Evaluation of γ-oryzanol Accumulation and Lipid Metabolism in the Body of Mice Following Long-Term Administration of γ-oryzanol

Abstract

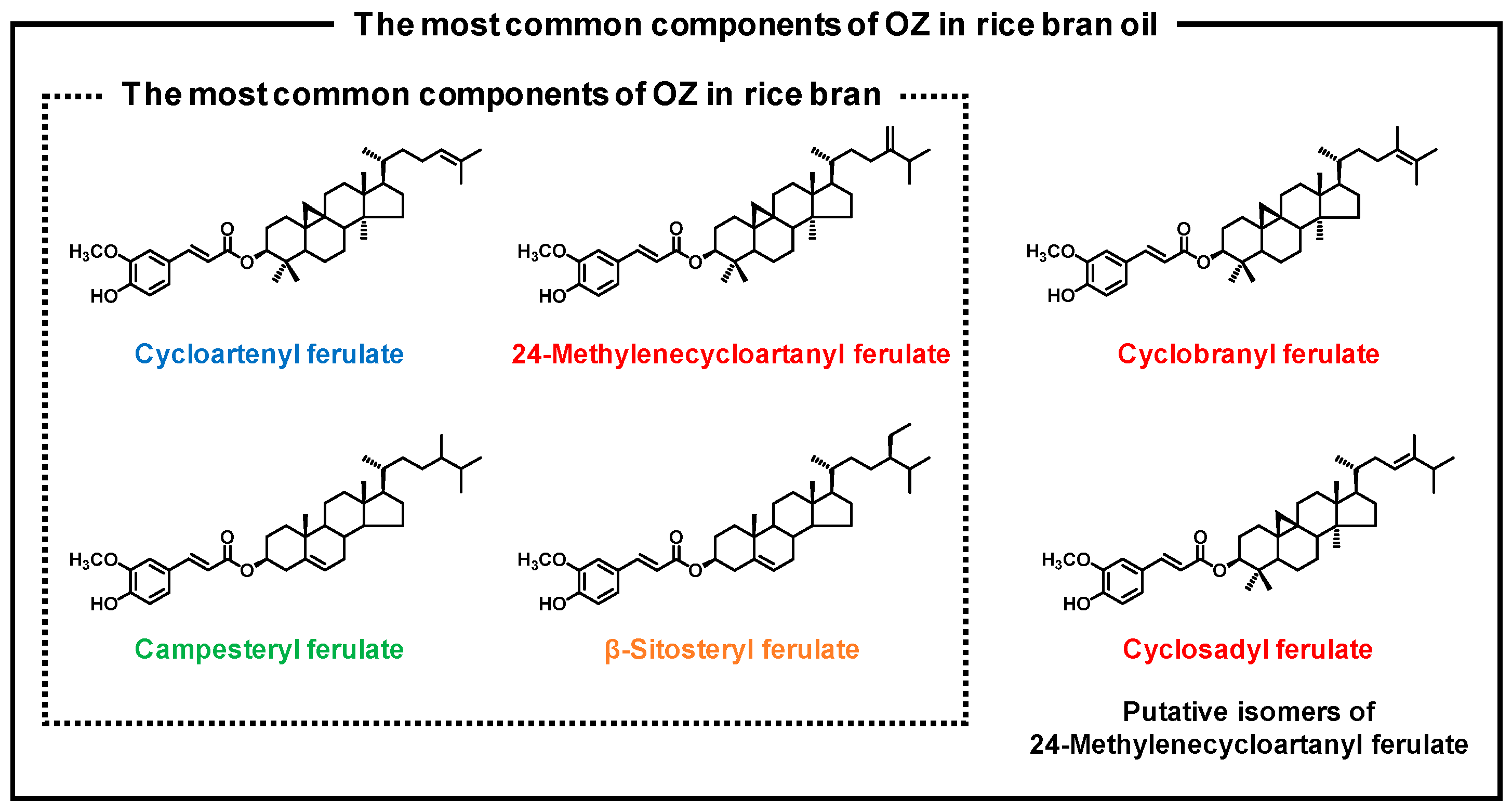
1. Introduction
2. Materials and Methods
2.1. Materials and Diets
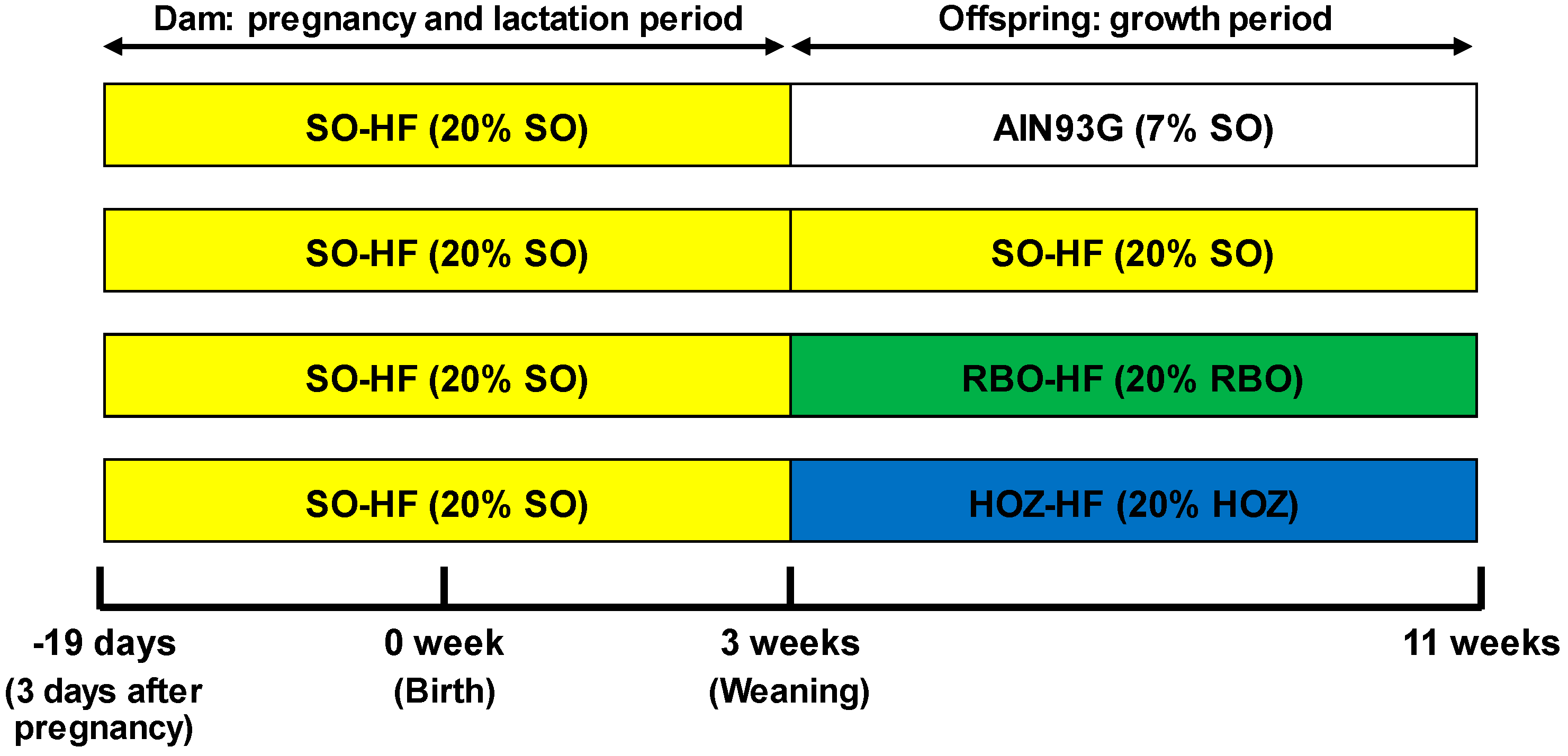
2.2. Animal Experiment of Obesity Model (Fetal Programming Model) Mice
2.3. Extraction of OZ from Plasma and Organs
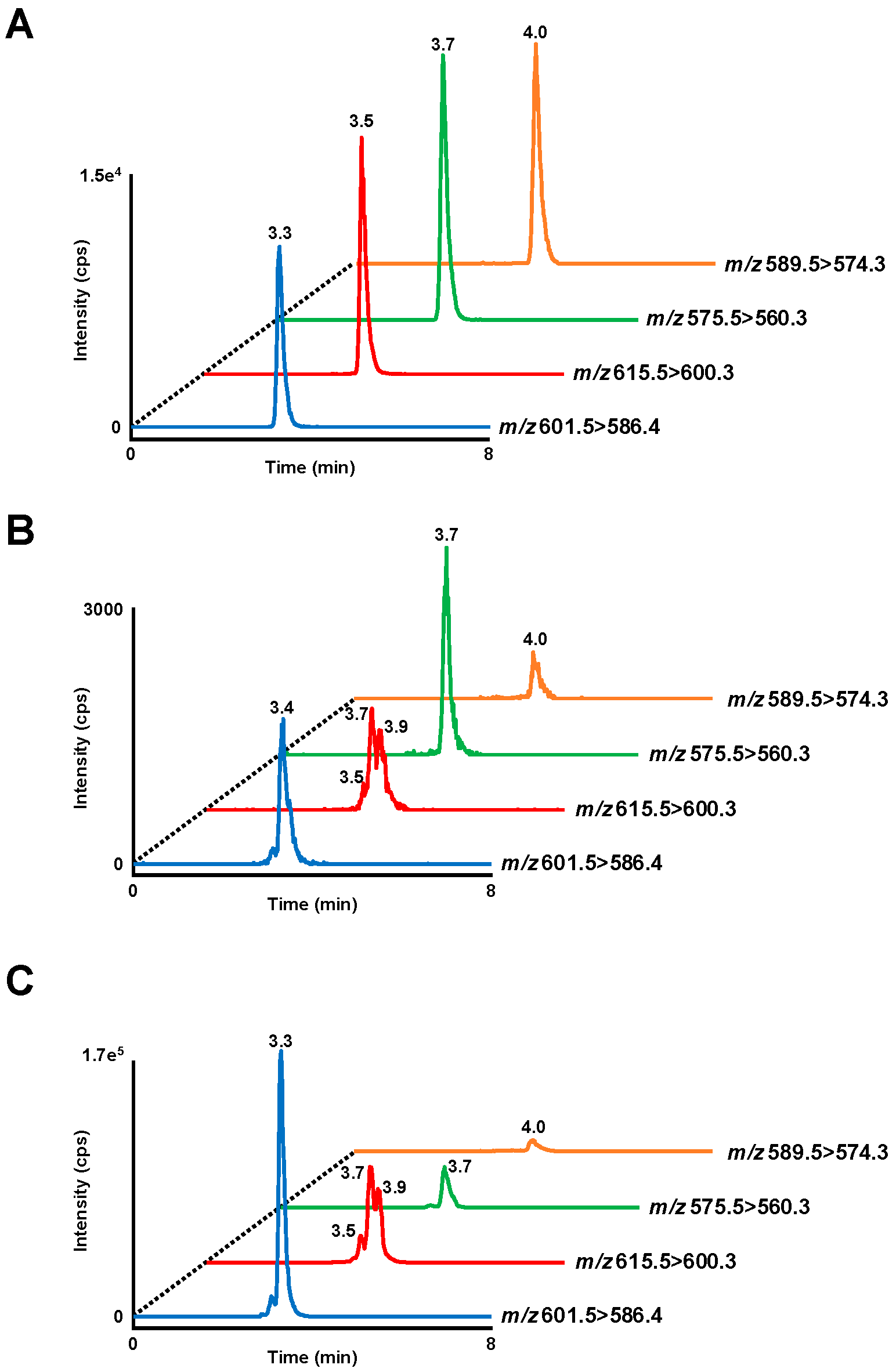
2.4. HPLC-MS/MS analysis of OZ
2.5. Biochemical Parameters
2.6. Single Oral Administration of OZ to Rats
2.7. Statistics
3. Results and Discussion
3.1. Body and Organ Weight of Fetal Programming Model Mice
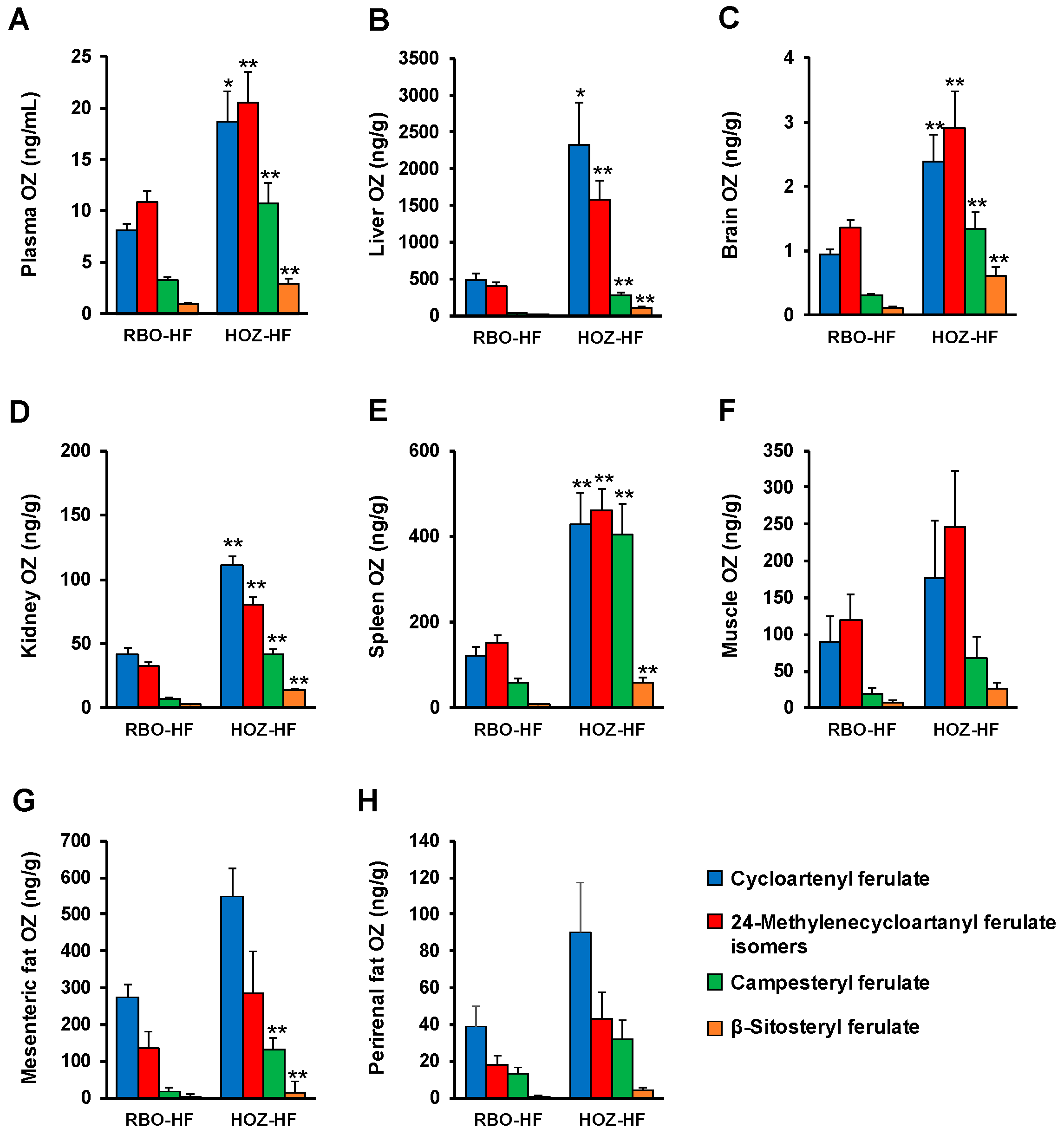
3.2. OZ Concentration in Plasma and Organs
3.3. Plasma and Liver Parameters of the Mice
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Clifford, M.N.; van der Hooft, J.J.; Crozier, A. Human studies on the absorption, distribution, metabolism, and excretion of tea polyphenols. Am. J. Clin. Nutr. 2013, 98, 1619s–1630s. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K.; Kiko, T.; Miyazawa, T.; Carpentero Burdeos, G.; Kimura, F.; Satoh, A.; Miyazawa, T. Antioxidant effect of astaxanthin on phospholipid peroxidation in human erythrocytes. Br. J. Nutr. 2011, 105, 1563–1571. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K.; Ninomiya, M.; Okubo, T.; Aoi, N.; Juneja, L.R.; Kim, M.; Yamanaka, K.; Miyazawa, T. Tea catechin supplementation increases antioxidant capacity and prevents phospholipid hydroperoxidation in plasma of humans. J. Agric. Food Chem. 1999, 47, 3967–3973. