Comparative Effects of Native and Defatted Flaxseeds on Intestinal Enzyme Activity and Lipid Metabolism in Rats Fed a High-Fat Diet Containing Cholic Acid

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Flaxseeds
2.2. Animal and Diets
2.3. Collection of Biological Material and Analytical Procedures
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hall, C., III; Tulbek, M.; Xu, Y. Flaxseed. Adv. Food Nutr. Res. 2006, 51, 1–97. [Google Scholar] [PubMed]
- McDowell, D.; Elliot, C.T.; Koidis, A. Pre-processing effects on cold press rapeseed oil quality indicators and phenolic compounds. Eur. J. Lipid Sci. Technol. 2017, 119, 1600357. [Google Scholar] [CrossRef]
- Morris, D.H. Flax—A Health and Nutrition Primer. 2007. Available online: https://flaxcouncil.ca/resources/nutrition/technical-nutrition-information/flax-a-health-and-nutrition-primer/ (accessed on 18 October 2017).
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Clarke, S.D. Polyunsaturated fatty acid regulation of gene transcription: A molecular mechanism to improve the metabolic syndrome. J. Nutr. 2001, 131, 1129–1132. [Google Scholar] [CrossRef] [PubMed]
- Lunn, J.; Theobald, H.E. The health effects of dietary unsaturated fatty acids. Nutr. Bull. 2006, 31, 178–224. [Google Scholar] [CrossRef] [Green Version]
- Kajla, P.; Sharma, A.; Sood, D.R. Flaxseed—A potential functional food source. J. Food Sci. Technol. 2015, 52, 1857–1871. [Google Scholar] [CrossRef] [PubMed]
- Marambe, P.W.M.L.H.K.; Shand, P.J.; Wanasundara, J.P.D. An in-vitro investigation of selected biological activities of hydrolysed flaxseed (Linum usitatissium L.) proteins. J. Am. Oil Chem. Soc. 2008, 85, 1155–1164. [Google Scholar] [CrossRef]
- Omoni, A.O.; Aluko, R.E. Effect of cationic flaxseed protein hydrolysate fractions on the in vitro structure and activity of calmodulin-dependent endothelial nitric oxide synthase. Mol. Nutr. Food Res. 2006, 50, 958–966. [Google Scholar] [CrossRef] [PubMed]
- Undenigwe, C.C.; Lu, Y.L.; Han, C.H.; Hou, W.C.; Aluko, R.E. Flaxseed protein-derived peptide fractions: Antioxidant properties and inhibition of lipopolysaccharide-induced nitric oxide production in murine macrophages. Food Chem. 2009, 116, 277–284. [Google Scholar] [CrossRef]
