Sugar Responses of Human Enterochromaffin Cells Depend on Gut Region, Sex, and Body Mass

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Cell Isolation
2.3. Secretion Assay, 5-HT Content and Sensitivity Measurements
2.4. Statistics
3. Results
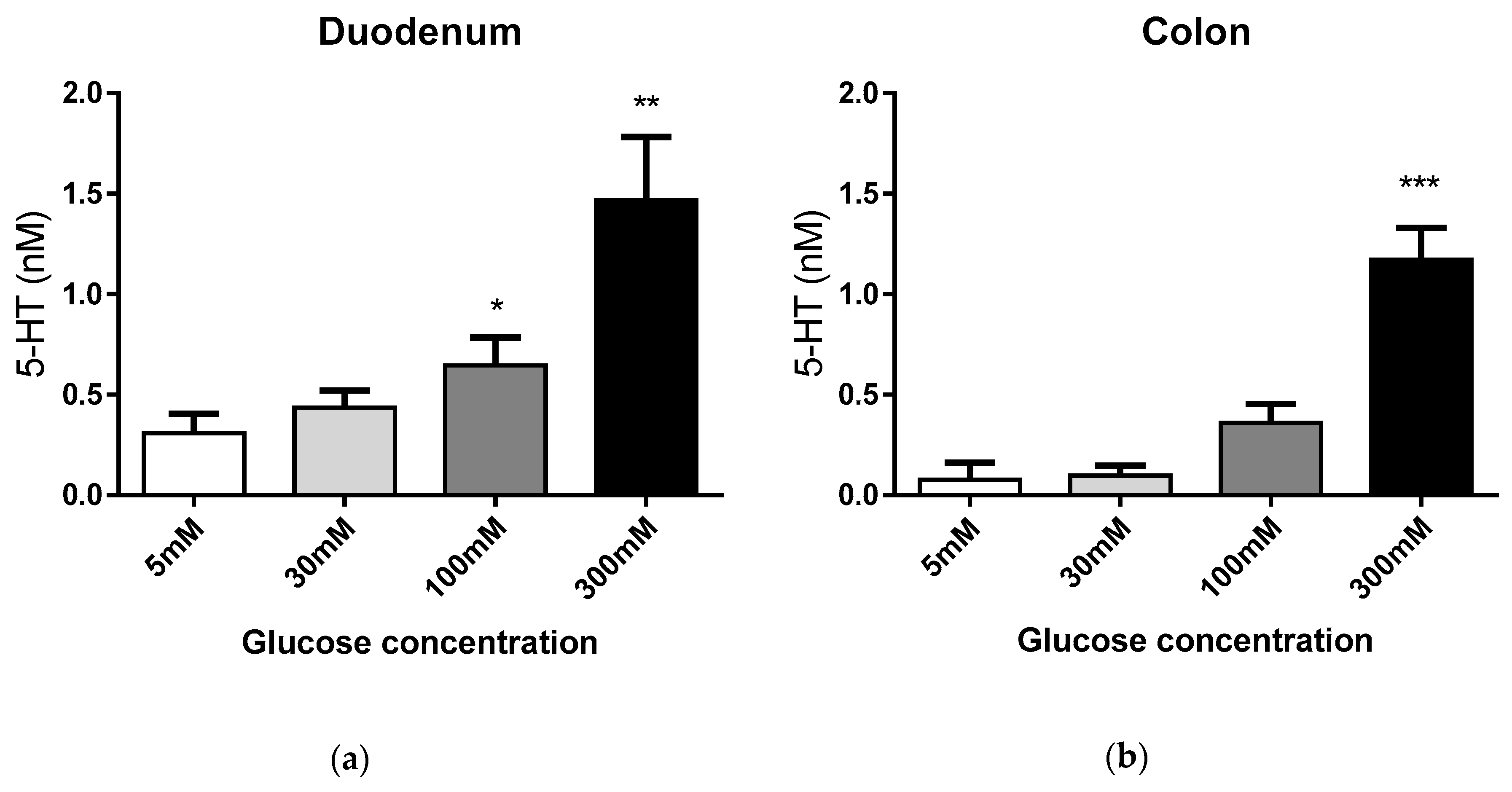
3.1. EC Cell Responses to Extracellular Glucose
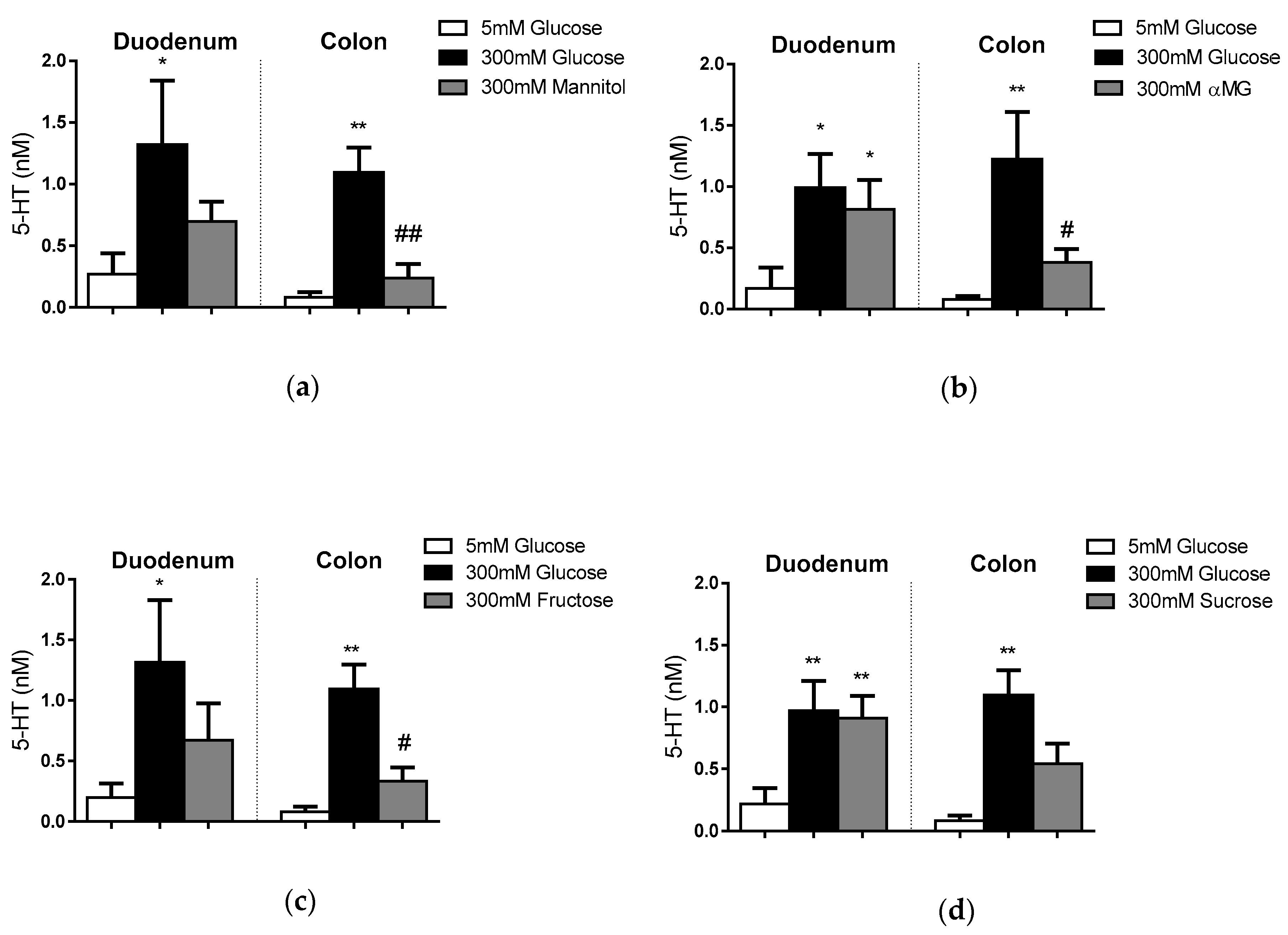
3.2. EC Cell Responses to Other Extracellular Sugars
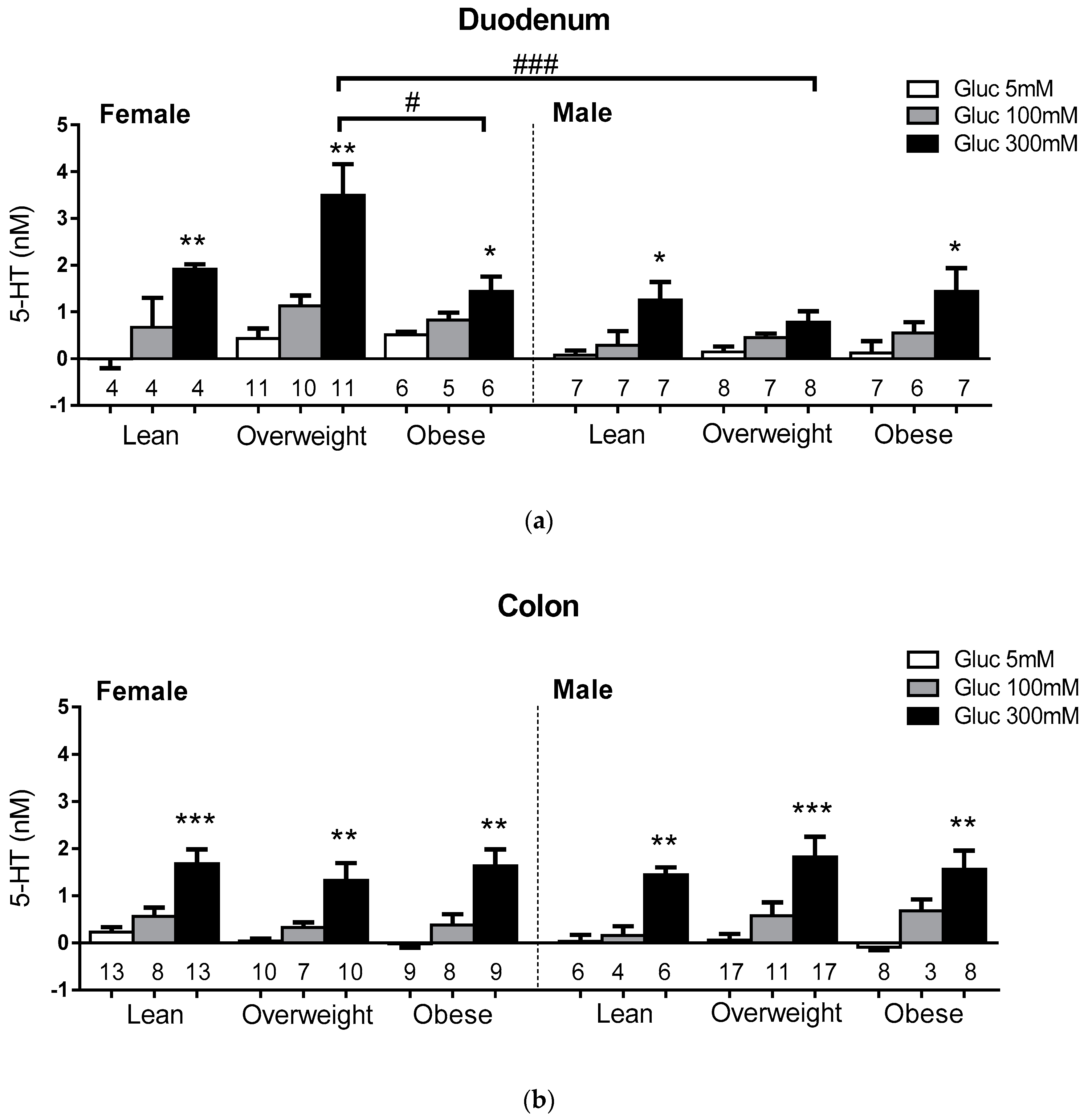
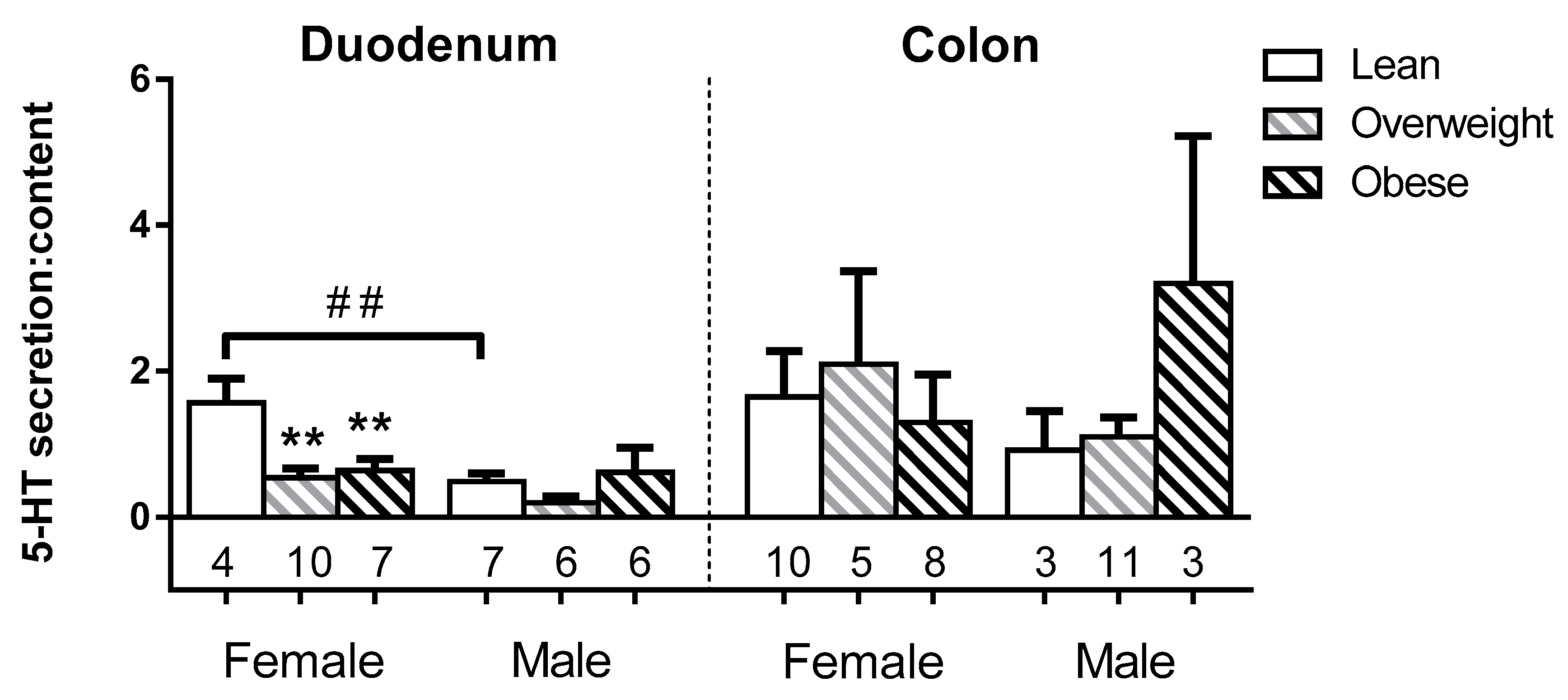
3.3. Sex and BMI Related Effects on EC 5-HT Content
3.4. Sex and BMI Related Effects on Glucose-Evoked 5-HT Release.
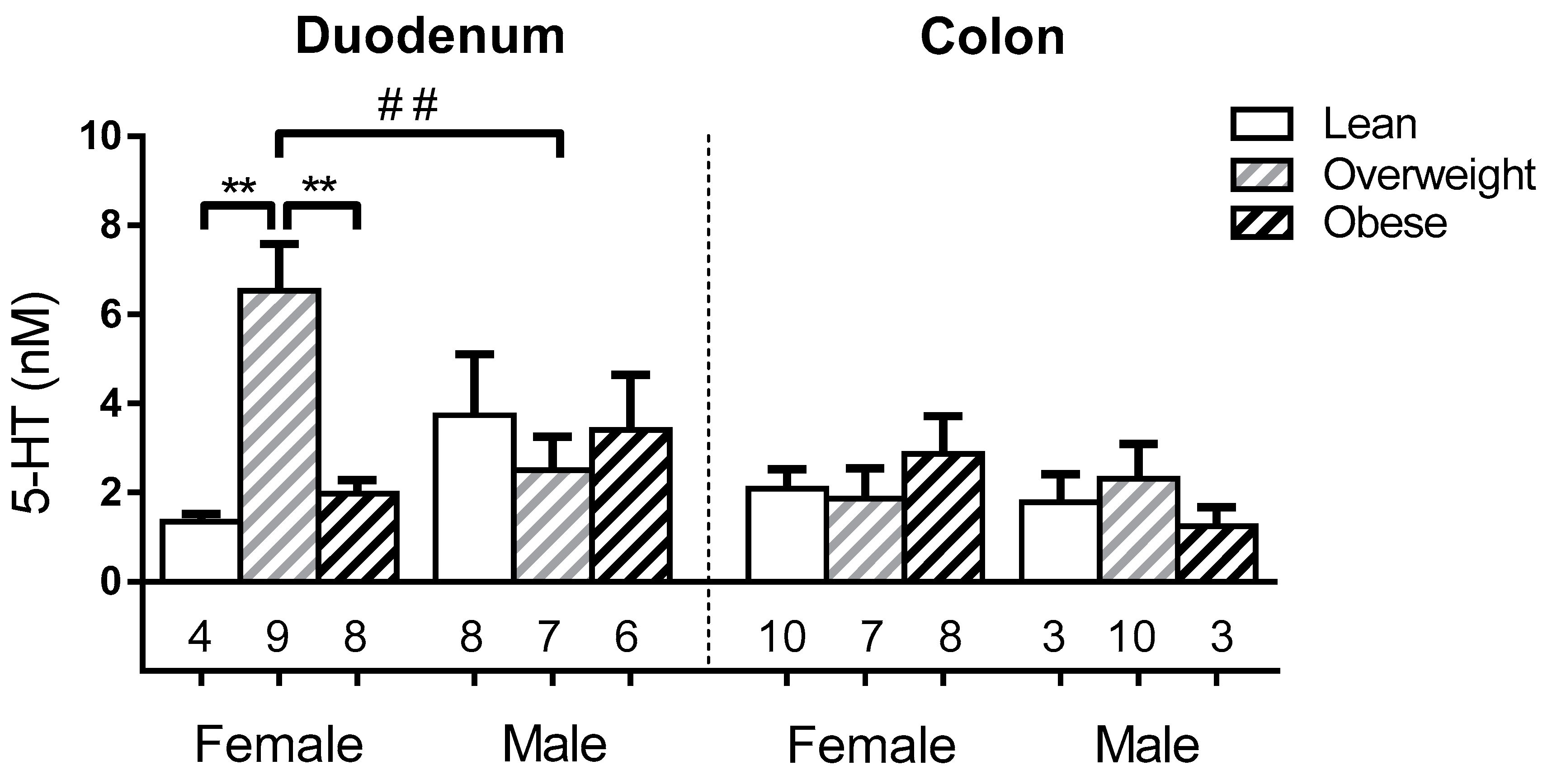
3.5. Sex and BMI Related Effects on EC Cell Glucose Sensitivity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Walther, D.J.; Bader, M. A unique central tryptophan hydroxylase isoform. Biochem. Pharmacol. 2003, 66, 1673–1680. [Google Scholar] [CrossRef]
- Côté, F.; Thévenot, E.; Fligny, C.; Fromes, Y.; Darmon, M.; Ripoche, M.A.; Bayard, E.; Hanoun, N.; Saurini, F.; Lechat, P.; et al. Disruption of the nonneuronal tph1 gene demonstrates the importance of peripheral serotonin in cardiac function. Proc. Natl. Acad. Sci. USA 2003, 100, 13525–13530. [Google Scholar] [CrossRef] [Green Version]
- Erspamer, V. Pharmacology of indole-alkylamines. Pharmacol. Rev. 1954, 6, 425–487. [Google Scholar]
- Kidd, M.; Modlin, I.M.; Gustafsson, B.I.; Drozdov, I.; Hauso, O.; Pfragner, R. Luminal regulation of normal and neoplastic human EC cell serotonin release is mediated by bile salts, amines, tastants, and olfactants. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G260–G272. [Google Scholar] [CrossRef]