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, E.; Ito, J.; Kato, S.; Sawada, K.; Matsuki, M.; Hashimoto, H.; Miyazawa, T.; Nakagawa, K. Presence of orally administered rice bran oil gamma-oryzanol in its intact form in mouse plasma. Food Funct. 2016, 7, 4816–4822. [Google Scholar] [CrossRef] [PubMed]
- Ghatak, S.B.; Panchal, S.J. Investigation of the immunomodulatory potential of oryzanol isolated from crude rice bran oil in experimental animal models. Phytother. Res. PTR 2012, 26, 1701–1708. [Google Scholar] [CrossRef] [PubMed]
- Ismail, M.; Al-Naqeeb, G.; Mamat, W.A.; Ahmad, Z. Gamma-oryzanol rich fraction regulates the expression of antioxidant and oxidative stress related genes in stressed rat’s liver. Nutr. Metab. 2010, 7, 23. [Google Scholar] [CrossRef] [PubMed]
- Ismail, N.; Ismail, M.; Imam, M.U.; Azmi, N.H.; Fathy, S.F.; Foo, J.B.; Abu Bakar, M.F. Mechanistic basis for protection of differentiated SH-SY5Y cells by oryzanol-rich fraction against hydrogen peroxide-induced neurotoxicity. BMC Complement. Altern. Med. 2014, 14, 467. [Google Scholar] [CrossRef]
- Kim, S.P.; Kang, M.Y.; Nam, S.H.; Friedman, M. Dietary rice bran component gamma-oryzanol inhibits tumor growth in tumor-bearing mice. Mol. Nutr. Food Res. 2012, 56, 935–944. [Google Scholar] [CrossRef]
- Wang, O.; Liu, J.; Cheng, Q.; Guo, X.; Wang, Y.; Zhao, L.; Zhou, F.; Ji, B. Effects of ferulic acid and gamma-oryzanol on high-fat and high-fructose diet-induced metabolic syndrome in rats. PLoS ONE 2015, 10, e0118135. [Google Scholar] [CrossRef]
