 diabetes patients with different urinary albumin excretion rates and their correlation with clinical variables. J. Diabetes Res. 2013, 2013, 138969. [Google Scholar] [CrossRef]
- Pickup, J.C.; Chusney, G.D.; Thomas, S.M.; Burt, D. Plasma interleukin-6, tumour necrosis factor alpha and blood cytokine production in type 2 diabetes. Life Sci. 2000, 67, 291–300. [Google Scholar] [CrossRef]
- Zaharieva, E.; Kamenov, Z.; Velikova, T.; Tsakova, A.; El-Darawish, Y.; Okamura, H. Interleukin-18 serum level is elevated in type 2 diabetes and latent autoimmune diabetes. Endocr. Connect. 2018, 7, 179–185. [Google Scholar] [CrossRef]
- Abranches, M.V.; Oliveira, F.C.; Conceicao, L.L.; Peluzio, M.D. Obesity and diabetes: The link between adipose tissue dysfunction and glucose homeostasis. Nutr. Res. Rev. 2015, 28, 121–132. [Google Scholar] [CrossRef]
- Kim, S.; Kim, K.A.; Suk, K.; Kim, Y.H.; Oh, S.H.; Lee, M.K.; Kim, K.W.; Lee, M.S. Apoptosis of human islet cells by cytokines. Immune Netw. 2012, 12, 113–117. [Google Scholar] [CrossRef]
- Dunger, A.; Cunningham, J.M.; Delaney, C.A.; Lowe, J.E.; Green, M.H.; Bone, A.J.; Green, I.C. Tumor necrosis factor-alpha and interferon-gamma inhibit insulin secretion and cause DNA damage in unweaned-rat islets. Extent of nitric oxide involvement. Diabetes 1996, 45, 183–189. [Google Scholar] [CrossRef]
- Zhang, S.; Kim, K.H. TNF-alpha inhibits glucose-induced insulin secretion in a pancreatic beta-cell line (INS-1). FEBS Lett. 1995, 377, 237–239. [Google Scholar] [CrossRef]
- Ballak, D.B.; Stienstra, R.; Tack, C.J.; Dinarello, C.A.; van Diepen, J.A. IL-1 family members in the pathogenesis and treatment of metabolic disease: Focus on adipose tissue inflammation and insulin resistance. Cytokine 2015, 75, 280–290. [Google Scholar] [CrossRef]
- Borst, S.E. The role of TNF-alpha in insulin resistance. Endocrine 2004, 23, 177–182. [Google Scholar] [CrossRef]
- De Taeye, B.M.; Novitskaya, T.; McGuinness, O.P.; Gleaves, L.; Medda, M.; Covington, J.W.; Vaughan, D.E. Macrophage TNF-alpha contributes to insulin resistance and hepatic steatosis in diet-induced obesity. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E713–E725. [Google Scholar] [CrossRef]
- Jager, J.; Gremeaux, T.; Cormont, M.; Le Marchand-Brustel, Y.; Tanti, J.F. Interleukin-1beta-induced insulin resistance in adipocytes through down-regulation of insulin receptor substrate-1 expression. Endocrinology 2007, 148, 241–251. [Google Scholar] [CrossRef]
- Plomgaard, P.; Bouzakri, K.; Krogh-Madsen, R.; Mittendorfer, B.; Zierath, J.R.; Pedersen, B.K. Tumor necrosis factor-alpha induces skeletal muscle insulin resistance in healthy human subjects via inhibition of Akt substrate 160 phosphorylation. Diabetes 2005, 54, 2939–2945. [Google Scholar] [CrossRef]
- Ballesteros, M.N.; Valenzuela, F.; Robles, A.E.; Artalejo, E.; Aguilar, D.; Andersen, C.J.; Valdez, H.; Fernandez, M.L. One Egg per Day Improves Inflammation when Compared to an Oatmeal-Based Breakfast without Increasing Other Cardiometabolic Risk Factors in Diabetic Patients. Nutrients 2015, 7, 3449–3463. [Google Scholar] [CrossRef]
- Olivares-Reyes, J.A.; Arellano-Plancarte, A.; Castillo-Hernandez, J.R. Angiotensin II and the development of insulin resistance: Implications for diabetes. Mol. Cell. Endocrinol. 2009, 302, 128–139. [Google Scholar] [CrossRef]
- Young, C.N.; Davisson, R.L. Angiotensin-II, the Brain, and Hypertension: An Update. Hypertension 2015, 66, 920–926. [Google Scholar] [CrossRef]