- Symonds, E.L.; Peiris, M.; Page, A.J.; Chia, B.; Dogra, H.; Masding, A.; Galanakis, V.; Atiba, M.; Bulmer, D.; Young, R.L.; et al. Mechanisms of activation of mouse and human enteroendocrine cells by nutrients. Gut 2015, 64, 618–626. [Google Scholar] [CrossRef]
- Martin, A.M.; Lumsden, A.L.; Young, R.L.; Jessup, C.F.; Spencer, N.J.; Keating, D.J. The nutrient-sensing repertoires of mouse enterochromaffin cells differ between duodenum and colon. Neurogastroenterol. Motil. 2017. [Google Scholar] [CrossRef]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef]
- Spencer, N.J.; Nicholas, S.J.; Robinson, L.; Kyloh, M.; Flack, N.; Brookes, S.J.; Zagorodnyuk, V.P.; Keating, D.J. Mechanisms underlying distension-evoked peristalsis in guinea pig distal colon: Is there a role for enterochromaffin cells? Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G519–G527. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Mizumoto, A.; Mochiki, E.; Haga, N.; Suzuki, H.; Itoh, Z. Relationship between intraduodenal 5-hydroxytryptamine release and interdigestive contractions in dogs. J. Smooth Muscle Res. 2004, 40, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Zelkas, L.; Raghupathi, R.; Lumsden, A.L.; Martin, A.M.; Sun, E.; Spencer, N.J.; Young, R.L.; Keating, D.J. Serotonin-secreting enteroendocrine cells respond via diverse mechanisms to acute and chronic changes in glucose availability. Nutr. Metab. (Lond.) 2015, 12, 55. [Google Scholar] [CrossRef]