- Wilson, T.A.; Nicolosi, R.J.; Woolfrey, B.; Kritchevsky, D. Rice bran oil and oryzanol reduce plasma lipid and lipoprotein cholesterol concentrations and aortic cholesterol ester accumulation to a greater extent than ferulic acid in hypercholesterolemic hamsters. J. Nutr. Biochem. 2007, 18, 105–112. [Google Scholar] [CrossRef]
- Taniguchi, H.; Hashimoto, H.; Hosoda, A.; Kometani, T.; Tsuno, T.; Adachi, S. Functionality of compounds contained in rice bran and their improvement. Nippon Shokuhin Kagaku Kogaku Kaishi 2012, 59, 301–318. [Google Scholar] [CrossRef]
- Fujiwara, S.; Sakurai, S.; Noumi, K.; Sugimoto, I.; Awata, N. Metabolism of gamma-oryzanol in rabbit (author’s transl). Yakugaku zasshi J. Pharm. Soc. Jpn. 1980, 100, 1011–1018. [Google Scholar] [CrossRef]
- Ito, J.; Nakagawa, K.; Kato, S.; Miyazawa, T.; Kimura, F.; Miyazawa, T. The combination of maternal and offspring high-fat diets causes marked oxidative stress and development of metabolic syndrome in mouse offspring. Life Sci. 2016, 151, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Odaka, Y.; Nakano, M.; Tanaka, T.; Kaburagi, T.; Yoshino, H.; Sato-Mito, N.; Sato, K. The influence of a high-fat dietary environment in the fetal period on postnatal metabolic and immune function. Obesity 2010, 18, 1688–1694. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Cudd, T.A.; Meininger, C.J.; Spencer, T.E. Maternal nutrition and fetal development. J. Nutr. 2004, 134, 2169–2172. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Boil. Chem. 1957, 226, 497–509. [Google Scholar]