- Brunton, S. GLP-1 receptor agonists vs. DPP-4 inhibitors for type 2 diabetes: Is one approach more successful or preferable than the other? Int. J. Clin. Pract. 2014, 68, 557–567. [Google Scholar] [CrossRef]
- Trujillo, J.M.; Nuffer, W.; Ellis, S.L. GLP-1 receptor agonists: A review of head-to-head clinical studies. Ther. Adv. Endocrinol. Metab. 2015, 6, 19–28. [Google Scholar] [CrossRef]
- Ochiai, M.; Azuma, Y. Egg White Hydrolysate Improves Glucose Tolerance in Type-2 Diabetic NSY Mice. J. Nutr. Sci. Vitaminol. 2017, 63, 422–429. [Google Scholar] [CrossRef]
- Jahandideh, F. Effects of Egg White Derived Peptides on Metabolic Syndrome Complications: Hypertension, Inflammation, and Insulin Resistance. Ph.D. Thesis, University of Alberta, Edmonton, AB, Canada, 2017. [Google Scholar]
- Ochiai, M.; Kuroda, T.; Matsuo, T. Increased muscular triglyceride content and hyperglycemia in Goto-Kakizaki rat are decreased by egg white hydrolysate. Int J. Food Sci. Nutr. 2014, 65, 495–501. [Google Scholar] [CrossRef]
- Ochiai, M.; Matsuo, T. Effect of egg white and its hydrolysate on stearoyl-CoA desaturase index and fat accumulation in rat tissues. Int. J. Food Sci. Nutr. 2014, 65, 948–952. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Landheer, S.; van Gilst, W.H.; van Amerongen, A.; Hammes, H.P.; Henning, R.H.; Deelman, L.E.; Buikema, H. Attenuation of renovascular damage in Zucker diabetic fatty rat by NWT-03, an egg protein hydrolysate with ACE- and DPP4-inhibitory Activity. PLoS ONE 2012, 7, e46781. [Google Scholar] [CrossRef]
- Kusminski, C.M.; Bickel, P.E.; Scherer, P.E. Targeting adipose tissue in the treatment of obesity-associated diabetes. Nat. Rev. Drug Discov. 2016, 15, 639–660. [Google Scholar] [CrossRef] [PubMed]
- Jahandideh, F.; Chakrabarti, S.; Davidge, S.T.; Wu, J. Egg white hydrolysate shows insulin mimetic and sensitizing effects in 3T3-F442A pre-adipocytes. PLoS ONE 2017, 12, e0185653. [Google Scholar] [CrossRef] [PubMed]
- Son, M.; Wu, J. Egg white hydrolysate and peptide reverse insulin resistance associated with tumor necrosis factor-alpha (TNF-alpha) stimulated mitogen-activated protein kinase (MAPK) pathway in skeletal muscle cells. Eur. J. Nutr. 2018. [Google Scholar] [CrossRef] [PubMed]
- Koushan, K.; Rusovici, R.; Li, W.; Ferguson, L.R.; Chalam, K.V. The role of lutein in eye-related disease. Nutrients 2013, 5, 1823–1839. [Google Scholar] [CrossRef]
- Fiedor, J.; Burda, K. Potential role of carotenoids as antioxidants in human health and disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef]
- Goodrow, E.F.; Wilson, T.A.; Houde, S.C.; Vishwanathan, R.; Scollin, P.A.; Handelman, G.; Nicolosi, R.J. Consumption of one egg per day increases serum lutein and zeaxanthin concentrations in older adults without altering serum lipid and lipoprotein cholesterol concentrations. J. Nutr. 2006, 136, 2519–2524. [Google Scholar] [CrossRef]
- Sommerburg, O.; Keunen, J.E.; Bird, A.C.; van Kuijk, F.J. Fruits and vegetables that are sources for lutein and zeaxanthin: The macular pigment in human eyes. Br. J. Ophthalmol. 1998, 82, 907–910. [Google Scholar] [CrossRef]
- Lima, V.C.; Rosen, R.B.; Maia, M.; Prata, T.S.; Dorairaj, S.; Farah, M.E.; Sallum, J. Macular pigment optical density measured by dual-wavelength autofluorescence imaging in diabetic and nondiabetic patients: A comparative study. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5840–5845. [Google Scholar] [CrossRef]