- Martin, A.M.; Lumsden, A.L.; Young, R.L.; Jessup, C.F.; Spencer, N.J.; Keating, D.J. Regional differences in nutrient-induced secretion of gut serotonin. Physiol. Rep. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.X.; Zhu, X.Y.; Owyang, C.; Li, Y. Intestinal serotonin acts as a paracrine substance to mediate vagal signal transmission evoked by luminal factors in the rat. J. Physiol. 2001, 530 Pt 3, 431–442. [Google Scholar] [CrossRef] [Green Version]
- Keating, D.J.; Spencer, N.J. Release of 5-hydroxytryptamine from the mucosa is not required for the generation or propagation of colonic migrating motor complexes. Gastroenterology 2010, 138, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Spencer, N.J.; Sia, T.C.; Brookes, S.J.; Costa, M.; Keating, D.J. CrossTalk opposing view: 5-HT is not necessary for peristalsis. J. Physiol. 2015, 593, 3229–3231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertrand, P.P.; Kunze, W.A.; Furness, J.B.; Bornstein, J.C. The terminals of myenteric intrinsic primary afferent neurons of the guinea-pig ileum are excited by 5-hydroxytryptamine acting at 5-hydroxytryptamine-3 receptors. Neuroscience 2000, 101, 459–469. [Google Scholar] [CrossRef]
- Spencer, N.J.; Keating, D.J. Is there a role for endogenous 5-ht in gastrointestinal motility? How recent studies have changed our understanding. Adv. Exp. Med. Biol. 2016, 891, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B.; Rivera, L.R.; Cho, H.J.; Bravo, D.M.; Callaghan, B. The gut as a sensory organ. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.B.; Witte, A.B. The role of serotonin in intestinal luminal sensing and secretion. Acta Physiol. (Oxf.) 2008, 193, 311–323. [Google Scholar] [CrossRef]