- Rouser, G.; Fkeischer, S.; Yamamoto, A. Two dimensional then layer chromatographic separation of polar lipids and determination of phospholipids by phosphorus analysis of spots. Lipids 1970, 5, 494–496. [Google Scholar] [CrossRef]
- Gluckman, P.D.; Hanson, M.A. Living with the past: Evolution, development, and patterns of disease. Science 2004, 305, 1733–1736. [Google Scholar] [CrossRef]
- Wang, L.; Lin, Q.; Yang, T.; Liang, Y.; Nie, Y.; Luo, Y.; Shen, J.; Fu, X.; Tang, Y.; Luo, F. Oryzanol modifies high fat diet-induced obesity, liver gene expression profile, and inflammation response in mice. J. Agric. Food Chem. 2017, 65, 8374–8385. [Google Scholar] [CrossRef]
- Kondo, H.; Yoshida, H.; Mohri, T.; Kitagawa, H. Distribution of ferulic acid [2-14C] Esters (14C-FAE) in Rats. Pharmacomet. Oyo Yakuri 1968, 2, 29–32. [Google Scholar]
- Noda, H.; Shimizu, T.; Higuchi, K. Study on absorption, distribution, metabolism and excretion of ferulic acid triterpene alcohol ester. Kiso Rinsho 1974, 8, 35–42. [Google Scholar]
- Kozuka, C.; Sunagawa, S.; Ueda, R.; Higa, M.; Tanaka, H.; Shimizu-Okabe, C.; Ishiuchi, S.; Takayama, C.; Matsushita, M.; Tsutsui, M.; et al. gamma-Oryzanol protects pancreatic beta-cells against endoplasmic reticulum stress in male mice. Endocrinology 2015, 156, 1242–1250. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, M.; Ito, Y.; Nakakita, T.; Maehama, T.; Hieda, S.; Yamamoto, K.; Ueno, N. Clinical effect of gamma-oryzanol on climacteric disturbance -on serum lipid peroxides (author’s transl). Nihon Sanka Fujinka Gakkai Zasshi 1982, 34, 243–251. [Google Scholar] [PubMed]