- Kristensen, M.; Savorani, F.; Christensen, S.; Engelsen, S.B.; Bügel, S.; Toubro, S.; Tenes, L.; Astrup, A. Flaxseed dietary fibers suppress postprandial lipemia and appetite sensation in young men. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, M.; Knudsen, K.E.B.; Jorgensen, H.; Oomah, D.; Bügel, S.; Toubro, S.; Tetens, I.; Astrup, A. Linseed dietary fibers reduce apparent digestibility of energy and fat and weight gain in growing rats. Nutrients 2013, 5, 3287–3298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boban, P.T.; Nambisan, B.; Sudhakaran, P.R. Hypolipidaemic effect of chemically different mucilages in rats: A comparative study. Br. J. Nutr. 2006, 96, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Kay, B.A.; Trigatti, K.; MacNeil, M.B.; Klingel, S.L.; Repin, N.; Goff, H.D.; Wright, A.J.; Duncan, A.M. Pudding products enriched with yellow mustard mucilage, fenugreek gum of flaxseed mucilage and matched for simulated intestinal viscosity significantly reduce postprandial peak glucose and insulin in adults at risk for type 2 diabetes. J. Funct. Food 2017, 37, 603–611. [Google Scholar] [CrossRef]
- Goyal, A.; Sharma, V.; Upadhyay, N.; Gill, S.; Sihag, M. Flax and flaxseed oil: An ancient medicine & modern functional food. J. Food Sci. Technol. 2014, 51, 1633–1653. [Google Scholar] [PubMed]
- Reeves, P.C. Components of the AIN-93 diets as improvements in the AIN-76A diet. J. Nutr. 1997, 127, 838–841. [Google Scholar] [CrossRef] [PubMed]
- Jurgoński, A.; Juśkiewicz, J.; Zduńczyk, Z. An anthocyanin-rich extract from Kamchatka honeysuckle increases enzymatic activity within the gut and ameliorates abnormal lipid and glucose metabolism in rats. Nutrition 2013, 29, 898–902. [Google Scholar] [CrossRef] [PubMed]
- Barczynska, R.; Jurgoński, A.; Slizewska, K.; Juśkiewicz, J.; Kapusniak, J. Effects of potato dextrin on the composition and metabolism of the gut microbiota in rats fed standard and high-fat diets. J. Funct. Food 2017, 34, 398–407. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2006, 6, 3159–3165. [Google Scholar] [CrossRef] [PubMed]
