Pistachio Consumption Prevents and Improves Lipid Dysmetabolism by Reducing the Lipid Metabolizing Gene Expression in Diet-Induced Obese Mice

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Prevention Study
2.3. Reversal Study
2.4. Biochemical Analysis
2.5. Micro-Computed X-Ray Tomography
2.6. Histological Analysis
2.7. Quantification of Hepatic Lipids
2.8. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.9. Statistical Analyses
3. Results
3.1. Prevention Study
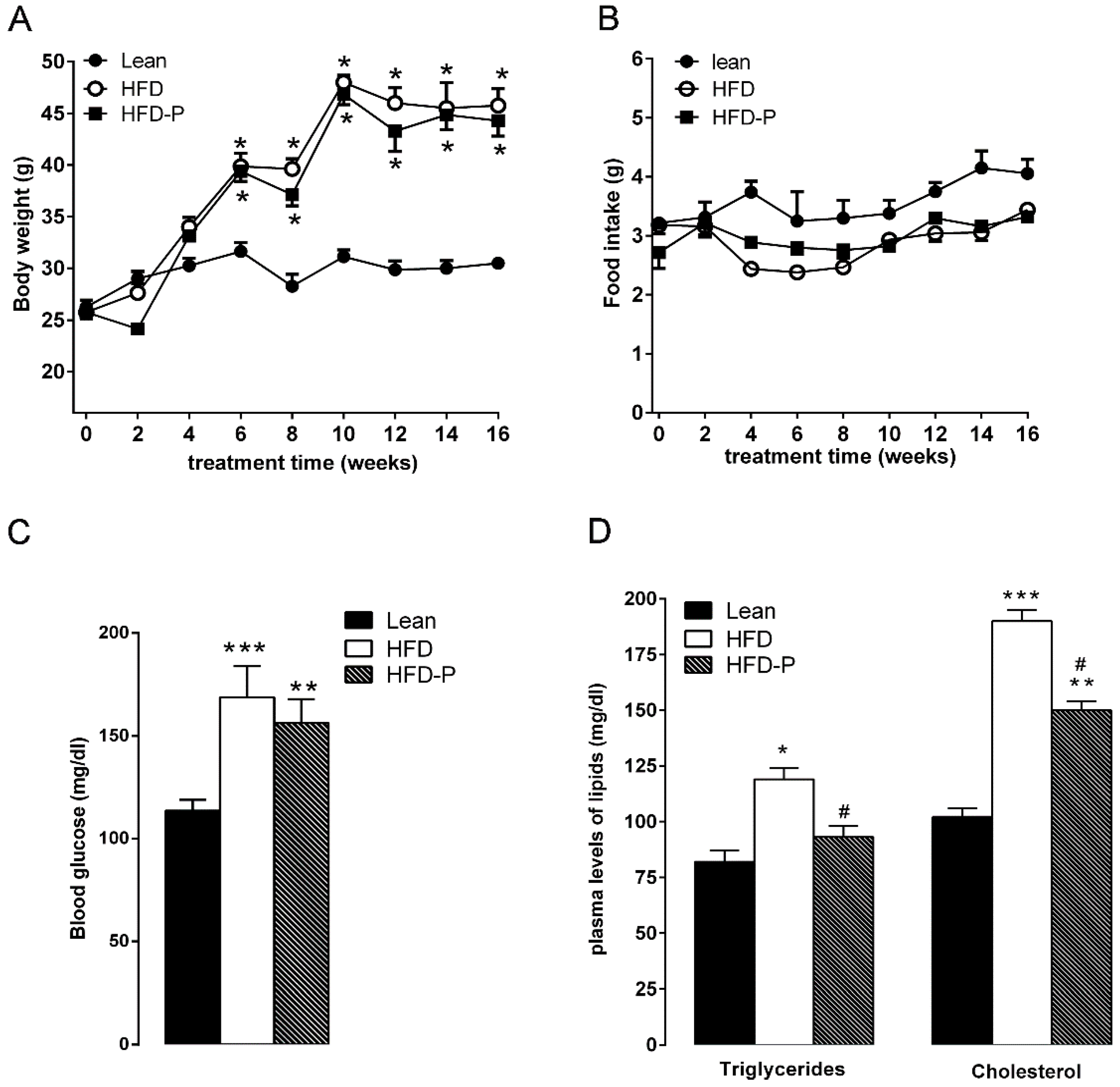
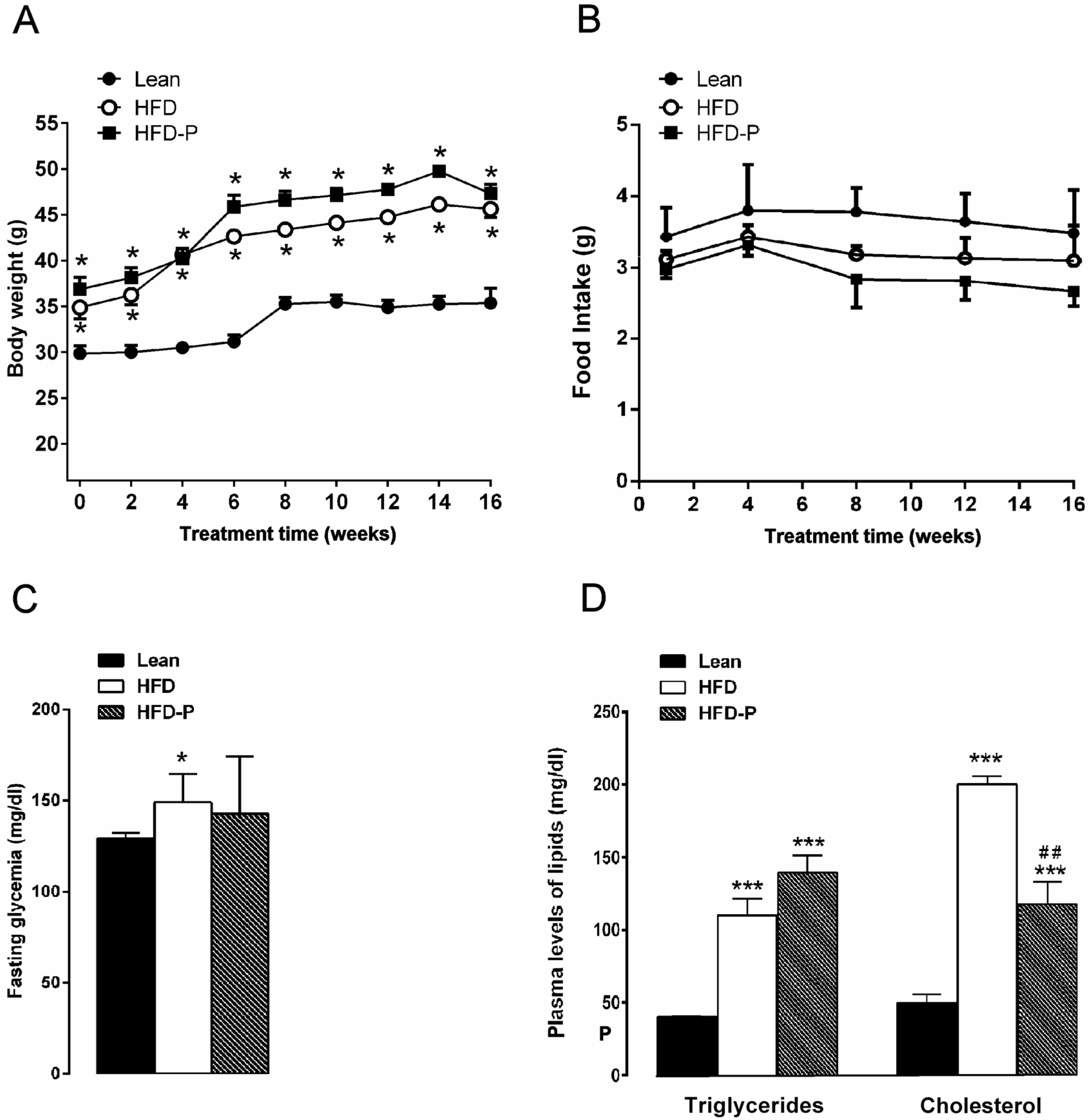
3.1.1. Effect of Pistachio Consumption on Metabolic Parameters
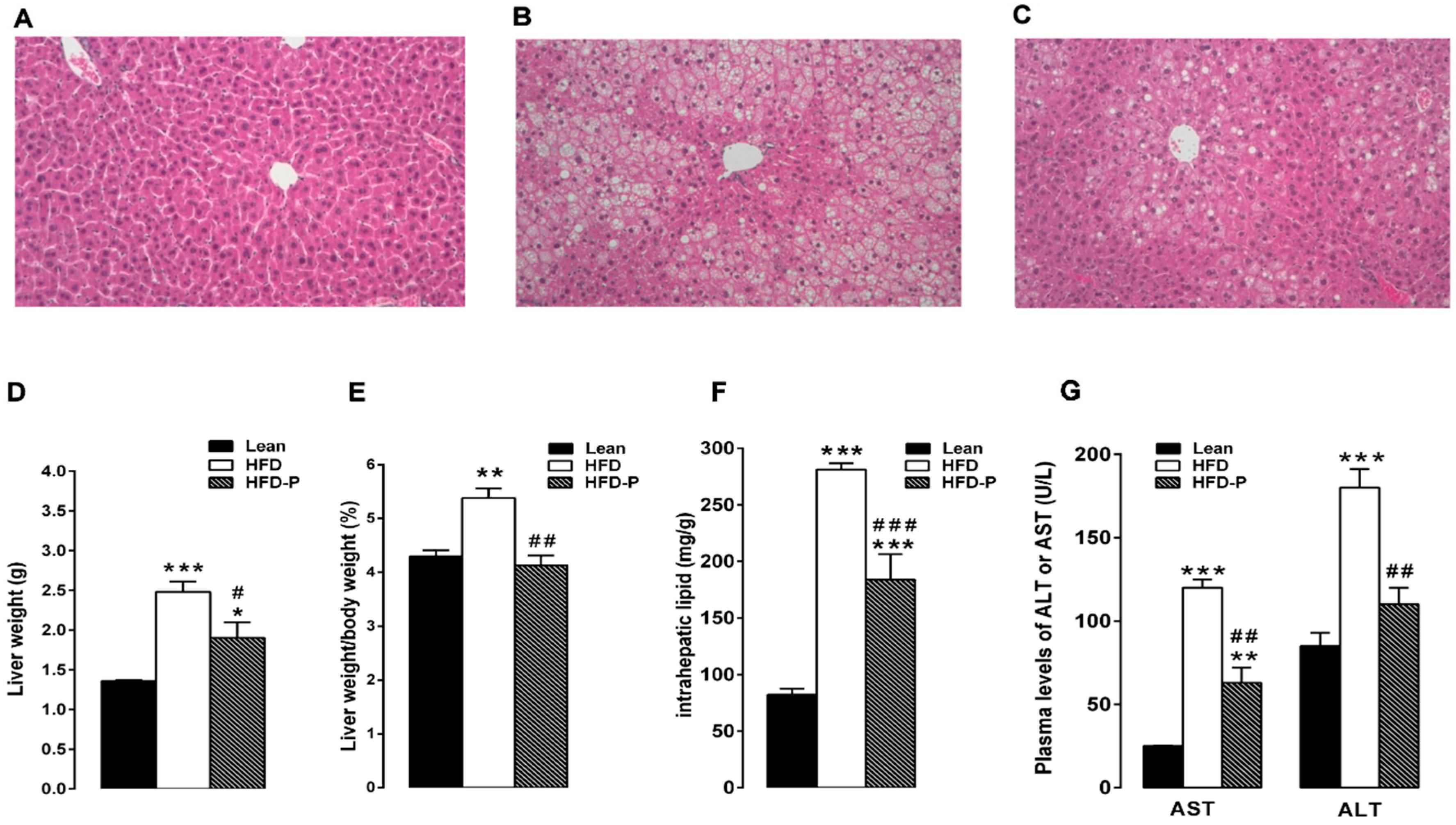
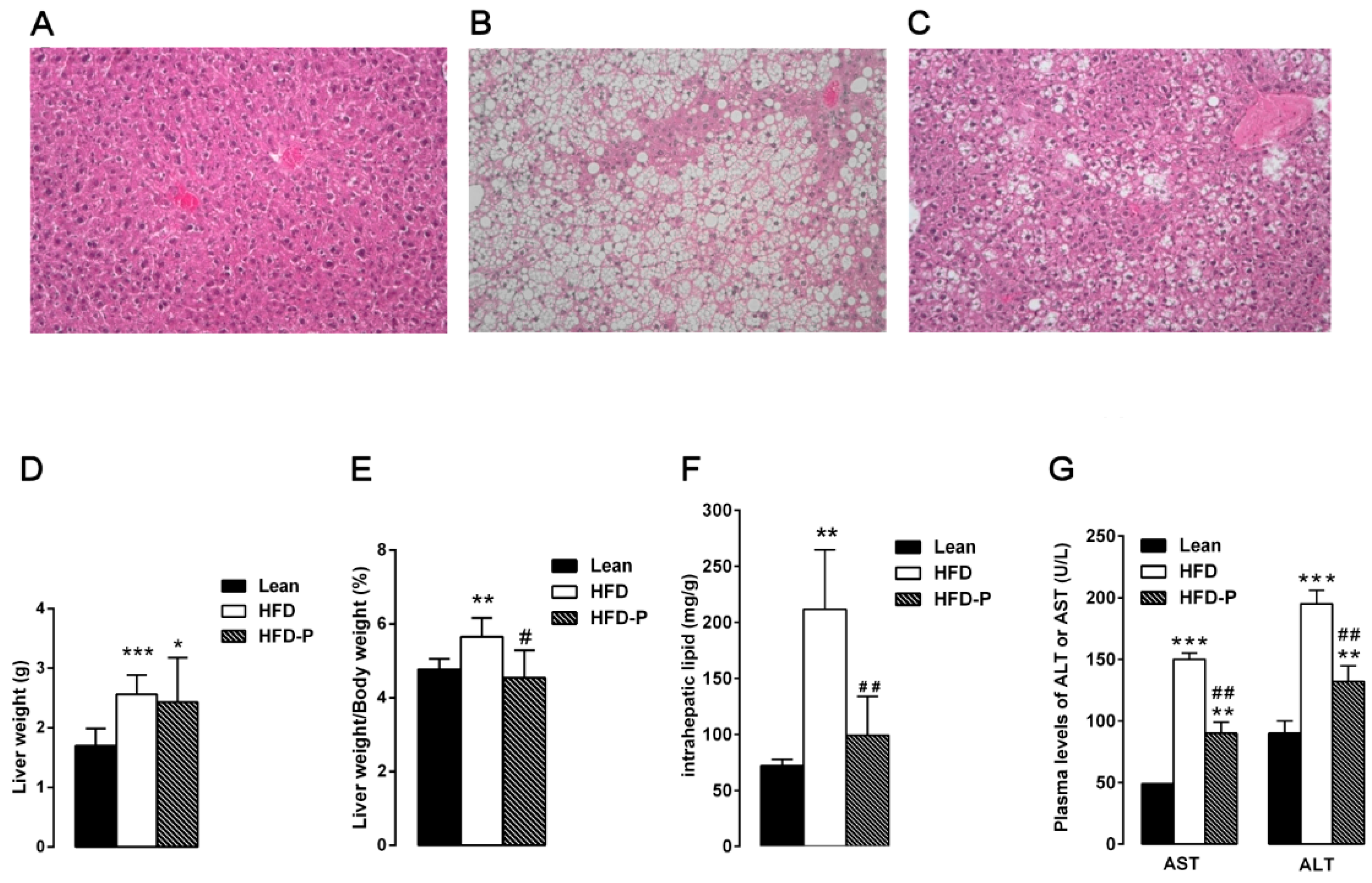
3.1.2. Pistachio Consumption and Liver Steatosis
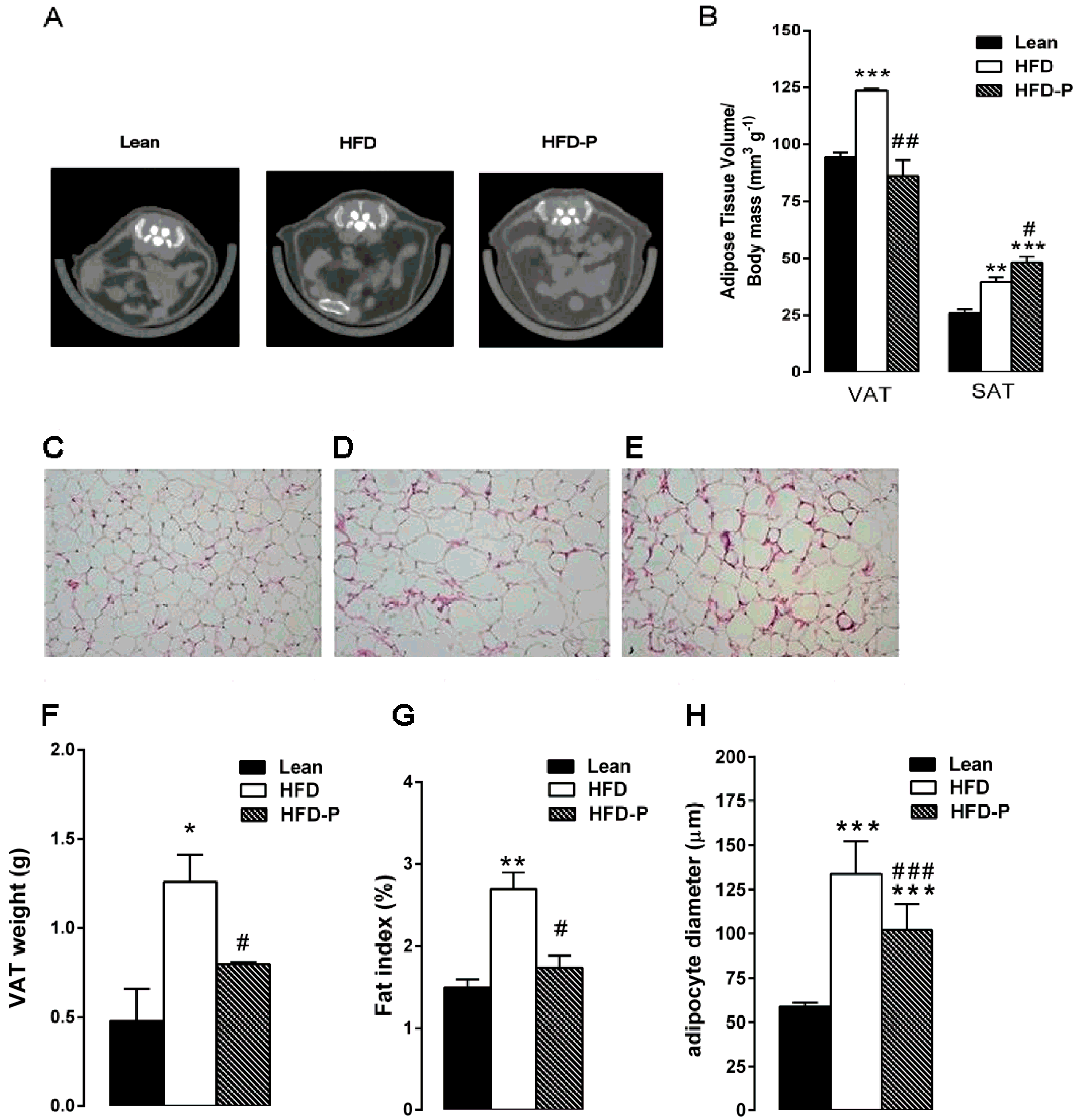
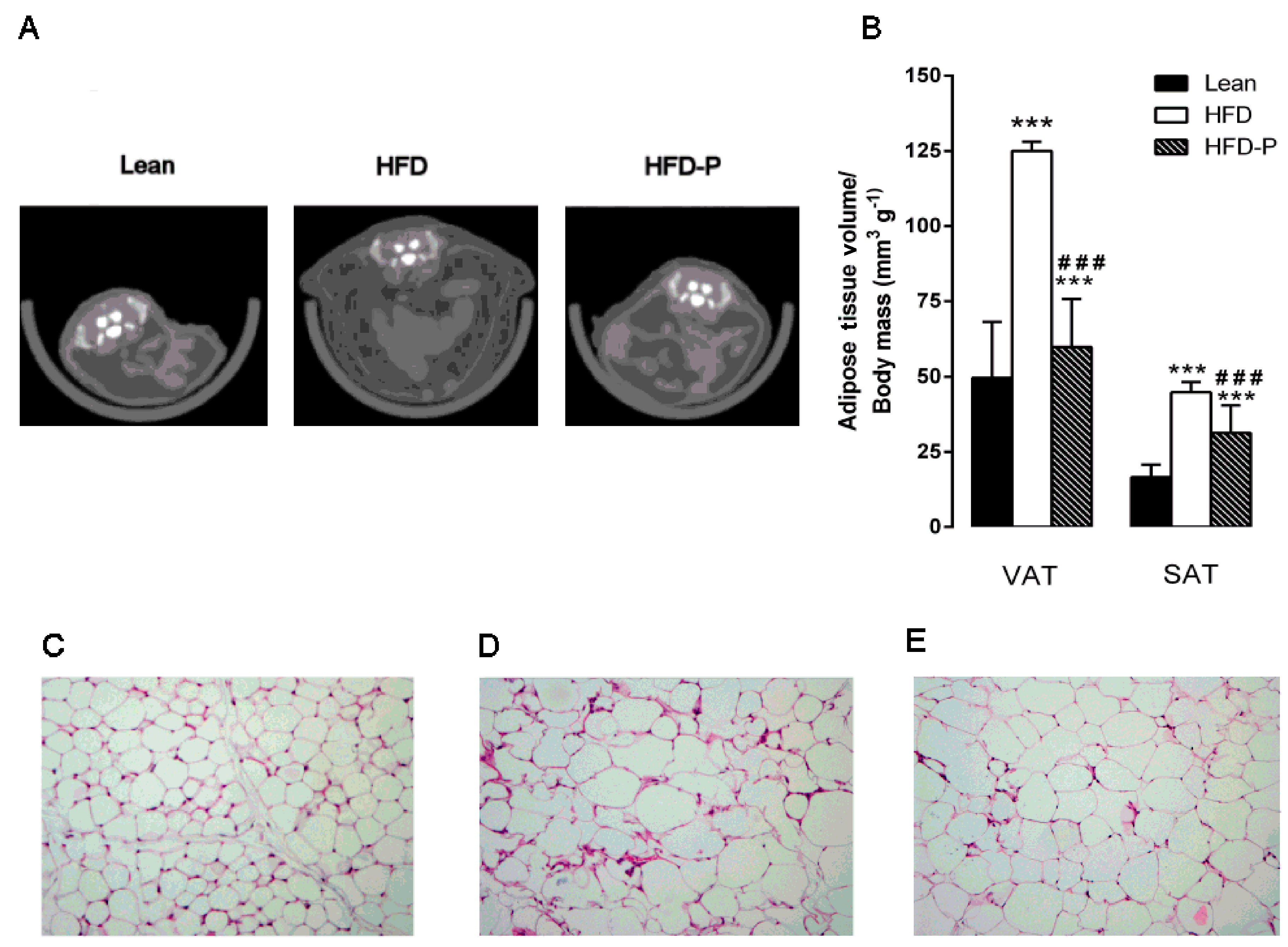
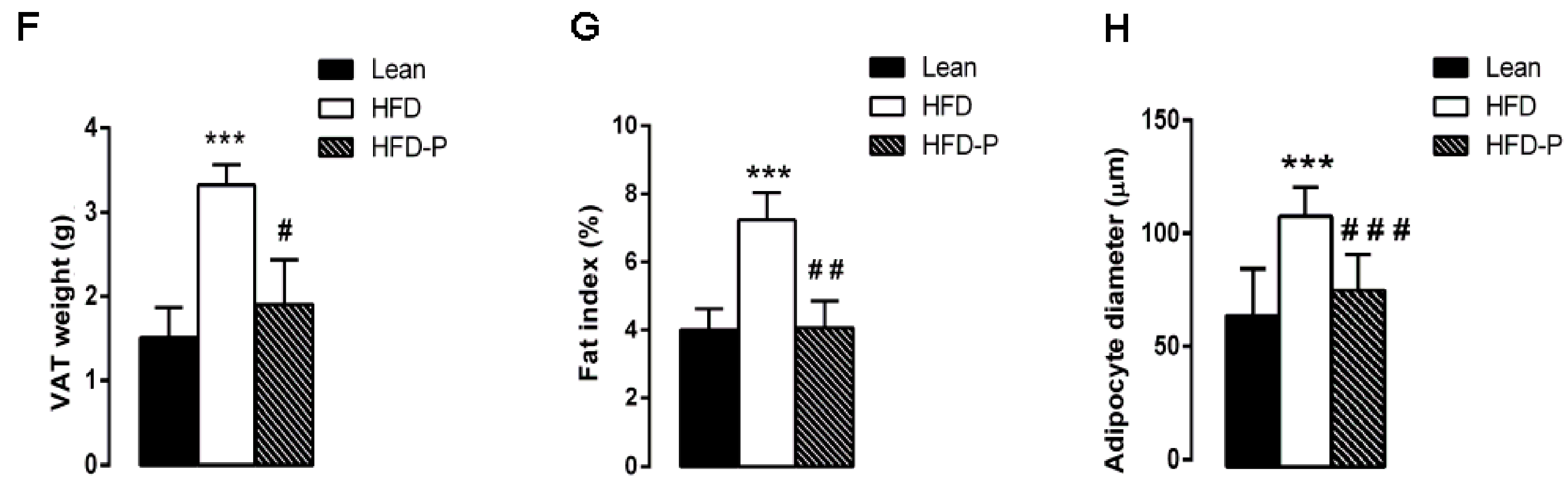
3.1.3. Pistachio Consumption and HFD-Induced Adipose Tissue Alterations
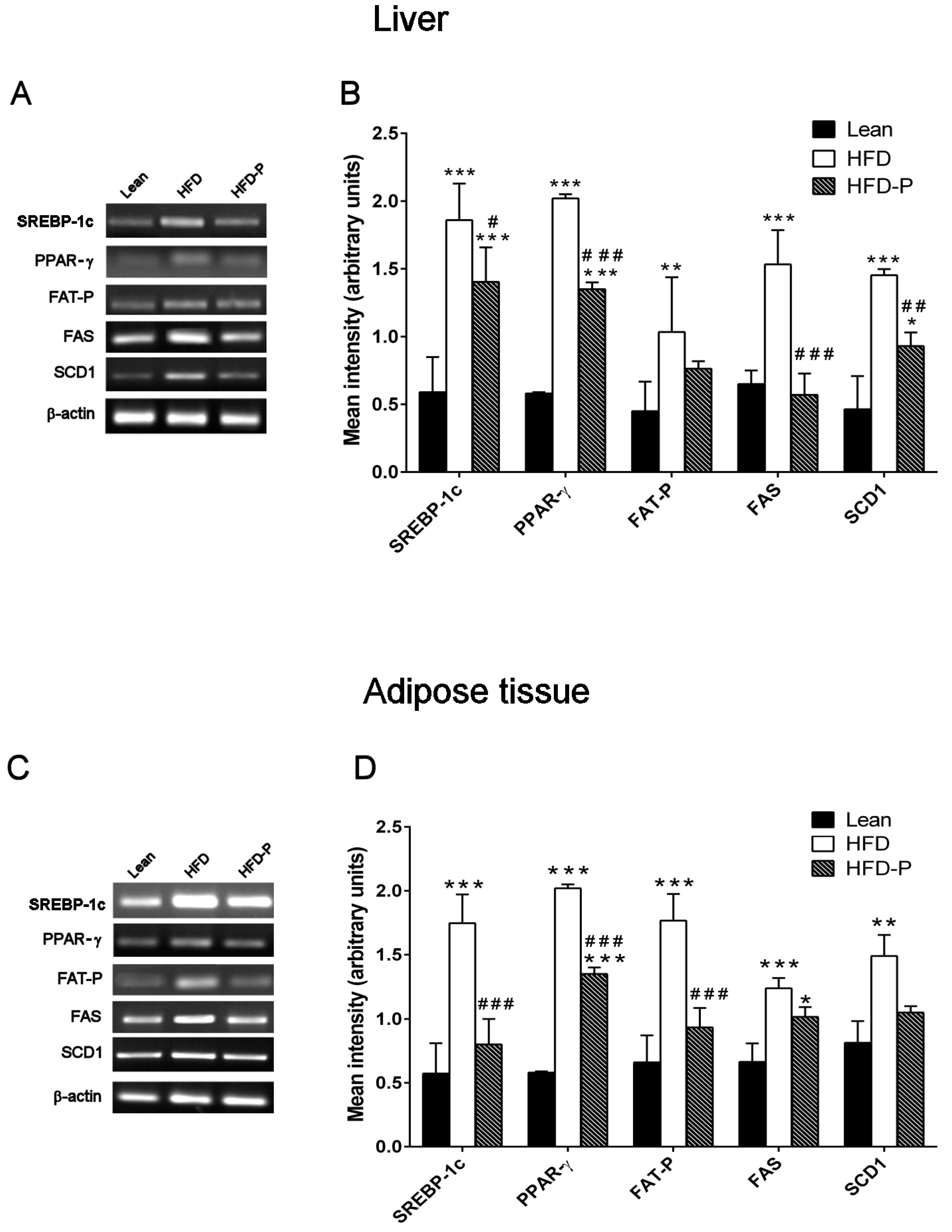
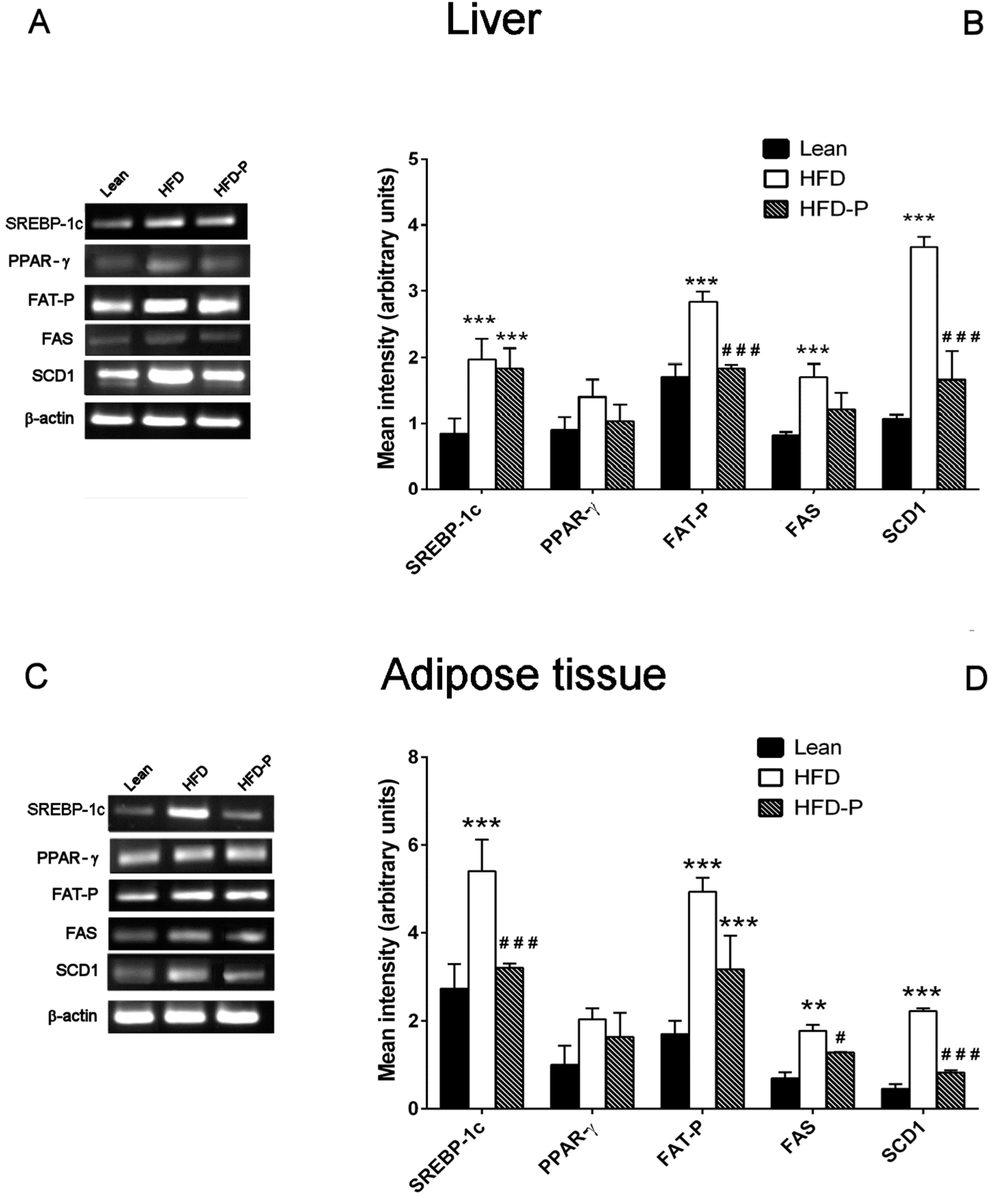
3.1.4. Effect of Pistachio Consumption on Lipid Metabolism-Related Genes Expression
3.2. Reversal Study
Pistachio Consumption Improves HFD-Induced Lipid Dysmetabolism
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Phillips, C.M. Nutrigenetics and Metabolic Disease: Current Status and implications for Personalised Nutrition. Nutrients 2013, 5, 32–57. [Google Scholar] [CrossRef] [PubMed]
- Suárez, M.; Boqué, N.; Del Bas, J.M.; Mayneris-Perxachs, J.; Arola, L.; Caimari, A. Mediterranean Diet and Multi-Ingredient-Based Interventions for the Management of Non-Alcoholic Fatty Liver Disease. Nutrients 2017, 9, 1052. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.H.; Zhang, C.; Zhang, D.G.; Li, L.; Chen, X.; Xu, D.X. Rifampicin-Induced Hepatic Lipid Accumulation: Association with Up-Regulation of Peroxisome Proliferator-Activated Receptor γ in Mouse Liver. PLoS ONE 2016, 11, e0165787. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.J.; Shan, C.W.; Li, B.; Pang, M.X.; Wang, H.; Luo, Y.; Liu, Y.L.; Song, Y.; Wang, N.N.; Chen, S.H.; et al. Linarin Enriched Extract Attenuates Liver Injury and Inflammation Induced by High-Fat High-Cholesterol Diet in Rats. Evid. Based Complement. Altern. Med. 2017, 2017, 4701570. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Park, H.S.; Hong, M.J.; Hur, H.J.; Kwon, D.Y.; Kim, M.S. Caffeic Acid Phenethyl Ester Improves Metabolic Syndrome by Activating PPAR-γ and Inducing Adipose Tissue Remodeling in Diet-Induced Obese Mice. Mol. Nutr. Food Res. 2018, 62, e1700701. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Han, J.M.; Lee, J.S.; Son, C.G.; Im, H.J.; Jo, H.K.; Yoo, H.R.; Kim, Y.S.; Seol, I.C. ACE Reduces Metabolic Abnormalities in a High-Fat Diet Mouse Model. Evid. Based Complement. Altern. Med. 2015, 2015, 352647. [Google Scholar] [CrossRef] [PubMed]