- Jin Son, M.; Rico, C.W.; Hyun Nam, S.; Young Kang, M. Influence of Oryzanol and Ferulic acid on the lipid metabolism and antioxidative status in high fat-fed mice. J. Clin. Biochem. Nutr. 2010, 46, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Makynen, K.; Chitchumroonchokchai, C.; Adisakwattana, S.; Failla, M.; Ariyapitipun, T. Effect of gamma-oryzanol on the bioaccessibility and synthesis of cholesterol. Eur. Rev. Med. Pharmacol. Sci. 2012, 16, 49–56. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AIN93G | SO-HF | RBO-HF | HOZ-HF | |
|---|---|---|---|---|
| Ingredients (g %) | ||||
| Casein | 20.0 | 20.0 | 20.0 | 20.0 |
| L-Cystine | 0.3 | 0.3 | 0.3 | 0.3 |
| Corn Starch | 39.7 | 26.7 | 26.7 | 26.7 |
| Maltodextrin 10 | 13.2 | 13.2 | 13.2 | 13.2 |
| Sucrose | 10.0 | 10.0 | 10.0 | 10.0 |
| SO | 7.0 | 20.0 | - | - |
| RBO | - | - | 20.0 | - |
| HOZ | - | - | - | 20.0 |
| Cellulose, BW200 | 5.0 | 5.0 | 5.0 | 5.0 |
| Mineral Mix S10022G | 3.5 | 3.5 | 3.5 | 3.5 |
| Vitamin Mix V10037 | 1.0 | 1.0 | 1.0 | 1.0 |
| Choline bitartrate | 0.3 | 0.3 | 0.3 | 0.3 |
| t-Butylhydroquinone | 0.0014 | 0.0014 | 0.0014 | 0.0014 |
| Total | 100.0 | 100.0 | 100.0 | 100.0 |
| Centesimal Composition (kcal %) | ||||
| Protein | 20 | 17 | 17 | 17 |
| Carbohydrate | 64 | 44 | 44 | 44 |
| Fat | 16 | 39 | 39 | 39 |