- Rogers, G.B.; Keating, D.J.; Young, R.L.; Wong, M.L.; Licinio, J.; Wesselingh, S. From gut dysbiosis to altered brain function and mental illness: Mechanisms and pathways. Mol. Psychiatry 2016, 21, 738–748. [Google Scholar] [CrossRef]
- Sumara, G.; Sumara, O.; Kim, J.K.; Karsenty, G. Gut-derived serotonin is a multifunctional determinant to fasting adaptation. Cell Metab. 2012, 16, 588–600. [Google Scholar] [CrossRef]
- Bertrand, R.L.; Senadheera, S.; Markus, I.; Liu, L.; Howitt, L.; Chen, H.; Murphy, T.V.; Sandow, S.L.; Bertrand, P.P. A Western diet increases serotonin availability in rat small intestine. Endocrinology 2011, 152, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, J.H.; Noh, S.; Hur, H.J.; Sung, M.J.; Hwang, J.T.; Park, J.H.; Yang, H.J.; Kim, M.S.; Kwon, D.Y.; et al. Metabolomic analysis of livers and serum from high-fat diet induced obese mice. J. Proteome. Res. 2011, 10, 722–731. [Google Scholar] [CrossRef] [PubMed]
- Le Beyec, J.; Pelletier, A.L.; Arapis, K.; Hourseau, M.; Cluzeaud, F.; Descatoire, V.; Ducroc, R.; Aparicio, T.; Joly, F.; Couvelard, A.; et al. Overexpression of gastric leptin precedes adipocyte leptin during high-fat diet and is linked to 5HT-containing enterochromaffin cells. Int. J. Obes. (Lond.) 2014, 38, 1357–1364. [Google Scholar] [CrossRef] [PubMed]
- Young, R.L.; Lumsden, A.L.; Martin, A.M.; Schober, G.; Pezos, N.; Thazhath, S.S.; Isaacs, N.J.; Cvijanovic, N.; Sun, E.W.L.; Wu, T.; et al. Augmented capacity for peripheral serotonin release in human obesity. Int. J. Obes. 2018. [Google Scholar] [CrossRef] [PubMed]