- Vadivel, V.; Kunyanga, C.N.; Biesalski, H.K. Health benefits of nut consumption with special reference to body weight control. Nutrition 2012, 28, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Ros, E. Health Benefits of Nut Consumption. Nutrients 2010, 2, 652–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzo, S.; Baldassano, S.; Caldara, G.F.; Ferrantelli, V.; Lo Dico, G.; Mulè, F.; Amato, A. Health benefits of pistachios consumption. Nat. Prod. Res. 2017, 15, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.Y.; Dhillon, J.; Mattes, R.D. A review of the effects of nuts on appetite, food intake, metabolism, and body weight. Am. J. Clin. Nutr. 2014, 100, 412S–422S. [Google Scholar] [CrossRef] [PubMed]
- Askari, G.; Yazdekhasti, N.; Mohammadifard, N.; Sarrafzadegan, N.; Bahonar, A.; Badiei, M.; Sajjadi, F.; Taheri, M. The relationship between nut consumption and lipid profile among the Iranian adult population; Isfahan Healthy Heart Program. Eur. J. Clin. Nutr. 2013, 67, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Del Gobbo, L.C.; Falk, M.C.; Feldman, R.; Lewis, K.; Mozaffarian, D. Effects of tree nuts on blood lipids, apolipoproteins, and blood pressure: Systematic review, meta-analysis, and dose-response of 61 controlled intervention trials. Am. J. Clin. Nutr. 2015, 102, 1347–1356. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Alonso, P.; Bulló, M.; Salas-Salvadó, J. Pistachios for Health: What Do We Know About This Multifaceted Nut? Nutr. Today 2016, 51, 133–138. [Google Scholar] [CrossRef] [PubMed]
- D’Evoli, L.; Lucarini, M.; Gabrielli, P.; Aguzzi, A.; Lombardi-Boccia, G. Nutritional value of italian pistachios from Bronte (Pistacia vera, L.) their nutrients, bioactive compounds and antioxidant activity. Food Nutr. Sci. 2015, 6. [Google Scholar] [CrossRef]
- Herbello-Hermelo, P.; Lamas, J.P.; Lores, M.; Domínguez-González, R.; Bermejo-Barrera, P.; Moreda-Piñeiro, A. Polyphenol bioavailability in nuts and seeds by an in vitro dialyzability approach. Food Chem. 2018, 254, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Gentile, C.; Allegra, M.; Angileri, F.; Pintaudi, A.M.; Livrea, M.A.; Tesoriere, L. Polymeric proanthocyanidins from Sicilian pistachio (Pistacia vera L.) nut extract inhibit lipopolysaccharide-induced inflammatory response in RAW 264.7 cells. Eur. J. Nutr. 2012, 51, 