| kcal/g | 4.4 | 5.1 | 5.1 | 5.1 |
| Groups | ||||
|---|---|---|---|---|
| AIN93G | SO-HF | RBO-HF | HOZ-HF | |
| Body weight (g) | 28.0 ± 0.5 | 29.5 ± 1.0 | 28.5 ± 0.5 | 27.7 ± 0.7 |
| Food intake (kcal/day) | 13.5 ± 0.2 | 13.0 ± 0.1 | 12.6 ± 0.5 | 13.8 ± 0.1 |
| Liver (g/100 g B.W.) | 3.80 ± 0.14 | 3.28 ± 0.15 | 3.79 ± 0.14 | 3.67 ± 0.21 |
| Spleen (g/100 g B.W.) | 0.35 ± 0.03 | 0.35 ± 0.05 | 0.35 ± 0.04 | 0.34 ± 0.04 |
| Pancreas (g/100 g B.W.) | 0.67 ± 0.10 | 0.84 ± 0.14 | 0.60 ± 0.03 | 0.72 ± 0.06 |
| Kidney (g/100 g B.W.) | 1.27 ± 0.06 | 1.29 ± 0.07 | 1.24 ± 0.05 | 1.31 ± 0.05 |
| Lung (g/100 g B.W.) | 0.87 ± 0.06 | 1.11 ± 0.11 | 0.89 ± 0.06 | 1.07 ± 0.10 |
| Heart (g/100 g B.W.) | 0.52 ± 0.02 | 0.52 ± 0.02 | 0.55 ± 0.03 | 0.57 ± 0.03 |
| Brain (g/100 g B.W.) | 1.53 ± 0.03 | 1.52 ± 0.04 | 1.56 ± 0.03 | 1.61 ± 0.05 |
| Mesenteric fat (g/100 g B.W.) | 0.61 ± 0.12 | 0.71 ± 0.16 | 0.47 ± 0.08 | 0.53 ± 0.07 |
| Perirenal fat (g/100 g B.W.) | 0.45 ± 0.10 ab | 0.88 ± 0.19 a | 0.60 ± 0.17 ab | 0.31 ± 0.08 b |
| Epidydimal fat (g/100 g B.W.) | 1.49 ± 0.23 | 2.30 ± 0.57 | 1.84 ± 0.50 | 1.14 ± 0.23 |
| Plasma TG (mg/100 g mL) | 141.3 ± 14.3 ab | 108.5 ± 7.5 ab | 144.8 ± 11.7 a | 102.0 ± 8.7 b |
| Plasma TC (mg/100 g mL) | 75.4 ± 5.1 | 66.2 ± 7.8 | 85.1 ± 2.4 | 62.2 ± 7.3 |
| Plasma PL (mg/100 g mL) | 36.8 ± 2.9 | 31.6 ± 3.3 | 39.0 ± 0.7 | 30.3 ± 3.0 |
| Plasma glucose (mg/100 g mL) | 86.8 ± 4.8 | 73.1 ± 11.2 | 88.8 ± 4.9 | 86.1 ± 5.0 |
| Liver TG (mg/g) | 53.8 ± 4.0 | 53.4 ± 3.8 | 42.4 ± 7.4 | 52.8 ± 6.1 |
