Estimation of Alpine Grassland Forage Nitrogen Coupled with Hyperspectral Characteristics during Different Growth Periods on the Tibetan Plateau

 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Grassland Observation Data
2.3. Spectral Variables
2.4. Data Analysis and Modeling
2.4.1. RF Algorithm
2.4.2. Variable Selection
2.4.3. Validation Strategies
3. Results
3.1. Variations in Forage N Contents and Reflectance Spectra during Different Growth Periods
3.2. Spectral Absorption Features and Red-Edge Shift
3.3. Relationship between the Forage N Content and Different Variables
3.4. Estimation Model for Forage N during Different Growth Periods
3.5. Estimation Model for Forage N throughout the Growth Periods
4. Discussion
4.1. Variation in Forage N Contents during Different Growth Periods
4.2. Spectral Characteristics of the Forage Canopy during Different Growth Periods
4.3. Applicability of Spectral Variables for Estimating Forage N during Different Growth Periods
4.4. Target-Oriented Validation
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- He, J.S.; Wang, L.; Flynn, D.F.B.; Wang, X.P.; Ma, W.H.; Fang, J.Y. Leaf nitrogen: Phosphorus stoichiometry across Chinese grassland biomes. Oecologia 2018, 155, 301–310. [Google Scholar] [CrossRef]
- Lu, X.; Yan, Y.; Sun, J.; Zhang, X.K.; Chen, Y.C.; Wang, X.D.; Cheng, G.W. Carbon, nitrogen, and phosphorus storage in alpine grassland ecosystems of Tibet: Effects of grazing exclusion. Ecol. Evol. 2015, 5, 4492–4504. [Google Scholar] [CrossRef]
- Adams, M.A.; Turnbull, T.L.; Sprent, J.I.; Buchmann, N. Legumes are different: Leaf nitrogen, photosynthesis, and water use efficiency. Proc. Natl. Acad. Sci. USA 2016, 113, 4098–4103. [Google Scholar] [CrossRef] [Green Version]
- Heckathorn, S.A.; Delucia, E.H.; Zielinski, R.E. The contribution of drought-related decreases in foliar nitrogen concentration to decreases in photosynthetic capacity during and after drought in prairie grasses. Physiol. Plant. 1997, 101, 173–182. [Google Scholar] [CrossRef]
- Lu, J.L.; Hu, A.T. Plant Nutriology; Higher Education Press: Beijing, China, 2006. [Google Scholar]
- Reich, P.B.; Walters, M.B.; Tjoelker, M.G.; Vanderklein, D.; Buschena, C. Photosynthesis and respiration rates depend on leaf and root morphology and nitrogen concentration in nine boreal tree species differing in relative growth rate. Funct. Ecol. 2010, 12, 395–405. [Google Scholar] [CrossRef]
- Beeri, O.; Phillips, R.; Hendrickson, J.; Frank, A.B.; Kronberg, S. Estimating forage quantity and quality using aerial hyperspectral imagery for northern mixed-grass prairie. Remote Sens. Environ. 2007, 110, 216–225. [Google Scholar] [CrossRef]
- Mutanga, O.; Adam, E.; Adjorlolo, C.; Rahman, E.M.A. Evaluating the robustness of models developed from field spectral data in predicting African grass foliar nitrogen concentration using WorldView-2 image as an independent test dataset. Int. J. Appl. Earth Obs. Geoinf. 2015, 34, 178–187. [Google Scholar] [CrossRef]