- Hu, B.J.; Hu, Y.N.; Lin, S.; Ma, W.J.; Li, X.R. Application of Lutein and Zeaxanthin in nonproliferative diabetic retinopathy. Int. J. Ophthalmol. 2011, 4, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Brazionis, L.; Rowley, K.; Itsiopoulos, C.; O’Dea, K. Plasma carotenoids and diabetic retinopathy. Br. J. Nutr. 2009, 101, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Gao, D.H.; Xiang, X.; Xiong, Y.F.; Zhu, T.S.; Liu, L.G.; Sun, X.F.; Hao, L.P. Ameliorative effects of lutein on non-alcoholic fatty liver disease in rats. World J. Gastroenterol. 2015, 21, 8061–8072. [Google Scholar] [CrossRef] [PubMed]
- Tuzcu, M.; Orhan, C.; Muz, O.E.; Sahin, N.; Juturu, V.; Sahin, K. Lutein and zeaxanthin isomers modulates lipid metabolism and the inflammatory state of retina in obesity-induced high-fat diet rodent model. BMC Ophthalmol. 2017, 17, 129. [Google Scholar] [CrossRef] [PubMed]
- Muriach, M.; Bosch-Morell, F.; Alexander, G.; Blomhoff, R.; Barcia, J.; Arnal, E.; Almansa, I.; Romero, F.J.; Miranda, M. Lutein effect on retina and hippocampus of diabetic mice. Free Radic. Biol. Med. 2006, 41, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Lenzen, S. The mechanisms of alloxan- and streptozotocin-induced diabetes. Diabetologia 2008, 51, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Kowluru, R.A.; Menon, B.; Gierhart, D.L. Beneficial effect of zeaxanthin on retinal metabolic abnormalities in diabetic rats. Investig. Ophthalmol. Vis. Sci. 2008, 49, 1645–1651. [Google Scholar] [CrossRef]
- Yu, H.; Wark, L.; Ji, H.; Willard, L.; Jaing, Y.; Han, J.; He, H.; Ortiz, E.; Zhang, Y.; Medeiros, D.M.; et al. Dietary wolfberry upregulates carotenoid metabolic genes and enhances mitochondrial biogenesis in the retina of db/db diabetic mice. Mol. Nutr. Food Res. 2013, 57, 1158–1169. [Google Scholar] [CrossRef]
- Tang, L.; Zhang, Y.; Jiang, Y.; Willard, L.; Ortiz, E.; Wark, L.; Medeiros, D.; Lin, D. Dietary wolfberry ameliorates retinal structure abnormalities in db/db mice at the early stage of diabetes. Exp. Biol. Med. 2011, 236, 1051–1063. [Google Scholar] [CrossRef]
- Arnal, E.; Miranda, M.; Johnsen-Soriano, S.; Alvarez-Nolting, R.; Diaz-Llopis, M.; Araiz, J.; Cervera, E.; Bosch-Morell, F.; Romero, F.J. Beneficial effect of docosahexanoic acid and lutein on retinal structural, metabolic, and functional abnormalities in diabetic rats. Curr. Eye Res. 2009, 34, 928–938. [Google Scholar] [CrossRef]
- Zeisel, S.H. Choline: Critical role during fetal development and dietary requirements in adults. Ann. Rev. Nutr. 2006, 26, 229–250. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.M.; Vance, D.E. The active synthesis of phosphatidylcholine is required for very low density lipoprotein secretion from rat hepatocytes. J. Biol. Chem. 1988, 263, 2998–3004. [Google Scholar] [PubMed]
- Sun, H.; Lee, E.J.; Samaraweera, H.; Persia, M.; Ahn, D.U. Effects of increasing concentrations of corn distillers dried grains with solubles on chemical composition and nutrient content of egg. Poult. Sci. 2013, 92, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Patterson, K.Y.; Bhagwat, S.A.; Williams, J.R.; Howe, J.C.; Holden, J.M. USDA Database for the Choline Content of Common Foods, Release 2. USDA, 2008. Available online: https://www.ars.usda.gov/ARSUserFiles/80400525/Data/Choline/Choln02.pdf (accessed on 2 February 2019).