| Liver TC (mg/g) | 3.9 ± 0.2 | 3.6 ± 0.1 | 3.8 ± 0.1 | 3.5 ± 0.2 |
| Liver PL (mg/g) | 4.0 ± 0.1 | 3.6 ± 0.1 | 3.8 ± 0.1 | 3.5 ± 0.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobayashi, E.; Ito, J.; Shimizu, N.; Kokumai, T.; Kato, S.; Sawada, K.; Hashimoto, H.; Eitsuka, T.; Miyazawa, T.; Nakagawa, K. Evaluation of γ-oryzanol Accumulation and Lipid Metabolism in the Body of Mice Following Long-Term Administration of γ-oryzanol. Nutrients 2019, 11, 104. https://doi.org/10.3390/nu11010104
Kobayashi E, Ito J, Shimizu N, Kokumai T, Kato S, Sawada K, Hashimoto H, Eitsuka T, Miyazawa T, Nakagawa K. Evaluation of γ-oryzanol Accumulation and Lipid Metabolism in the Body of Mice Following Long-Term Administration of γ-oryzanol. Nutrients. 2019; 11(1):104. https://doi.org/10.3390/nu11010104
Chicago/Turabian StyleKobayashi, Eri, Junya Ito, Naoki Shimizu, Takumi Kokumai, Shunji Kato, Kazue Sawada, Hiroyuki Hashimoto, Takahiro Eitsuka, Teruo Miyazawa, and Kiyotaka Nakagawa. 2019. "Evaluation of γ-oryzanol Accumulation and Lipid Metabolism in the Body of Mice Following Long-Term Administration of γ-oryzanol" Nutrients 11, no. 1: 104. https://doi.org/10.3390/nu11010104
APA StyleKobayashi, E., Ito, J., Shimizu, N., Kokumai, T., Kato, S., Sawada, K., Hashimoto, H., Eitsuka, T., Miyazawa, T., & Nakagawa, K. (2019). Evaluation of γ-oryzanol Accumulation and Lipid Metabolism in the Body of Mice Following Long-Term Administration of γ-oryzanol. Nutrients, 11(1), 104. https://doi.org/10.3390/nu11010104