- Mutanga, O.; Skidmore, A.K.; Prins, H.H.T. Predicting in situ pasture quality in the Kruger National Park, South Africa, using continuum-removed absorption features. Remote Sens. Environ. 2004, 89, 393–408. [Google Scholar] [CrossRef]
- Singh, L.; Mutanga, O. Remote sensing of key grassland nutrients using hyperspectral techniques in KwaZulu-Natal, South Africa. J. Appl. Remote Sens. 2017, 11, 036005. [Google Scholar] [CrossRef]
- Shoko, C.; Mutanga, O.; Dube, T. Progress in the remote sensing of C3 and C4 grass species aboveground biomass over time and space. ISPRS J. Photogramm. Remote Sens. 2016, 120, 13–24. [Google Scholar] [CrossRef]
- Adjorlolo, C.; Cho, M.A.; Mutanga, O.; Ismail, R. Optimizing spectral resolutions for the classification of C3 and C4 grass species, using wavelengths of known absorption features. J. Appl. Remote Sens. 2012, 6, 217–224. [Google Scholar] [CrossRef]
- Mccann, C.; Repasky, K.S.; Lawrence, R.; Powell, S. Multi-temporal mesoscale hyperspectral data of mixed agricultural and grassland regions for anomaly detection. ISPRS J. Photogramm. Remote Sens. 2017, 131, 121–133. [Google Scholar] [CrossRef]
- Ramoelo, A.; Skidmore, A.K.; Cho, M.A.; Mathieu, R.; Heitkönig, I.M.A.; Dudeni-Tlhone, N.; Schlerf, M.; Prins, H.H.T. Non-linear partial least square regression increases the estimation accuracy of grass nitrogen and phosphorus using in situ hyperspectral and environmental data. ISPRS J. Photogramm. Remote Sens. 2013, 82, 27–40. [Google Scholar] [CrossRef]
- Ramoelo, A.A. Monitoring grass nutrients and biomass as indicators of rangeland quality and quantity using random forest modelling and WorldView-2 data. Int. J. Appl. Earth Obs. Geoinf. 2015, 43, 43–54. [Google Scholar] [CrossRef]
- Curran, P.J. Remote sensing of foliar chemistry. Remote Sens. Environ. 1989, 30, 271–278. [Google Scholar] [CrossRef]
- Knox, N. Observing Temporal and Spatial Variability of Forage Quality. Ph.D. Thesis, Faculty Geo-Information Science and Earth Observation and Twente University, Enschede, The Netherlands, 2010. [Google Scholar]
- Schlerf, M.; Atzberger, C.; Hill, J.; Buddenbaum, H.; Werner, W.; Schüler, G. Retrieval of chlorophyll and nitrogen in Norway spruce (Picea abies L. Karst.) using imaging spectroscopy. Int. J. Appl. Earth Obs. Geoinf. 2010, 12, 17–26. [Google Scholar] [CrossRef]
- Skidmore, A.K.; Ferwerda, J.G.; Mutanga, O.; Van Wieren, S.E.; Peel, M.; Grant, R.C.; Prins, H.H.T.; Balcik, F.B.; Venus, V. Forage quality of savannas–simultaneously mapping foliar protein and polyphenols for trees and grass using hyperspectral imagery. Remote Sens. Environ. 2010, 114, 64–72. [Google Scholar] [CrossRef]
- Mutanga, O.; Skidmore, A.K. Red edge shift and biochemical content in grass canopies. ISPRS J. Photogramm. Remote Sens. 2007, 62, 34–42. [Google Scholar] [CrossRef]