- Fullerton, M.D.; Hakimuddin, F.; Bonen, A.; Bakovic, M. The development of a metabolic disease phenotype in CTP: Phosphoethanolamine cytidylyltransferase-deficient mice. J. Biol. Chem. 2009, 284, 25704–25713. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Wang, Y.; Sun, G. High dietary choline and betaine intake is associated with low insulin resistance in the Newfoundland population. Nutrition 2017, 33, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Ibsen, K.; Brown, T.; Chen, R.; Agatemor, C.; Mitragotri, S. Ionic liquids for oral insulin delivery. Proc. Natl. Acad. Sci. USA 2018, 115, 7296–7301. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, R.L.; Zhao, Y.; Koonen, D.P.; Sletten, T.; Su, B.; Lingrell, S.; Cao, G.; Peake, D.A.; Kuo, M.S.; Proctor, S.D.; et al. Impaired de novo choline synthesis explains why phosphatidylethanolamine N-methyltransferase-deficient mice are protected from diet-induced obesity. J. Biol. Chem. 2010, 285, 22403–22413. [Google Scholar] [CrossRef]
- Wu, G.; Zhang, L.; Li, T.; Lopaschuk, G.; Vance, D.E.; Jacobs, R.L. Choline Deficiency Attenuates Body Weight Gain and Improves Glucose Tolerance in ob/ob Mice. J. Obes. 2012, 2012, 319172. [Google Scholar] [CrossRef]
- Li, Y.; Wang, D.D.; Chiuve, S.E.; Manson, J.E.; Willett, W.C.; Hu, F.B.; Qi, L. Dietary phosphatidylcholine intake and type 2 diabetes in men and women. Diabetes Care 2015, 38, e13–e14. [Google Scholar] [CrossRef]
- Fujisawa, T.; Ikegami, H.; Inoue, K.; Kawabata, Y.; Ogihara, T. Effect of two alpha-glucosidase inhibitors, voglibose and acarbose, on postprandial hyperglycemia correlates with subjective abdominal symptoms. Metabolism 2005, 54, 387–390. [Google Scholar] [CrossRef]
- Krentz, A.J.; Bailey, C.J. Oral antidiabetic agents: Current role in type 2 diabetes mellitus. Drugs 2005, 65, 385–411. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.Y.; Choi, D.S.; Lee, M.K.; Lee, H.W.; Park, T.S.; Kim, D.M.; Chung, C.H.; Kim, D.K.; Kim, I.J.; Jang, H.C.; et al. Comparison of acarbose and voglibose in diabetes patients who are inadequately controlled with basal insulin treatment: Randomized, parallel, open-label, active-controlled study. J. Korean Med. Sci. 2014, 29, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Moritoh, Y.; Takeuchi, K.; Hazama, M. Chronic administration of voglibose, an alpha-glucosidase inhibitor, increases active glucagon-like peptide-1 levels by increasing its secretion and decreasing dipeptidyl peptidase-4 activity in ob/ob mice. J. Pharmacol. Exp. Ther. 2009, 329, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Goke, B.; Fuder, H.; Wieckhorst, G.; Theiss, U.; Stridde, E.; Littke, T.; Kleist, P.; Arnold, R.; Lucker, P.W. Voglibose (AO-128) is an efficient alpha-glucosidase inhibitor and mobilizes the endogenous GLP-1 reserve. Digestion 1995, 56, 493–501. [Google Scholar] [CrossRef]