- Botsoglou, N.A.; Fletouris, D.J.; Papageorgiou, G.E.; Vassilopoulos, V.N.; Mantys, A.J.; Trakatellis, A.G. Rapid, Sensitive, and Specific Thiobarbituric Acid Metod for Measuring Lipid Peroxidation in Animal Tissue, Food, and Feedstuff Samples. J. Agric. Food Chem. 1994, 42, 1931–1937. [Google Scholar] [CrossRef]
- Stajic, S.; Stanisic, N.; Levic, S.; Tomovic, V.; Lilic, S.; Vranic, D.; Jokanovic, M.; Zivkovic, D. Physico-chemical characteristics and sensory quality of dry fermented sausages with flaxseed oil preparations. Pol. J. Food Nutr. Sci. 2018, 68. [Google Scholar] [CrossRef]
- Vial, G.; Dubouchaud, H.; Couturier, K.; Cottet-Rousselle, C.; Taleux, N.; Athias, A.; Galinier, A.; Casteilla, L.; Leverve, X.M. Effects of a high-fat diet on energy metabolism and ROS production in rat liver. J. Hepatol. 2011, 54, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Quesada, H.; del Bas, J.M.; Pajuelo, D.; Diaz, S.; Fernandez-Larrea, J.; Pintent, M.; Arola, L.; Salvado, M.J. Grape seed proanthocyanidins correct dyslipidemia associated with a high-fat diet in rats and repress genes controlling lipogenesis and VLDL assembling in liver. Int. J. Obes. 2009, 33, 1007–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fotschki, B.; Juśkiewicz, J.; Jurgoński, A.; Rigby, N.; Sójka, M.; Kołodziejczyk, K.; Mackie, A.; Zduńczyk, Z. Raspberry pomace alters cecal microbial activity and reduces secondary bile acids in rats fed a high-fat diet. J. Nutr. Biochem. 2017, 46, 13–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmann, A.F.; Hagey, L.R. Bile acids: Chemistry, pathochemistry, biology, pathobiology, and therapeutics. Cell. Mol. Life Sci. 2008, 65, 2461–2483. [Google Scholar] [CrossRef] [PubMed]
- Jurgoński, A.; Juśkiewicz, J.; Sójka, M.; Karlińska, E. Diet-induced disorders in rats are more efficiency attenated by initial rather than delayed supplementation with polyphenol-rich berry fibers. J. Funct. Food 2016, 22, 556–564. [Google Scholar] [CrossRef]
- Cui, W.; Kenaschuk, E.; Mazza, G. Influence of genotype on chemical composition and rheological properties of flaxseed gums. Food Hydrocolloid 1996, 10, 221–227. [Google Scholar] [CrossRef]