- Clevers, J.G.P.W.; Gitelson, A.A. Remote estimation of crop and grass chlorophyll and nitrogen content using red-edge bands on Sentinel-2 and -3. Int. J. Appl. Earth Obs. Geoinf. 2013, 23, 344–351. [Google Scholar] [CrossRef]
- Lin, D.; Wei, G.; Jian, Y. Application of spectral indices and reflectance spectrum on leaf nitrogen content analysis derived from hyperspectral LiDAR data. Opt. Laser Technol. 2018, 107, 372–379. [Google Scholar]
- Choubey, V.K.; Choubey, R. Spectral reflectance, growth and chlorophyll relationships for rice crop in a semi-arid region of India. Water Resour. Manag. 1999, 13, 73–84. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Ramoelo, A.; Cho, M.A.; Madonsela, S.; Mathieu, R.; Korchove, R.V.D.; Kaszta, Z.; Wolf, E. A potential to monitor nutrients as an indicator of rangeland quality using space borne remote sensing. IOP Conf. Ser. Earth Environ. Sci. 2014, 18, 012094. [Google Scholar] [CrossRef] [Green Version]
- Gong, Z.; Kawamura, K.; Ishikawa, N.; Inaba, M.; Alateng, D. Estimation of herbage biomass and nutritive status using band depth features with partial least squares regression in Inner Mongolia grassland, China. Grassl. Sci. 2016, 62, 45–54. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Mutanga, O.; Adam, E.; Cho, M.A. High density biomass estimation for wetland vegetation using WorldView-2 imagery and random forest regression algorithm. Int. J. Appl. Earth Obs. Geoinf. 2012, 18, 399–406. [Google Scholar] [CrossRef]
- Singh, L.; Mutanga, O.; Mafongoya, P.; Peerbhay, K.Y. Multispectral mapping of key grassland nutrients in KwaZulu-Natal, South Africa. J. Spat. Sci. 2018, 63, 155–172. [Google Scholar] [CrossRef]
- Gao, J.L.; Meng, B.P.; Liang, T.G.; Feng, Q.S.; Ge, J.; Yin, J.P.; Wu, C.X.; Cui, X.; Hou, M.J.; Liu, J.; et al. Modeling alpine grassland forage phosphorus based on hyperspectral remote sensing and a multi-factor machine learning algorithm in the east of Tibetan Plateau, China. ISPRS J. Photogramm. Remote Sens. 2019, 147, 104–117. [Google Scholar] [CrossRef]
- Mutanga, O. Hyperspectral Remote Sensing of Tropical Grass Quality and Quantity. Ph.D. Thesis, International Institute for Geoinformation Science and Earth Observation and Wageningen University, Wageningen, The Netherlands, 2004. [Google Scholar]
- Horler, D.N.H.; Dockray, M.; Barber, J. The red edge of plant leaf reflectance. Int. J. Remote Sens. 1983, 4, 273–288. [Google Scholar] [CrossRef]
- Guo, C.F.; Guo, X.Y. Estimating leaf chlorophyll and nitrogen content of wetland emergent plants using hyperspectral data in the visible domain. Spectrosc. Lett. 2016, 49, 180–187. [Google Scholar] [CrossRef]
- Jordan, C.F. Derivation of leaf area index from quality of light on the forest floor. Ecology 1969, 50, 663–666. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N. Spectral reflectance changes associated with autumn senescence of Aesculus hippocastanum L. and Acer platanoides L. leaves. Spectral Features and Relation to Chlorophyll Estimation. J. Plant Physiol. 1994, 143, 286–292. [Google Scholar] [CrossRef]