353–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaidya, H.B.; Gangadaran, S.; Cheema, S.K. A high fat-high sucrose diet enriched in blue mussels protects against systemic inflammation, metabolic dysregulation and weight gain in C57BL/6 mice. Food Res. Int. 2017, 100, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Vilahur, G.; Ben-Aicha, S.; Diaz, E.; Badimon, L.; Padro, T. Phytosterols and inflammation. Curr. Med. Chem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, Z.; Liu, Y.; Lv, X.; Yang, W. Effects of pistachios on body weight in Chinese subjects with metabolic syndrome. Nutr. J. 2012, 11, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebauer, S.K.; West, S.G.; Kay, C.D.; Alaupovic, P.; Bagshaw, D.; Kris-Etherton, P.M. Effects of pistachios on cardiovascular disease risk factors and potential mechanisms of action: A dose-response study. Am. J. Clin. Nutr. 2008, 88, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Aldemir, M.; Okulu, E.; Neşelioğlu, S.; Erel, O.; Kayıgil, O. Pistachio diet improves erectile function parameters and serum lipid profiles in patients with erectile dysfunction. Int. J. Impot. Res. 2011, 23, 32–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, K.; Kwaw, I.; Matud, J.; Kurtz, I. Effect of pistachio nuts on serum lipid levels in patients with moderate hypercholesterolemia. J. Am. Coll. Nutr. 1999, 18, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Kocyigit, A.; Koylu, A.A.; Keles, H. Effects of pistachio nuts consumption on plasma lipid profile and oxidative status in healthy volunteers. Nutr. Metab. Cardiovasc. Dis. 2006, 16, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, M.J.; Cooper, J.N.; Erario, M.; Cheifetz, C.E. Pistachio nut consumption and serum lipid levels. J. Am. Coll. Nutr. 2007, 26, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Alonso, P.; Salas-Salvadó, J.; Baldrich-Mora, M.; Juanola-Falgarona, M.; Bulló, M. Beneficial effect of pistachio consumption on glucose metabolism, insulin resistance, inflammation, and related metabolic risk markers: A randomized clinical trial. Diabetes Care 2014, 37, 3098–3105. [Google Scholar] [CrossRef] [PubMed]
- Baldassano, S.; Amato, A.; Cappello, F.; Rappa, F.; Mulè, F. Glucagon-like peptide-2 and mouse intestinal adaptation to a high-fat diet. J. Endocrinol. 2013, 217, 11–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldassano, S.; Rappa, F.; Amato, A.; Cappello, F.; Mulè, F. GLP-2 as Beneficial Factor in the Glucose Homeostasis in Mice Fed a High Fat Diet. J. Cell. Physiol. 2015, 230, 3029–3036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amato, A.; Caldara, G.F.; Nuzzo, D.; Baldassano, S.; Picone, P.; Rizzo, M.; Mulè, F.; Di Carlo, M. NAFLD and Atherosclerosis Are Prevented by a Natural Dietary Supplement Containing Curcumin, Silymarin, Guggul, Chlorogenic Acid and Inulin in Mice Fed a High-Fat Diet. Nutrients 2017, 13, 492. [Google Scholar] [CrossRef] [PubMed]