- Crane, J.D.; Palanivel, R.; Mottillo, E.P.; Bujak, A.L.; Wang, H.; Ford, R.J.; Collins, A.; Blümer, R.M.; Fullerton, M.D.; Yabut, J.M.; et al. Inhibiting peripheral serotonin synthesis reduces obesity and metabolic dysfunction by promoting brown adipose tissue thermogenesis. Nat. Med. 2014. [Google Scholar] [CrossRef]
- Raghupathi, R.; Duffield, M.D.; Zelkas, L.; Meedeniya, A.; Brookes, S.J.; Sia, T.C.; Wattchow, D.A.; Spencer, N.J.; Keating, D.J. Identification of unique release kinetics of serotonin from guinea-pig and human enterochromaffin cells. J. Physiol. 2013, 591, 5959–5975. [Google Scholar] [CrossRef] [Green Version]
- Balen, D.; Ljubojevic, M.; Breljak, D.; Brzica, H.; Zlender, V.; Koepsell, H.; Sabolic, I. Revised immunolocalization of the Na+-D-glucose cotransporter SGLT1 in rat organs with an improved antibody. Am. J. Physiol. Cell Physiol. 2008, 295, C475–C489. [Google Scholar] [CrossRef]
- Sotak, M.; Marks, J.; Unwin, R.J. Putative tissue location and function of the SLC5 family member SGLT3. Exp. Physiol. 2017, 102, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Cooke, H.J.; Javed, N.H.; Carey, H.V.; Christofi, F.; Raybould, H.E. D-glucose releases 5-hydroxytryptamine from human BON cells as a model of enterochromaffin cells. Gastroenterology 2001, 121, 1400–1406. [Google Scholar] [CrossRef]
- Sun, E.W.; de Fontgalland, D.; Rabbitt, P.; Hollington, P.; Sposato, L.; Due, S.L.; Wattchow, D.A.; Rayner, C.K.; Deane, A.M.; Young, R.L.; et al. Mechanisms controlling glucose-induced Glp-1 Secretion in human small intestine. Diabetes 2017. [Google Scholar] [CrossRef]