- Datt, B. A new reflectance index for remote sensing of chlorophyll content in higher plants: Tests using Eucalyptus leaves. J. Plant Physiol. 1991, 154, 30–36. [Google Scholar] [CrossRef]
- Clevers, J.G.P.W. Imaging Spectrometry in Agriculture—Plant Vitality and Yield Indicators Imaging Spectrometry—A Tool for Environmental Observations; Springer: Dordrecht, The Netherlands, 1994; pp. 193–219. [Google Scholar]
- Vogelmann, J.E.; Rock, B.N.; Moss, D.M. Red edge spectral measurements from sugar maple leaves. Int. J. Remote Sens. 1993, 14, 1563–1575. [Google Scholar] [CrossRef]
- Fourty, T.; Baret, F.; Jacquemoud, S. Leaf optical properties with explicit description of its biochemical composition: Direct and inverse problems. Remote Sens. Environ. 1996, 56, 104–117. [Google Scholar] [CrossRef]
- Serrano, L.; Peñuelas, J.; Ustin, S.L. Remote sensing of nitrogen and lignin in Mediterranean vegetation from AVIRIS data: Decomposing biochemical from structural signals. Remote Sens. Environ. 2002, 81, 355–364. [Google Scholar] [CrossRef]
- Gamon, J.A.; Penuelas, J.; Field, C.B. A narrow-waveband spectral index that tracks diurnal changes in photosynthetic efficiency. Remote Sens. Environ. 1992, 41, 35–44. [Google Scholar] [CrossRef]
- Penuelas, J.; Baret, F.; Filella, I. Semiempirical indexes to assess carotenoids chlorophyll-a ratio from leaf spectral reflectance. Photosynthetica 1995, 31, 221–230. [Google Scholar]
- Rondeaux, G.; Steven, M.; Baret, F. Optimization of soil-adjusted vegetation indices. Remote Sens. Environ. 1996, 55, 95–107. [Google Scholar] [CrossRef]
- Richardson, A.J.; Wiegand, C.L. Distinguishing vegetation from soil background information (by gray mapping of Landsat MSS data). Photogramm. Eng. Remote Sens. 1997, 43, 1541–1552. [Google Scholar]
- Gitelson, A.A.; Merzlyak, M.N. Signature analysis of leaf reflectance spectra: Algorithm development for remote sensing of chlorophyll. J. Plant Physiol. 1996, 148, 494–500. [Google Scholar] [CrossRef]
- Marshak, A.; Knyazikhin, Y.; Davis, A.; Wiscombe, W.; Pilewskie, P. Cloud-vegetation interaction: Use of normalized difference cloud index for estimation of cloud optical thickness. Geophys. Res. Lett. 2000, 27, 1695–1698. [Google Scholar] [CrossRef] [Green Version]
- Huete, A.R. A soil-adjusted vegetation index (SAVI). Remote Sens. Environ. 1988, 25, 295–309. [Google Scholar] [CrossRef]
- Roujean, J.L.; Breon, F.M. Estimating PAR absorbed by vegetation from bidirectional reflectance measurements. Remote Sens. Environ. 1995, 51, 375–384. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Schleicher, T.D.; Bausch, W.C.; Delgado, J.A.; Ayers, P.D. Evaluation and Refinement of the Nitrogen Reflectance Index (NRI) for Site-Specific Fertilizer Management. In ASAE Annual International Meeting Report; American Society of Agricultural and Biological Engineers: St Joseph, MI, USA, 1998. [Google Scholar]