- Awień, J.; Nastałek, P.; Korbut, R. Mouse models of experimental atherosclerosis. J. Physiol. Pharmacol. 2004, 55, 503–517. [Google Scholar]
- Nuzzo, D.; Picone, P.; Baldassano, S.; Caruana, L.; Messina, E.; Marino Gammazza, A.; Cappello, F.; Mulè, F.; Di Carlo, M. Insulin resistance as common molecular denominator linking obesity to Alzheimer’s disease. Curr. Alzheimer Res. 2015, 12, 723–735. [Google Scholar] [CrossRef] [PubMed]
- Donato, A.J.; Henson, G.D.; Hart, C.R.; Layec, G.; Trinity, J.D.; Bramwell, R.C.; Enz, R.A.; Morgan, R.G.; Reihl, K.D.; Hazra, S.; et al. The impact of ageing on adipose structure, function and vasculature in the B6D2F1 mouse: Evidence of significant multisystem dysfunction. J. Physiol. 2014, 592, 4083–4096. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Menke, A.L.; Driessen, A.; Koek, G.H.; Lindeman, J.H.; Stoop, R.; Havekes, L.M.; Kleemann, R.; van den Hoek, A.M. Establishment of a general NAFLD scoring system for rodent models and comparison to human liver pathology. PLoS ONE 2014, 23, e115922. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, A.; Varedi, M.; Hadjzadeh, M.A.; Omrani, G.H. Type-1 diabetes induces depot-specific alterations in adipocyte diameter and mass of adipose tissues in the rat. Exp. Clin. Endocrinol. Diabetes 2010, 118, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Baldassano, S.; Amato, A.; Caldara, G.F.; Mulè, F. Glucagon-like peptide-2 treatment improves glucose dysmetabolism in mice fed a high-fat diet. Endocrine 2016, 54, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Baldassano, S.; Amato, A.; Rappa, F.; Cappello, F.; Mulè, F. Influence of endogenous glucagon-like peptide-2 on lipid disorders in mice fed a high-fat diet. Endocr. Res. 2016, 41, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Tokuşoglu, O.; Unal, M.K.; Yemiş, F. Determination of the phytoalexin resveratrol (3,5,4′-trihydroxystilbene) in peanuts and pistachios by high-performance liquid chromatographic diode array (HPLC-DAD) and gas chromatography-mass spectrometry (GC-MS). J. Agric. Food Chem. 2005, 53, 5003–5009. [Google Scholar] [CrossRef] [PubMed]