- Hucking, K.; Kostic, Z.; Pox, C.; Ritzel, R.; Holst, J.J.; Schmiegel, W.; Nauck, M.A. alpha-Glucosidase inhibition (acarbose) fails to enhance secretion of glucagon-like peptide 1 (7-36 amide) and to delay gastric emptying in Type 2 diabetic patients. Diabet. Med. 2005, 22, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Yin, Y.; Zhao, W.; Liu, J.; Chen, F. Anti-diabetic activity peptides from albumin against alpha-glucosidase and alpha-amylase. Food Chem. 2012, 135, 2078–2085. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Yin, Y.; Zhao, W.; Yu, Y.; Liu, B.; Liu, J.; Chen, F. Novel peptides derived from egg white protein inhibiting alpha-glucosidase. Food Chem. 2011, 129, 1376–1382. [Google Scholar] [CrossRef]
- Zambrowicz, A.; Pokora, M.; Setner, B.; Dabrowska, A.; Szoltysik, M.; Babij, K.; Szewczuk, Z.; Trziszka, T.; Lubec, G.; Chrzanowska, J. Multifunctional peptides derived from an egg yolk protein hydrolysate: Isolation and characterization. Amino Acids 2015, 47, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2. [Google Scholar] [CrossRef] [PubMed]
- Muniyappa, R.; Yavuz, S. Metabolic actions of angiotensin II and insulin: A microvascular endothelial balancing act. Mol. Cell. Endocrinol. 2013, 378, 59–69. [Google Scholar] [CrossRef]
- Cordonnier, D.J.; Zaoui, P.; Halimi, S. Role of ACE inhibitors in patients with diabetes mellitus. Drugs 2001, 61, 1883–1892. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Wu, J. Milk-derived tripeptides IPP (Ile-Pro-Pro) and VPP (Val-Pro-Pro) promote adipocyte differentiation and inhibit inflammation in 3T3-F442A cells. PLoS ONE 2015, 10, e0117492. [Google Scholar] [CrossRef] [PubMed]
- Son, M.; Chan, C.B.; Wu, J. Egg White Ovotransferrin-Derived ACE Inhibitory Peptide Ameliorates Angiotensin II-Stimulated Insulin Resistance in Skeletal Muscle Cells. Mol. Nutr. Food Res. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Rohrborn, D.; Wronkowitz, N.; Eckel, J. DPP4 in Diabetes. Front. Immunol. 2015, 6, 386. [Google Scholar] [CrossRef] [PubMed]
- Uenishi, H.; Kabuki, T.; Seto, Y.; Serizawa, A.; Nakajima, H. Isolation and identification of casein-derived dipeptidyl-peptidase 4 (DPP-4)-inhibitory peptide LPQNIPPL from gouda-type cheese and its effect on plasma glucose in rats. Int. Dairy J. 2012, 22, 24–30. [Google Scholar] [CrossRef]
- Huang, S.L.; Hung, C.C.; Jao, C.L.; Tung, Y.S.; Hsu, K.C. Porcine skin gelatin hydrolysate as a dipeptidyl peptidase IV inhibitor improves glycemic control in streptozotocin-induced diabetic rats. J. Funct. Foods 2014, 11, 235–242. [Google Scholar] [CrossRef]
Figure 1.
Major bioactivities of egg components and egg-derived peptides that possibly contribute to the benefits in T2D.
Figure 1.
Major bioactivities of egg components and egg-derived peptides that possibly contribute to the benefits in T2D.

{kind=link}
Table 1.