- Tian, Y.C.; Gu, K.J.; Chu, X.; Yao, X.; Cao, W.X.; Zhu, Y. Comparison of different hyperspectral vegetation indices for canopy leaf nitrogen concentration estimation in rice. Plant Soil 2014, 376, 193–209. [Google Scholar] [CrossRef]
- Kumar, L.; Schmidt, K.S.; Dury, S.; Skidmore, A.K. Imaging spectroscopy and vegetation science. Imaging Spectrom. Basic Princ. Prospect. Appl. 2001, 4, 111–155. [Google Scholar]
- Mutanga, O.; Skidmore, A.K.; Kumar, L.; Ferwerda, J. Estimating tropical pasture quality at canopy level using band depth analysis with continuum removal in the visible domain. Int. J. Remote Sens. 2005, 26, 1093–1108. [Google Scholar] [CrossRef]
- Han, Z.Y.; Zhu, X.C.; Fang, X.Y.; Wang, Z.Y.; Wang, L.; Zhao, G.X.; Jiang, Y.M. Hyperspectral estimation of apple tree canopy LAI based on SVM and RF regression. Spectrosc. Spectr. Anal. 2016, 36, 800–805. [Google Scholar]
- Lebedev, A.; Westman, E.; Van Westen, G.; Kramberger, M.; Lundervold, A.; Aarsland, D.; Soininen, H.; Kłoszewska, I.; Mecocci, P.; Tsolaki, M. Random Forest ensembles for detection and prediction of Alzheimer’s disease with a good between-cohort robustness. Neuroimage Clin. 2014, 6, 115–125. [Google Scholar] [CrossRef]
- Meyer, H.; Reudenbach, C.; Hengl, T.; Katurji, M.; Nauss, T. Improving performance of spatio-temporal machine learning models using forward feature selection and target-oriented validation. Environ. Model. Softw. 2018, 101, 1–9. [Google Scholar] [CrossRef]
- Efron, B.; Gong, G. A leisurely look at the bootstrap, the jackknife, and cross-validation. Am. Stat. 1983, 37, 36–48. [Google Scholar]
- Zhao, Z.; Wang, A.L.; Ma, H.S.; Song, H.Q. Studies on dynamics monitor and sustainable development in eastern edge of Qinghai-Tibetan alpine grassland (III Seasonal variational dynamics of nutrional contents of the predominant plants on 8 main grassland types). Pratacult. Sci. 2002, 19, 5–9. [Google Scholar]
- Liang, J.Y.; Jiao, T.; Wu, J.P.; Gong, X.Y.; Du, W.H.; Liu, H.B.; Xiao, Y.M. The relationship between seasonal forage digestibility and forage nutritive value in different grazing pastures. Acta Pratacult. Sin. 2015, 24, 108–115. [Google Scholar]
- Zhou, H.J.; Mao, Z.X.; Huang, D.J.; Fu, H. Analysis on nutritional quality of Elymus nutans among different populations on Qinghai-Tibet Plateau. Pratacult. Sci. 2011, 28, 1198–1202. [Google Scholar]
- Niklas, K.J. Plant allometry, leaf nitrogen and phosphorus stoichiometry, and interspecific trends in annual growth rates. Ann. Bot. 2005, 97, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Niklas, K.J.; Owens, T.; Reich, P.B.; Cobb, E.D. Nitrogen/phosphorus leaf stoichiometry and the scaling of plant growth. Ecol. Lett. 2005, 8, 636–642. [Google Scholar] [CrossRef]