- Dreher, M.L. Pistachio nuts: Composition and potential health benefits. Nutr. Rev. 2012, 70, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Gentile, C.; Perrone, A.; Attanzio, A.; Tesoriere, L.; Livrea, M.A. Sicilian pistachio (Pistacia vera L.) nut inhibits expression and release of inflammatory mediators and reverts the increase of paracellular permeability in IL-1β-exposed human intestinal epithelial cells. Eur. J. Nutr. 2015, 54, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Paterniti, I.; Impellizzeri, D.; Cordaro, M.; Siracusa, R.; Bisignano, C.; Gugliandolo, E.; Carughi, A.; Esposito, E.; Mandalari, G.; Cuzzocrea, S. The Anti-Inflammatory and Antioxidant Potential of Pistachios (Pistacia vera L.) In Vitro and In Vivo. Nutrients 2017, 22, 915. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kris-Etherton, P.M.; Thompson, J.T.; Vanden Heuvel, J.P. Effect of pistachio oil on gene expression of IFN-induced protein with tetratricopeptide repeats 2: A biomarker of inflammatory response. Mol. Nutr Food Res. 2010, 54, S83–S92. [Google Scholar] [CrossRef] [PubMed]
- Alturfan, A.A.; Emekli-Alturfan, E.; Uslu, E. Consumption of pistachio nuts beneficially affected blood lipids and total antioxidant activity in rats fed a high-cholesterol diet. Folia Biol. (Praha) 2009, 55, 132–136. [Google Scholar] [PubMed]
- Sari, I.; Baltaci, Y.; Bagci, C.; Davutoglu, V.; Erel, O.; Celik, H.; Ozer, O.; Aksoy, N.; Aksoy, M. Effect of pistachio diet on lipid parameters, endothelial function, inflammation, and oxidative status: A prospective study. Nutrition 2010, 26, 399–404. [Google Scholar] [CrossRef] [PubMed]
- West, S.G.; Gebauer, S.K.; Kay, C.D.; Bagshaw, D.M.; Savastano, D.M.; Diefenbach, C.; Kris-Etherton, P.M. Diets containing pistachios reduce systolic blood pressure and peripheral vascular responses to stress in adults with dyslipidemia. Hypertension 2012, 60, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Kendall, C.W.; West, S.G.; Augustin, L.S.; Esfahani, A.; Vidgen, E.; Bashyam, B.; Sauder, K.A.; Campbell, J.; Chiavaroli, L.; Jenkins, A.L.; et al. Acute effects of pistachio consumption on glucose and insulin, satiety hormones and endothelial function in the metabolic syndrome. Eur. J. Clin. Nutr. 2014, 68, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Gulati, S.; Misra, A.; Pandey, R.M.; Bhatt, S.P.; Saluja, S. Effects of pistachio nuts on body composition, metabolic, inflammatory and oxidative stress parameters in Asian Indians with metabolic syndrome: A 24-wk, randomized control trial. Nutrition 2014, 30, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Aksoy, N.; Aksoy, M.; Bagci, C.; Gergerlioglu, H.S.; Celik, H.; Herken, E.; Yaman, A.; Tarakcioglu, M.; Soydinc, S.; Sari, I.; et