- Alzueta, C.; Rodriguez, M.L.; Cutuli, M.T.; Rebole, A.; Ortiz, L.T.; Centeno, C.; Trevino, J. Effect of whole and decmucilaged linseed in broiler chicken diets on digesta viscosity, nutrient utilization and intestinal microflora. Br. Poult. Sci. 2003, 44, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Gill, C.I.; Rowland, I.R. Diet and cancer: Assessing the risk. Br. J. Nutr. 2002, 88, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Kaur, M.; Gill, B.S. Phenolic acid composition of flaxseed cultivars by ultra-performance liquid chromatography (UPLC) and their antioxidant activities: Effect of sand roasting and microwave heating. J. Food Process. Preserv. 2017, 45, e13181. [Google Scholar] [CrossRef]
- Alu’datt, M.H.; Rababah, T.; Ereifej, K.; Alli, L. Distribution, antioxidant and characterization of phenolic compounds in soybean, flaxseed and olives. Food Chem. 2013, 139, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Patsouris, D.; Reddy, J.K.; Muller, M.; Kersten, S. Peroxisome proliferator-acivated receptor α mediates the effect of high-fat diet on hepatic gene expression. Endocrinology 2006, 147, 1508–1516. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Olson, P.; Evans, R.M. Minireview: Lipid metabolism, metabolic diseases, and peroxisome proliferator-activated receptors. Endocrinology 2003, 144, 2201–2207. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Tang, Y.; Yu, X.; Liu, P.; Xiao, L.; Liu, L.; Deng, Q.; Yao, P. Flaxseed lignan alleviate high fat diet-induced hepatic steatosis and insulin resistance in mice: Potential involvement of AMP-activated protein kinase. J. Funct. Food 2016, 24, 482–491. [Google Scholar] [CrossRef]
- Pacheco, J.T.; Daleprame, J.B.; Boaventura, G.T. Impact of dietary flaxseed (Linum usitatissimum) supplementation on biochemical profile in healthy rats. Nutr. Hosp. 2011, 26, 798–802. [Google Scholar]
- Ribeiro, D.C. Assessment of body composition and bone parameters of lactating rats treated with diet containing flaxseed meal (Linum usitatissimum) during post-weaning period. Nutr. Hosp. 2014, 30, 366–371. [Google Scholar]