- Freeman, S.L.; Bohan, D.; Darcel, N.; Raybould, H.E. Luminal glucose sensing in the rat intestine has characteristics of a sodium-glucose cotransporter. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 291, G439–G445. [Google Scholar] [CrossRef] [PubMed]
- Vincent, K.M.; Sharp, J.W.; Raybould, H.E. Intestinal glucose-induced calcium-calmodulin kinase signaling in the gut-brain axis in awake rats. Neurogastroenterol. Motil. 2011, 23, e282–e293. [Google Scholar] [CrossRef] [PubMed]
- Modlin, I.M.; Kidd, M.; Pfragner, R.; Eick, G.N.; Champaneria, M.C. The functional characterization of normal and neoplastic human enterochromaffin cells. J. Clin. Endocrinol. Metab. 2006, 91. [Google Scholar] [CrossRef] [PubMed]
- Afarideh, M.; Behdadnia, A.; Noshad, S.; Mirmiranpour, H.; Mousavizadeh, M.; Khajeh, E.; Rad, M.V.; Mazaheri, T.; Nakhjavani, M.; Esteghamati, A. Association of peripheral 5-hydroxyindole-3-acetic acid, a serotonin derivative, with metabolic syndrome and low-grade inflammation. Endocr. Pract. 2015, 21, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Toyofuku, Y.; Lynn, F.C.; Chak, E.; Uchida, T.; Mizukami, H.; Fujitani, Y.; Kawamori, R.; Miyatsuka, T.; Kosaka, Y.; et al. Serotonin regulates pancreatic beta cell mass during pregnancy. Nat. Med. 2010, 16, 804–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Oh, C.M.; Ohara-Imaizumi, M.; Park, S.; Namkung, J.; Yadav, V.K.; Tamarina, N.A.; Roe, M.W.; Philipson, L.H.; Karsenty, G.; et al. Functional role of serotonin in insulin secretion in a diet-induced insulin-resistant state. Endocrinology 2015, 156, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Young, R.L.; Lumsden, A.L.; Keating, D.J. Gut Serotonin Is a Regulator of Obesity and Metabolism. Gastroenterology 2015, 149, 253–255. [Google Scholar] [CrossRef]