al. Pistachio intake increases high density lipoprotein levels and inhibits low-density lipoprotein oxidation in rats. Tohoku J. Exp. Med. 2007, 212, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Holligan, S.D.; West, S.G.; Gebauer, S.K.; Kay, C.D.; Kris-Etherton, P.M. A moderate-fat diet containing pistachios improves emerging markers of cardiometabolic syndrome in healthy adults with elevated LDL levels. Br. J. Nutr. 2014, 112, 744–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Alonso, P.; Salas-Salvadó, J.; Baldrich-Mora, M.; Mallol, R.; Correig, X.; Bulló, M. Effect of pistachio consumption on plasma lipoprotein subclasses in pre-diabetic subjects. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Silva Figueiredo, P.; Carla Inada, A.; Marcelino, G.; Maiara Lopes Cardozo, C.; de Cássia Freitas, K.; de Cássia Avellaneda Guimarães, R.; Pereira de Castro, A.; Aragão do Nascimento, V.; Aiko Hiane, P. Fatty Acids Consumption: The Role Metabolic Aspects Involved in Obesity and Its Associated Disorders. Nutrients 2017, 9, 1158. [Google Scholar] [CrossRef] [PubMed]
- Altmann, S.W.; Davis, H.R., Jr.; Zhu, L.J.; Yao, X.; Hoos, L.M.; Tetzloff, G.; Iyer, S.P.; Maguire, M.; Golovko, A.; Zeng, M.; et al. Niemann-Pick C1 Like 1 protein is critical for intestinal cholesterol absorption. Science 2004, 303, 1201–1204. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Mah, X.J.; Garcia, M.C.; Antonypillai, C.; van der Poorten, D. Oily fish, coffee and walnuts: Dietary treatment for nonalcoholic fatty liver disease. World J. Gastroenterol. 2015, 21, 10621–10635. [Google Scholar] [CrossRef] [PubMed]
- Han, J.M.; Jo, A.N.; Lee, S.M.; Bae, H.S.; Jun, D.W.; Cho, Y.K.; Suk, K.T.; Yoon, J.H.; Ahn, S.B.; Cho, Y.J.; et al. Associations between intakes of individual nutrients or whole food groups and non-alcoholic fatty liver disease among Korean adults. J. Gastroenterol. Hepatol. 2014, 29, 1265–1272. [Google Scholar] [CrossRef] [PubMed]
- Jump, D.B. Fatty acid regulation of hepatic lipid metabolism. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, P.; Leray, V.; Diez, M.; Serisier, S.; Le Bloc’h, J.; Siliart, B.; Dumon, H. Liver lipid metabolism. J. Anim. Physiol. Anim. Nutr. 2008, 92, 272–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Z.; Pei, Z.; Maiguel, D.; Toomer, C.J.; Watkins, P.A. The fatty acid transport protein (FATP) family: Very long chain acyl-CoA synthetases or solute carriers? J. Mol. Neurosci. 2007, 33, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.F.; Le, G.W.; Wang, P.; Qiu, Y.Y.; Jiang, Y.Y.; Tang, X. Regressive Effect of Myricetin on Hepatic Steatosis in Mice Fed a High-Fat Diet. Nutrients 2016, 8, 799. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.S.; Liao, S.N.; Tsai, M.L.; Kalyanam, N.; Majeed, M.; Majeed, A.; Ho, C.T.; Pan, M.H. Calebin-A inhibits adipogenesis and hepatic steatosis in high-fat diet-induced obesity via activation of AMPK signaling. Mol. Nutr. Food Res. 2015, 59, 1883–1895. [Google Scholar] [CrossRef] [PubMed]
- Oliveira Andrade, J.M.; Paraíso, A.F.; Garcia, Z.M.; Ferreira, A.V.; Sinisterra, R.D.; Sousa, F.B.; Guimarães, A.L.; de Paula, A.M.; Campagnole-Santos, M.J.; dos Santos, R.A.; Santos, S.H. Cross talk between angiotensin-(1-7)/Mas axis and sirtuins in adipose tissue and metabolism of high-fat feed mice. Peptides 2014, 55, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.; Dela Cruz, J.F.; Kim, W.S.; Yoo, K.; Hwang, S.G. Glehnia littoralis Root Extract Inhibits Fat Accumulation in 3T3-L1 Cells and High-Fat Diet-Induced Obese Mice by Downregulating Adipogenic Gene Expression. Evid. Based Complement. Altern. Med. 2018, 2018, 1243049. [Google Scholar] [CrossRef] [PubMed]
- Ayoub, H.M.; McDonald, M.R.; Sullivan, J.A.; Tsao, R.; Meckling, K.A. Proteomic Profiles of Adipose and Liver Tissues from an Animal Model of Metabolic Syndrome Fed Purple Vegetables. Nutrients 2018, 10, 456. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Cantú, L.N.; Gutiérrez-Uribe, J.A.; Arriola-Vucovich, J.; Díaz-De La Garza, R.I.; Fahey, J.W.; Serna-Saldivar, S.O. Broccoli (Brassica oleracea var. italica) sprouts and extracts rich in glucosinolates and isothiocyanates affect cholesterol metabolism and genes involved in lipid homeostasis in hamsters. J. Agric. Food Chem. 2011, 59, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Abraham, T.M.; Pedley, A.; Massaro, J.M.; Hoffmann, U.; Fox, C.S. Association between visceral and subcutaneous adipose depots and incident cardiovascular disease risk factors. Circulation 2015, 132, 1639–1647. [Google Scholar] [CrossRef] [PubMed]
- Bays, H. Central obesity as a clinical marker of adiposopathy; increased visceral adiposity as a surrogate marker for global fat dysfunction. Curr. Opin. Endocrinol. Diabetes Obes. 2014, 21, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Pandžić Jakšić, V.; Grizelj, D. Under the surface of subcutaneous adipose tissue biology. Acta Dermatovenerol. Croat. 2016, 24, 250–260. [Google Scholar] [PubMed]