- Wu, H.; Dwyer, K.M.; Fan, Z.; Shircore, A.; Fan, J.; Dwyer, J.H. Dietary fiber and progression of atherosclerosis: The Los Angeles atherosclerosis study. Am. J. Clin. Nutr. 2003, 78, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Gunness, P.; Gidley, M.J. Mechanisms underlying the cholesterol-lowering properties of soluble dietary fibre polysaccharides. Food Funct. 2010, 1, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Zanwar, A.A.; Hegde, M.V.; Bodhankar, S.L. Antihyperlipidemic effect of flax lignan concentrate in triton induced hyperlipidemic rats. Int. J. Pharmacol. 2012, 8, 355–363. [Google Scholar]
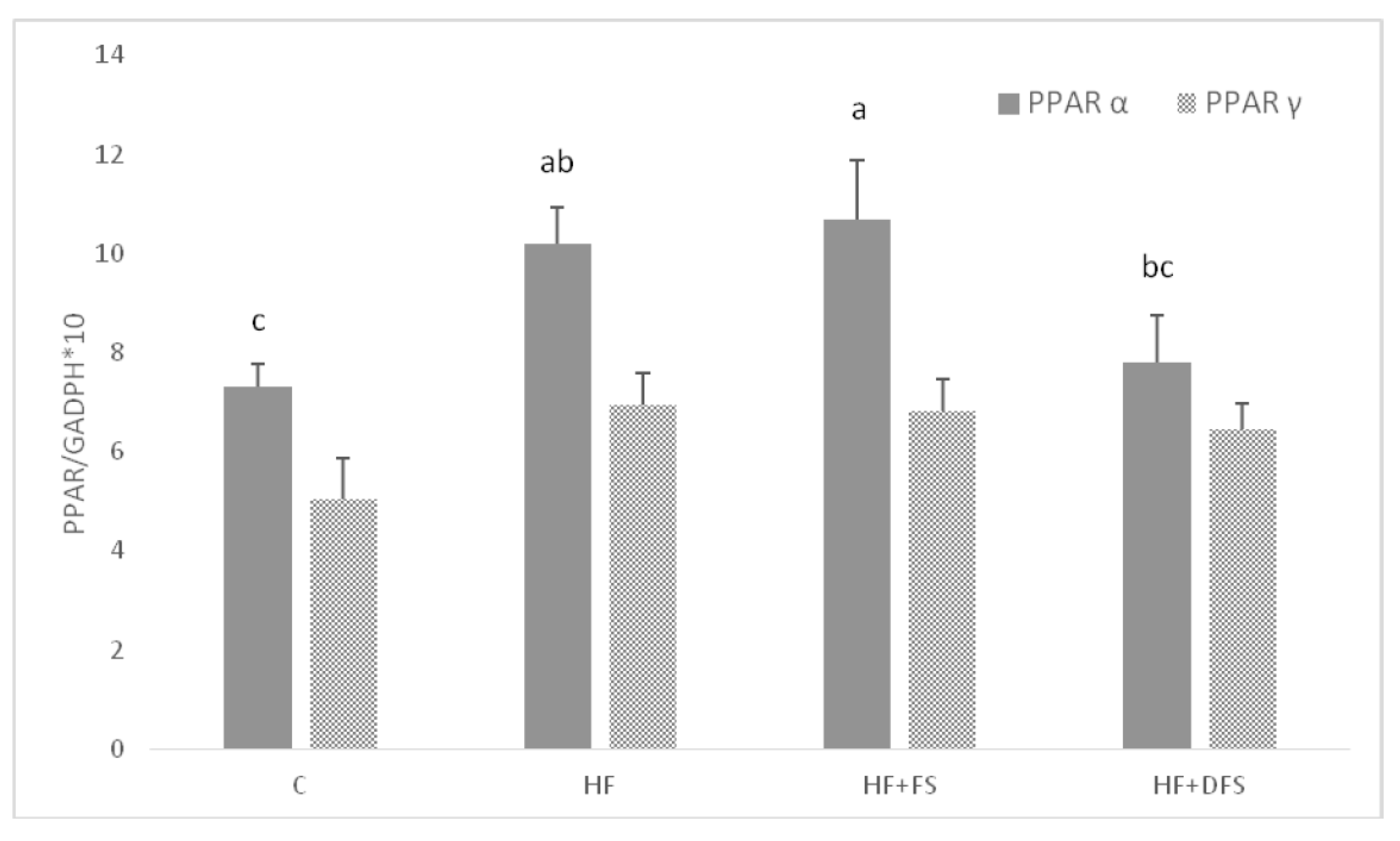

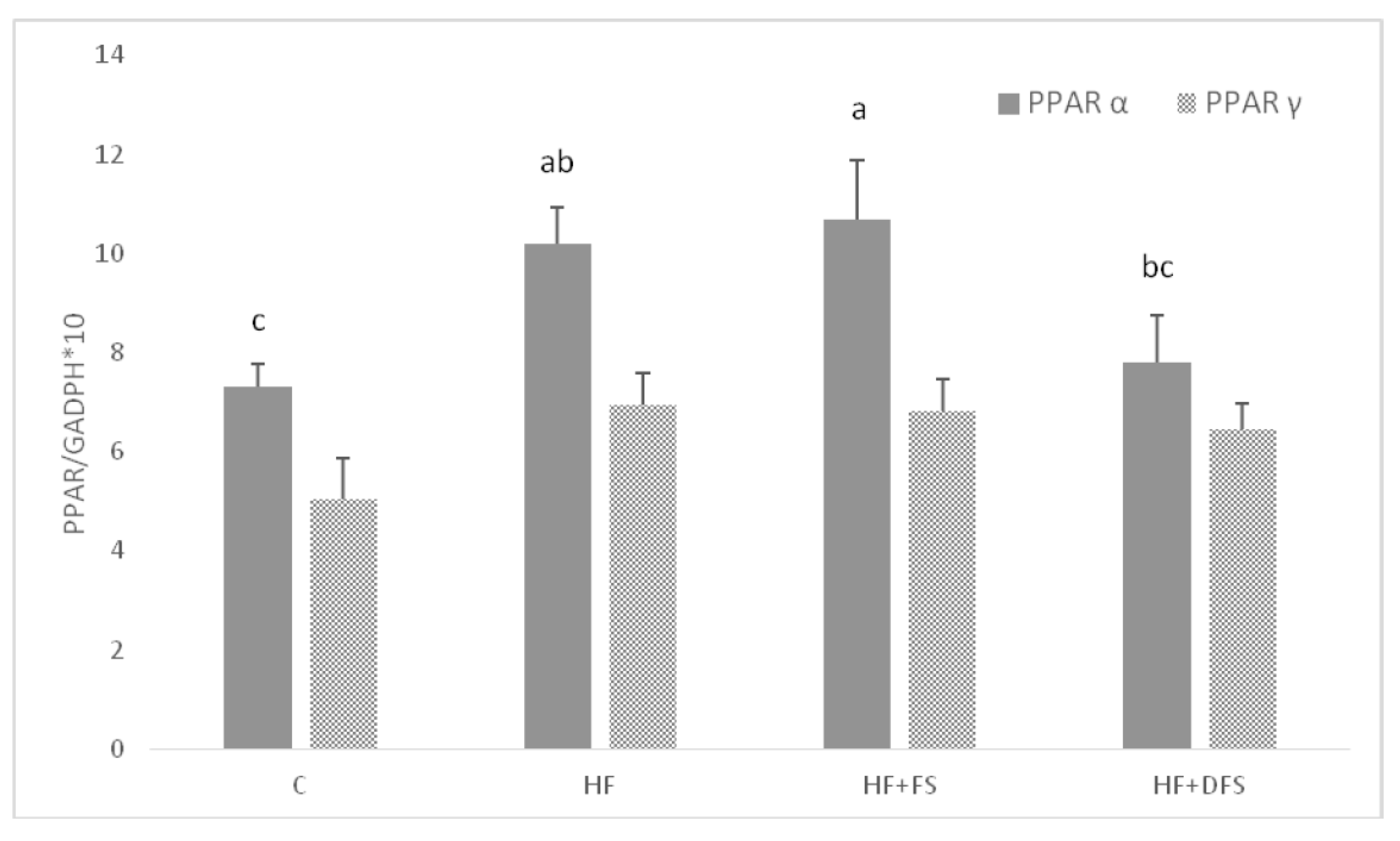{kind=link}
| Diet Components | C | HF | HF + FS | HF + DFS |
|---|---|---|---|---|
| Casein | 20 | 20 | 20 | 20 |
| dl-methionine | 0.3 | 0.3 | 0.3 | 0.3 |
| Rapeseed oil | 7 | 7 | 7 | 7 |
| Lard | - | 14 | 14 | 14 |
| Cholic acid | - | 0.1 | 0.1 | 0.1 |
| Corn starch | 53.0 | 38.9 | 37.9 | 37.9 |
| Flaxseeds 1 | - | - | 1 | - |
| Defatted flaxseeds 2 | - | - | - | 1 |
| Saccharose | 10 | 10 | 10 | 10 |
| Cellulose | 5 | 5 | 5 | 5 |
| Mineral mix 3 | 3.5 | 3.5 | 3.5 | 3.5 |
| Vitamin mix 3 | 1 | 1 | 1 | 1 |
| Choline chloride | 0.2 | 0.2 | 0.2 | 0.2 |
| Indices | C | HF | HF + FS | HF + DFS | ANOVA p Value |
|---|---|---|---|---|---|
| Initial BW, g | 290 ± 2.2 | 296 ± 2.5 | 295 ± 2.2 | 295 ± 3.0 | NS |
| Final BW, g | 404 ± 5.1 b | 429 ± 5.4 a | 423 ± 8.8 a,b | 429 ± 6.1 a | <0.05 |
| Small intestine | |||||
| Mass with digesta, g/100 g BW | 1.44 ± 0.025 b | 1.52 ± 0.044 a,b | 1.62 ± 0.041 a | 1.52 ± 0.035 a,b | <0.05 |
| pH of digesta | 7.31 ± 0.134 a | 6.85 ± 0.088 b | 6.90 ± 0.107 b | 6.72 ± 0.133 b | <0.01 |
| Digesta viscosity, mPa*s | 2.01 ± 0.169 | 1.91 ± 0.130 | 1.74 ± 0.133 | 1.63 ± 0.188 | NS |
| Mucosal disaccharidase activity 1 | |||||
| Sucrase | 18.5 ± 2.46 b | 23.3 ± 3.45 b | 24.5 ± 3.84 a,b | 35.8 ± 5.31 a | <0.05 |
| Maltase | 81.0 ± 12.6 b | 102.6 ± 15.7 b | 105.4 ± 16.0 b | 158.3 ± 22.1 a | <0.05 |
| Lactase | 2.95 ± 0.581 b | 3.03 ± 0.433 b | 3.58 ± 0.442 b | 5.47 ± 0.500 a | <0.01 |