- Raghupathi, R.; Jessup, C.F.; Lumsden, A.L.; Keating, D.J. Fusion pore size limits 5-HT release from single enterochromaffin cell vesicles. J. Cell Physiol. 2016, 231, 1593–1600. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, K.D.; Duffield, M.D.; Peiris, H.; Phillips, L.; Zanin, M.P.; Teo, E.H.; Zhou, X.F.; Keating, D.J. Huntingtin-associated protein 1 regulates exocytosis, vesicle docking, readily releasable pool size and fusion pore stability in mouse chromaffin cells. J. Physiol. 2014, 592, 1505–1518. [Google Scholar] [CrossRef] [Green Version]
- Jackson, J.; Papadopulos, A.; Meunie, F.A.; McCluskey, A.; Robinson, P.J.; Keating, D.J. Small molecules demonstrate the role of dynamin as a bi-directional regulator of the exocytosis fusion pore and vesicle release. Mol. Psychiatry 2015, 20, 810–819. [Google Scholar] [CrossRef]
- Thorn, P.; Zorec, R.; Rettig, J.; Keating, D.J. Exocytosis in non-neuronal cells. J. Neurochem. 2016, 137, 849–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkman, H.P.; Yates, K.; Hasler, W.L.; Nguyen, L.; Pasricha, P.J.; Snape, W.J.; Farrugia, G.; Koch, K.L.; Abell, T.L.; McCallum, R.W.; et al. Clinical features of idiopathic gastroparesis vary with sex, body mass, symptom onset, delay in gastric emptying, and gastroparesis severity. Gastroenterology 2011, 140, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Troy, A.E.; Simmonds, S.S.; Stocker, S.D.; Browning, K.N. High fat diet attenuates glucose-dependent facilitation of 5-HT3 -mediated responses in rat gastric vagal afferents. J. Physiol. 2016, 594, 99–114. [Google Scholar] [CrossRef] [PubMed]