- Shi, Y.; Ma, Y.L.; Ma, W.H.; Liang, C.Z.; Zhao, X.Q.; Fang, J.Y.; He, J.S. Large scale patterns of forage yield and quality across Chinese grasslands. Chin. Sci. Bull. 2013, 58, 1187–1199. [Google Scholar] [CrossRef]
- Huang, J.Y.; Zhu, X.G.; Yuan, Z.Y.; Song, S.H.; Li, X.; Li, L.H. Changes in nitrogen resorption traits of six temperate grassland species along a multi-level N addition gradient. Plant Soil 2008, 306, 149–158. [Google Scholar] [CrossRef]
- Li, L.; Gao, X.P.; Li, X.Y.; Lin, L.S.; Zeng, F.J.; Gui, D.W.; Lu, Y. Nitrogen (N) and phosphorus (P) resorption of two dominant alpine perennial grass species in response to contrasting N and P availability. Environ. Exp. Bot. 2016, 127, 37–44. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Gritz, Y.; Merzlyak, M.N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for non-destructive chlorophyll assessment in higher plant leaves. J. Plant Physiol. 2003, 160, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Feng, W. Monitoring Nitrogen Status and Growth Characters with Hyperspectral Remote Sensing in Wheat. Ph.D. Thesis, Nanjing Agriculture University, Nanjing, China, 2007. [Google Scholar]
- Yoder, B.J.; Pettigrew-Crosby, R.E. Predicting nitrogen and chlorophyll content and concentrations from reflectance spectra (400–2500 nm) at leaf and canopy scales. Remote Sens. Environ. 1995, 53, 199–211. [Google Scholar] [CrossRef]
- Filella, I.; Penuelas, J. The red edge position and shape as indicators of plant chlorophyll content, biomass and hydric status. Int. J. Remote Sens. 1994, 15, 1459–1470. [Google Scholar] [CrossRef]
- Sibanda, M.; Mutanga, O.; Rouget, M. Testing the capabilities of the new WorldView-3 space-borne sensor’s red-edge spectral band in discriminating and mapping complex grassland management treatments. Int. J. Remote Sens. 2017, 38, 1–22. [Google Scholar] [CrossRef]
- Zhang, F.L.; Yin, Q.; Kuang, D.B.; Li, F.X.; Zhou, B.R. Analysis of time series spectrum feature parameters derived from dominant natural grasslands in the region around Qinghai lake. Acta Ecol. Sin. 2005, 25, 3155–3160. [Google Scholar]
- Zhang, F.L.; Yin, Q.; Kuang, D.B.; Li, F.X.; Zhou, B.R. Analysis of a spectral experiment conducted through the growth period of the main grassland types found in the region around Qinghai Lake. Acta Pratacult. Sin. 2006, 15, 42–47. [Google Scholar]
- Janitza, S.; Hornung, R. On the overestimation of random forest’s out-of-bag error. PLoS ONE 2018, 13, e0201904. [Google Scholar] [CrossRef]
- Penuelas, J.; Gamon, J.A.; Freeden, A.L.; Merino, J.; Field, C. Reflectance indices associated with physiological changes in nitrogen and water-limited sunflower leaves. Remote Sens. Environ. 1994, 48, 135–146. [Google Scholar] [CrossRef]
- Jago, R.A.; Cutler, M.E.J.; Curran, P.J. Estimating canopy chlorophyll concentration from field and airborne spectra. Remote Sens. Environ. 1999, 68, 217–224. [Google Scholar] [CrossRef]