| Caecum | |||||
| Tissue mass, g/100 g BW | 0.155 ± 0.004 | 0.141 ± 0.008 | 0.151 ± 0.003 | 0.159 ± 0.005 | NS |
| Digesta mass, g/100 g tissue | 1.69 ± 0.156 | 1.62 ± 0.152 | 1.40 ± 0.224 | 1.23 ± 0.151 | NS |
| pH of digesta | 7.45 ± 0.091 | 7.56 ± 0.073 | 7.57 ± 0.109 | 7.41 ± 0.100 | NS |
| Ammonia, mg/g digesta | 0.314 ± 0.027 | 0.366 ± 0.050 | 0.328 ± 0.022 | 0.340 ± 0.019 | NS |
| SCFA concentration 2 | 65.7 ± 3.89 | 65.8 ± 3.35 | 64.6 ± 5.68 | 62.3 ± 3.78 | NS |
| Colon | |||||
| Tissue mass, g/100 g BW | 0.259 ± 0.019 | 0.262 ± 0.017 | 0.253 ± 0.013 | 0.232 ± 0.009 | NS |
| Digesta mass, g/100 g tissue | 0.835 ± 0.122 a | 0.785 ± 0.124 a | 0.469 ± 0.073 b | 0.642 ± 0.043 a | <0.05 |
| pH of digesta | 7.23 ± 0.069 | 7.21 ± 0.114 | 7.22 ± 0.097 | 7.02 ± 0.088 | NS |
| Digesta bacterial enzymes activity 3 | |||||
| α-glucosidase | 31.1 ± 2.73 a | 21.9 ± 1.19 b | 29.4 ± 1.78 a | 28.2 ± 1.78 a | <0.05 |
| β-glucosidase | 6.79 ± 0.77 b | 3.65 ± 0.54 b | 10.10 ± 0.98 a | 13.26 ± 1.70 a | <0.001 |
| α-galactosidase | 22.7 ± 1.34 a | 15.6 ± 1.15 b | 25.3 ± 2.09 a | 21.7 ± 1.62 a | <0.01 |
| β-galactosidase | 100 ± 10.3 b | 92 ± 3.0 b | 133 ± 10.1 a | 129 ± 7.7 a | <0.01 |
| β-glucuronidase | 62.2 ± 7.72 b | 91.2 ± 4.97 a | 63.4 ± 6.18 b | 88.7 ± 12.29 a | <0.05 |
| Indices | C | HF | HF + FS | HF + DFS | ANOVA p Value |
|---|---|---|---|---|---|
| Liver markers | |||||
| Liver mass, g/100 g BW | 2.31 ± 0.029 | 2.47 ± 0.064 | 2.73 ± 0.166 | 2.66 ± 0.196 | NS |
| Liver fat, % | 8.09 ± 0.476 b | 13.85 ± 1.753 a | 12.10 ± 1.900 a,b | 12.41 ± 1.150 a,b | <0.05 |
| Liver cholesterol, mg/g | 3.86 ± 0.452 b | 14.37 ± 0.749 a | 13.14 ± 0.736 a | 13.63 ± 1.036 a | <0.001 |
| Liver triglycerides, mg/g | 25.8 ± 1.42 c | 50.7 ± 3.22 a | 40.2 ± 3.21 b | 40.0 ± 3.02 b | <0.001 |
| Serum ALT, U/L | 93.2 ± 5.83 | 108.0 ± 5.42 | 115.1 ± 10.43 | 90.9 ± 6.34 | NS |
| Serum AST, U/L | 20.1 ± 1.56 b | 32.1 ± 5.14 b | 43.3 ± 8.18 a | 28.2 ± 3.90 b | <0.05 |
| Liver TBARS, µg/g | 2.33 ± 0.178 | 2.69 ± 0.311 | 2.80 ± 0.369 | 2.44 ± 0.143 | NS |
| Liver GSH/GSSG, µmol/g | 9.68 ± 0.485 | 10.74 ± 0.354 | 11.95 ± 1.209 | 10.35 ± 1.185 | NS |