- France, M.; Skorich, E.; Kadrofske, M.; Swain, G.M.; Galligan, J.J. Sex-related differences in small intestinal transit and serotonin dynamics in high-fat-diet-induced obesity in mice. Exp. Physiol. 2016, 101, 81–99. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Duodenum | Colon | |
|---|---|---|
| n | 15 | 17 |
| BMI (kg/m2) | 24.9 ± 0.9 | 25.0 ± 0.9 |
| Age (years) | 50.2 ± 5.4 | 69.6 ± 4.4 |
| M:F | 10:5 | 6:11 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lumsden, A.L.; Martin, A.M.; Sun, E.W.; Schober, G.; Isaacs, N.J.; Pezos, N.; Wattchow, D.A.; de Fontgalland, D.; Rabbitt, P.; Hollington, P.; et al. Sugar Responses of Human Enterochromaffin Cells Depend on Gut Region, Sex, and Body Mass. Nutrients 2019, 11, 234. https://doi.org/10.3390/nu11020234
Lumsden AL, Martin AM, Sun EW, Schober G, Isaacs NJ, Pezos N, Wattchow DA, de Fontgalland D, Rabbitt P, Hollington P, et al. Sugar Responses of Human Enterochromaffin Cells Depend on Gut Region, Sex, and Body Mass. Nutrients. 2019; 11(2):234. https://doi.org/10.3390/nu11020234
Chicago/Turabian StyleLumsden, Amanda L., Alyce M. Martin, Emily W. Sun, Gudrun Schober, Nicole J. Isaacs, Nektaria Pezos, David A. Wattchow, Dayan de Fontgalland, Philippa Rabbitt, Paul Hollington, and et al. 2019. "Sugar Responses of Human Enterochromaffin Cells Depend on Gut Region, Sex, and Body Mass" Nutrients 11, no. 2: 234. https://doi.org/10.3390/nu11020234