- Sellers, P.J. Canopy reflectance, photosynthesis and transpiration. Int. J. Remote Sens. 1985, 6, 1335–1372. [Google Scholar] [CrossRef]
- Thenkabail, P.S.; Smith, R.B.; De Pauw, E. Hyperspectral vegetation indices and their relationships with agricultural crop characteristics. Remote Sens. Environ. 2000, 71, 158–182. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Prueger, J.H. Value of using different vegetative indices to quantify agricultural crop characteristics at different growth stages under varying management practices. Remote Sens. 2010, 2, 562–578. [Google Scholar] [CrossRef]
- Ke, Y.; Im, J.; Park, S.; Gong, H. Downscaling of MODIS one kilometer evapotranspiration using Landsat-8 data and machine learning approaches. Remote Sens. 2016, 8, 215. [Google Scholar] [CrossRef]
- Ludwig, A.; Meyer, H.; Nauss, T. Automatic classification of Google Earth images for a larger scale monitoring of bush encroachment in South Africa. Int. J. Appl. Earth Obs. Geoinf. 2016, 50, 89–94. [Google Scholar] [CrossRef]
- Gudmundsson, L.; Seneviratne, S.I. Towards observation-based gridded runoff estimates for Europe. Hydrol. Earth Syst. Sci. 2015, 19, 2859–2879. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable Type | Variables | Formula and Description | References |
|---|---|---|---|
| Vegetation indexes (VIs) | Simple ratio index (SR) | [34] | |
| Red-edge normalized difference vegetation index (NDVI705) | [35] | ||
| Modified red-edge normalized difference vegetation index (mNDVI705) | [24,36] | ||
| Modified red-edge simple ratio index (mSR705) | [24,36] | ||
| Red-edge inflection point (REIP) | [37] | ||
| Vogelmann red-edge index 1 (VOG1) | [38] | ||
| Vogelmann red-edge index 2 (VOG2) | [38] | ||
| Vogelmann red-edge index 3 (VOG3) | [38] | ||
| Normalized difference nitrogen index (NDNI) | [39,40] | ||
| Photochemical reflectance index (PRI) | [41] | ||
| Structure insensitive pigment index (SIPI) | . | [42] | |
| Optimized soil-adjusted vegetation index (OSAVI) | [43] | ||
| Difference vegetation index (DVI) | [44] | ||
| Normalized difference greenness index (NDGI) | . | [45] | |
| Normalized difference cloud index (NDCI) | [46] | ||
| Soil-adjusted vegetation index (SAVI) | , = 0.5 | [47] | |
| Renormalized difference vegetation index (RDVI) | [48] | ||
| Normalized difference vegetation index 1 (NDVI 1) | [49] | ||
| Nitrogen reflectance index (NRI) | [50] | ||
| Three-band spectral index (TBSI) | [51] | ||
| Absorption bands | The spectral reflectance at λ nm (Rλ) | Λ = 430, 460, 640, 660, 910, 1510, 1940, 2060, 2180, 2300 | [16,52] |
| Red-edge parameters | Red-edge position (REP) AMP | Wavelength of the red-edge peak (maximum slope position) | [31] |
| Amplitude (AMP) | First derivative value at the red-edge peak (maximum slope) | ||
| Slope725 | First derivative value at 725 nm | ||