| Kidney markers | |||||
| Kidneys mass, g/100 g BW | 0.516 ± 0.011 a | 0.454 ± 0.011 b | 0.492 ± 0.019 a,b | 0.505 ± 0.016 a | <0.05 |
| Serum creatinine, µmol/L | 28.5 ± 4.32 | 26.1 ± 3.39 | 27.5 ± 3.76 | 25.7 ± 2.56 | NS |
| Serum urea, mmol/L | 5.92 ± 0.307 | 5.62 ± 0.298 | 6.24 ± 0.167 | 5.45 ± 0.325 | NS |
| Kidney TBARS, µg/g | 3.51 ± 0.333 a | 3.65 ± 0.371 a | 2.43 ± 0.428 b | 2.66 ± 0.226 a,b | <0.05 |
| Indices | C | HF | HF + FS | HF + DFS | ANOVA p Value |
|---|---|---|---|---|---|
| Total cholesterol, mmol/L | 1.90 ± 0.109 | 1.88 ± 0.126 | 1.91 ± 0.090 | 1.83 ± 0.073 | NS |
| HDL cholesterol, mmol/L | 0.615 ± 0.027 a | 0.434 ± 0.042 b | 0.521 ± 0.029 a,b | 0.534 ± 0.025 a | <0.01 |
| LDL cholesterol, mmol/L | 0.129 ± 0.013 | 0.180 ± 0.016 | 0.159 ± 0.026 | 0.163 ± 0.009 | NS |
| Triglycerides, mmol/L | 0.848 ± 0.115 | 0.736 ± 0.077 | 0.740 ± 0.060 | 0.766 ± 0.060 | NS |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Opyd, P.M.; Jurgoński, A.; Juśkiewicz, J.; Fotschki, B.; Koza, J. Comparative Effects of Native and Defatted Flaxseeds on Intestinal Enzyme Activity and Lipid Metabolism in Rats Fed a High-Fat Diet Containing Cholic Acid. Nutrients 2018, 10, 1181. https://doi.org/10.3390/nu10091181
Opyd PM, Jurgoński A, Juśkiewicz J, Fotschki B, Koza J. Comparative Effects of Native and Defatted Flaxseeds on Intestinal Enzyme Activity and Lipid Metabolism in Rats Fed a High-Fat Diet Containing Cholic Acid. Nutrients. 2018; 10(9):1181. https://doi.org/10.3390/nu10091181
Chicago/Turabian StyleOpyd, Paulina M., Adam Jurgoński, Jerzy Juśkiewicz, Bartosz Fotschki, and Jarosław Koza. 2018. "Comparative Effects of Native and Defatted Flaxseeds on Intestinal Enzyme Activity and Lipid Metabolism in Rats Fed a High-Fat Diet Containing Cholic Acid" Nutrients 10, no. 9: 1181. https://doi.org/10.3390/nu10091181
APA StyleOpyd, P. M., Jurgoński, A., Juśkiewicz, J., Fotschki, B., & Koza, J. (2018). Comparative Effects of Native and Defatted Flaxseeds on Intestinal Enzyme Activity and Lipid Metabolism in Rats Fed a High-Fat Diet Containing Cholic Acid. Nutrients, 10(9), 1181. https://doi.org/10.3390/nu10091181