Recent clinical studies investigating the effect of egg consumption on T2D
| Egg Dose | Subjects | Duration | Study Design | Primary and Secondary Outcomes | Results |
|---|---|---|---|---|---|
| 3 eggs/day [52] | 37 women with metabolic syndrome *; aged 30–70 years | 12 weeks | Randomized, single blind, parallel design | Plasma lipids, apolipoprotein, oxLDL, CETP and LCAT | Improved HDL, large HDL particles, total and medium VLDL particles, HOMA-IR, and LCAT activity |
| 2 eggs/day [53] | 65 subjects with T2D or impaired glucose tolerance; aged 54 ± 8.2; BMI 34.1 ± 4.8 kg/m2 | 12 weeks | Randomized, controlled, parallel design | Blood lipid, glucose, insulin, HbA1c, CRP and apoprotein-B, homocystein | Increased HDL cholesterol; improved glycemic and lipid profiles |
| Egg breakfast [57] | 21 healthy men; aged 20–70 years | 1 week | Randomized, Cross-over | Fasting blood glucose, plasma insulin, ghrelin, leptin, GLP-1, PYY | Less variation in plasma glucose and insulin; reduced ghrelin response and energy intake |
| 2 eggs per day [58] | 34 adults with T2D (14 postmenopausal women and 20 men); mean age = 64.5 years | 12 weeks | Randomized, controlled, single-blind, cross-over | Glycated hemoglobin, systolic blood pressure, body mass index, visceral fat rating, waist circumference, and percent body fat | Reduced body mass index, visceral fat, waist circumference and percent body fat; unchanged glycemic control |
| High-egg diet (≥12 eggs/week) or low-egg diet (<2 eggs/week) [54] | 140 aged subjects with prediabetes or T2D; BMI ≥ 25 kg/m2 | 3 months | Randomized, controlled, parallel-arm | Plasma blood glucose, HbA1c, TC, LDL-C, HDL-C, TG, apolipoprotein B, CRP | No significant changes between groups |
| High-egg diet (≥12 eggs/week) or low-egg diet (<2 eggs/week) [59] | 128 subjects with prediabetes or T2D; aged ≥18 years; BMI ≥25 kg/m2 | 12 months | Randomized, controlled, parallel-arm | Plasma glucose, HbA1c, 1,5-anhydroglucitol, traditional serum lipids, markers of inflammation, high-sensitivity C-reactive protein, interleukin 6, soluble E-selectin, oxidative stress, and adiponectin | No significant changes between groups |
* The National cholesterol Education Program’s Adult Treatment Panel III report definition [60]. cholesteryl ester transfer protein: CETP; C-reactive protein: CRP; glucagon-like peptide 1: GLP-1; lecithin cholesterol acyltransferase: LCAT; total cholesterol: TC; triglyceride: TG; oxidized LDL: oxLDL.
Table 2.
In vivo studies of the effect of EWH on T2D.
| EWH Preparation Method | Animal Model/Group | EWH Dose/ Duration | Major Results |
|---|---|---|---|
| Protease [90] | Nagoya-Shibata-Yasuda mice | 27.6% (w/w, diet) /8 weeks | Decreased plasma glucose and insulin concentration; improved insulin resistance |
| Alcalase [94] | Obese Zucker rats | 1 g/kg (body weight)/day/15 weeks | Reduced renal mRNA expression of IL-1β, IL-13, and TNF-α; decreased renal P22(phox)protein expression; unchanged blood GLP-1 and glucose concentration |
| Thermolysin and Pepsin [91] | High-fat diet-fed rats | 4% (w/w, diet)/6 weeks | Reduced plasma IL-1α, IL-β, and MCP-1 concentrations and fat pad mass; increased lean mass and upregulated Akt phosphorylation in liver, muscle, and fat tissues; improved glucose tolerance and insulin sensitivity |
| Protease [92] | Goto-Kakizaki rats | 27.6% (w/w, diet)/6 weeks | Decreased fasting blood glucose concentration and triglyceride content in muscle; improved HOMA-IR; |
| Protease [93] | Rats fed with a high-fat and high-sucrose diet | 39.4% (w/w, diet)/6 weeks | Reduced food intake, body weight gain and fat deposition; decreased stearoyl-CoA desaturase and glucose-6-phosphate dehydrogenase activity in liver and muscle; suppressed serum levels of triacylglycerol and leptin; increased muscle weight; upregulated fecal excretion of triacylglycerol, cholesterol, and total bile acids |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, X.; Son, M.; Meram, C.; Wu, J. Mechanism and Potential of Egg Consumption and Egg Bioactive Components on Type-2 Diabetes. Nutrients 2019, 11, 357. https://doi.org/10.3390/nu11020357
AMA Style
Wang X, Son M, Meram C, Wu J. Mechanism and Potential of Egg Consumption and Egg Bioactive Components on Type-2 Diabetes. Nutrients. 2019; 11(2):357. https://doi.org/10.3390/nu11020357
Chicago/Turabian StyleWang, Xiaofeng, Myoungjin Son, Chalamaiah Meram, and Jianping Wu. 2019. "Mechanism and Potential of Egg Consumption and Egg Bioactive Components on Type-2 Diabetes" Nutrients 11, no. 2: 357. https://doi.org/10.3390/nu11020357
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.