| Slope_mean | First derivative value obtained from the corresponding mean red-edge position | ||
| Absorption features | Absorption position (AP) | Absorption position | [53] |
| Absorption depth (AD) | AD = 1 − R′, R′ = continuum-removed spectra reflectance value | ||
| Band depth ratio (BDR) | BDR = BD/BDc, BDc = band depth (BD) of band centre, and BD = AD | ||
| Normalized band depth index (NBDI) | NBDI = (BD − BDc)/(BD + BDc) |
| Sample Areas | GP170624 | GP170727 | GP170830 | GP170927 | GP171115 |
|---|---|---|---|---|---|
| AZ | 2.13 ± 0.14 a | 1.70 ± 0.16 b | 1.58 ± 0.15 c | 1.10 ± 0.12 d | 0.92 ± 0.08 e |
| XC | 1.99 ± 0.29 a | 1.84 ± 0.26 a | 1.60 ± 0.23 ab | 1.29 ± 0.27 b | 0.92 ± 0.26 c |
| YLJ | 1.86 ± 0.20 a | 1.50 ± 0.08 b | 1.46 ± 0.18 b | 1.29 ± 0.20 b | 0.77 ± 0.08 c |
| GJ | 2.67 ± 0.04 a | 2.20 ± 0.15 b | 2.18 ± 0.16 b | 1.96 ± 0.30 b | 0.93 ± 0.12 c |
| Average | 2.16 ± 0.17 | 1.81 ± 0.16 | 1.72 ± 0.16 | 1.41 ± 0.22 | 0.89 ± 0.14 |
| Coefficient of variation (%) | 7.71 | 8.97 | 9.21 | 15.81 | 15.22 |
| Typical photographs of samples |  |  |  |  |  |
| Growth Periods | Selected Variables (from the Largest to Smallest Importance) | CV | Number of Variables | V-R2 | V-RMSE | CVRMSE (%) |
|---|---|---|---|---|---|---|
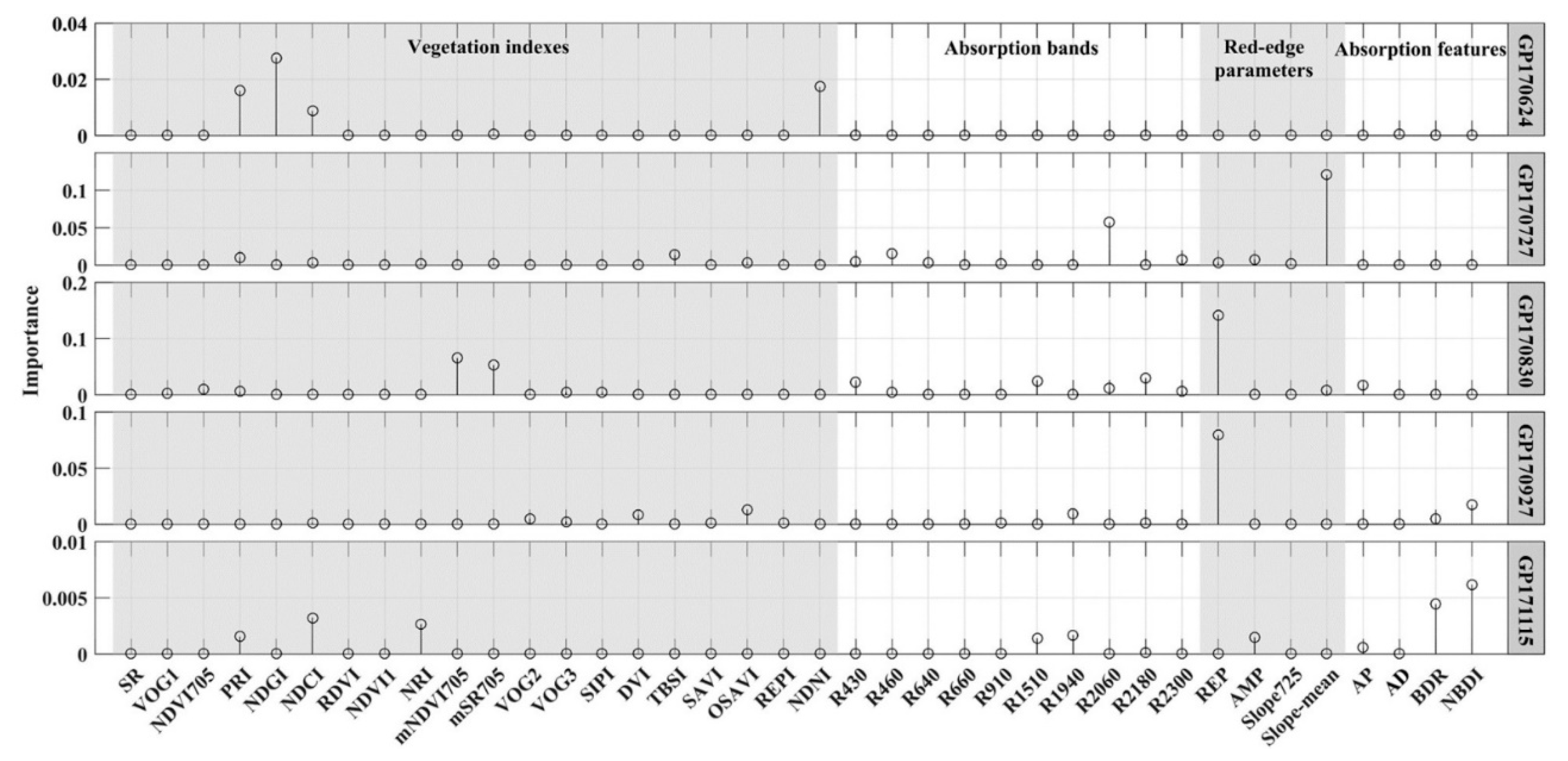
| GP170624 | NDGI, NDNI, PRI1, NDCI | LOO | 4 | 0.68 | 0.2046 | 9.46 |
| GP170727 | Slope_mean, R2060, R460, TBSI | LOO | 4 | 0.62 | 0.1870 | 10.32 |
| GP170830 | REP, mNDVI705, mSR705, R2180, R1510, R430 | LOO | 6 | 0.67 | 0.2202 | 12.78 |
| GP170927 | REP, NBDI, OSAVI, R1940, BDR | LOO | 5 | 0.58 | 0.2526 | 17.95 |
| GP171115 | NBDI, BDR, NDCI, NRI | LOO | 4 | 0.23 | 0.1341 | 15.12 |
| Selected Variables | Feature Selection Algorithm | CV | MAE | V-R2 | V-RMSE |
|---|---|---|---|---|---|
| NDNI, PRI, REP, Slope_mean, R640 | FFS | LOO | 0.29 | 0.51 | 0.3741 |
| FFS | LTO | 0.45 | 0.28 | 0.5412 | |
| FFS | LLO | 0.43 | 0.55 | 0.5114 | |
| FFS | LLTO | 0.38 | 0.29 | 0.4207 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, J.; Liang, T.; Yin, J.; Ge, J.; Feng, Q.; Wu, C.; Hou, M.; Liu, J.; Xie, H. Estimation of Alpine Grassland Forage Nitrogen Coupled with Hyperspectral Characteristics during Different Growth Periods on the Tibetan Plateau. Remote Sens. 2019, 11, 2085. https://doi.org/10.3390/rs11182085
Gao J, Liang T, Yin J, Ge J, Feng Q, Wu C, Hou M, Liu J, Xie H. Estimation of Alpine Grassland Forage Nitrogen Coupled with Hyperspectral Characteristics during Different Growth Periods on the Tibetan Plateau. Remote Sensing. 2019; 11(18):2085. https://doi.org/10.3390/rs11182085
Chicago/Turabian StyleGao, Jinlong, Tiangang Liang, Jianpeng Yin, Jing Ge, Qisheng Feng, Caixia Wu, Mengjing Hou, Jie Liu, and Hongjie Xie. 2019. "Estimation of Alpine Grassland Forage Nitrogen Coupled with Hyperspectral Characteristics during Different Growth Periods on the Tibetan Plateau" Remote Sensing 11, no. 18: 2085. https://doi.org/10.3390/rs11182085