- Smith, U.; Kahn, B.B. Adipose tissue regulates insulin sensitivity: Role of adipogenesis, de novo lipogenesis and novel lipids. J. Intern. Med. 2016, 280, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.V.; Murthy, V.L.; Allison, M.A.; Ding, J.; Budoff, M.; Frazier-Wood, A.C.; Lima, J.A.; Steffen, L.; Siscovick, D.; Tucker, K.L.; et al. Diet and adipose tissue distributions: The Multi-Ethnic Study of Atherosclerosis. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (g/kg) | STD | HFD | HFD-P |
|---|---|---|---|
| Acid Casein 741 | 200 | 265.00 | 210.00 |
| L-Cystine | 2.8 | 4 | 4 |
| Maltodextrine-0032 | 33.2 | 160 | 125.5 |
| Sucrose | 300 | 90 | 100 |
| Cellulose (Arbocel) | 50 | 65.5 | 50 |
| Soybean Oil | 25 | 30 | 30 |
| Lard | 19 | 220 | 135 |
| Vitamin mix AIN-93-VX-PF2439 | 10 | 21 | 21 |
| Mineral mix AIN-93G-MX-PF2348 | 45 | 48 | 48 |
| Choline Bitartrate | 1.9 | 3 | 3 |
| Calcium Phosphate Dibasic | 13 | 3.4 | 3.4 |
| Pistachio | - | - | 180 |
| Total Energy, Kcal/g | 3.5 | 6 | 6 |
| Protein, % | 20 | 20 | 20 |
| Carbohydrate, % | 70 | 20 | 20 |
| Fat, % | 10 | 60 | 60 |
| Fatty Acids (g/kg) | HFD | HFD-P |
|---|---|---|
| total saturated | 92.5 | 71.12 |
| total monounsaturated | 117.5 | 136.65 |
| total polyunsaturated | 40 | 42.23 |
| Gene | Forward Primer | Reverse Primer | Size (bp) |
|---|---|---|---|
| FAS | 5′-TACTTTGTGGCCTTCTCCTCTGTAA-3′ | 5′-CTTCCACACCCATGAGCGAGTCCAGGCCGA-3′ | 445 |
| SCD1 | 5′-GCCAGACCGGGCTGAACACC-3′ | 5′-GGCCTCCCAAGTGCAGCAGG-3′ | 397 |
| SREBP-1c | 5′-GGAGACATCGCAAACAAGC-3′ | 5′-GGTAGACAACAGCCGCATC-3′ | 273 |
| PPAR-ɣ | 5′-GGGCTGAGGAGAAGTCACAC-3′ | 5′-TCAGTGGTTCACCGCTTCTT-3′ | 142 |
| FAT-P | 5′-CGCCGATGTGCTCTATGACT-3′ | 5′-ACACAGTCATCCCAGAAGCG-3′ | 138 |
| β-actin | 5′-GGATCCCCGCCCTAGGCACCAGGGT-3′ | 5′-GGAATTCGGCTGGGGTGTTGAAGGTCTCAAA-3′ | 289 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terzo, S.; Caldara, G.F.; Ferrantelli, V.; Puleio, R.; Cassata, G.; Mulè, F.; Amato, A. Pistachio Consumption Prevents and Improves Lipid Dysmetabolism by Reducing the Lipid Metabolizing Gene Expression in Diet-Induced Obese Mice. Nutrients 2018, 10, 1857. https://doi.org/10.3390/nu10121857
Terzo S, Caldara GF, Ferrantelli V, Puleio R, Cassata G, Mulè F, Amato A. Pistachio Consumption Prevents and Improves Lipid Dysmetabolism by Reducing the Lipid Metabolizing Gene Expression in Diet-Induced Obese Mice. Nutrients. 2018; 10(12):1857. https://doi.org/10.3390/nu10121857
Chicago/Turabian StyleTerzo, Simona, Gaetano Felice Caldara, Vincenzo Ferrantelli, Roberto Puleio, Giovanni Cassata, Flavia Mulè, and Antonella Amato. 2018. "Pistachio Consumption Prevents and Improves Lipid Dysmetabolism by Reducing the Lipid Metabolizing Gene Expression in Diet-Induced Obese Mice" Nutrients 10, no. 12: 1857. https://doi.org/10.3390/nu10121857