Retrieving Secondary Forest Aboveground Biomass from Polarimetric ALOS-2 PALSAR-2 Data in the Brazilian Amazon

 , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
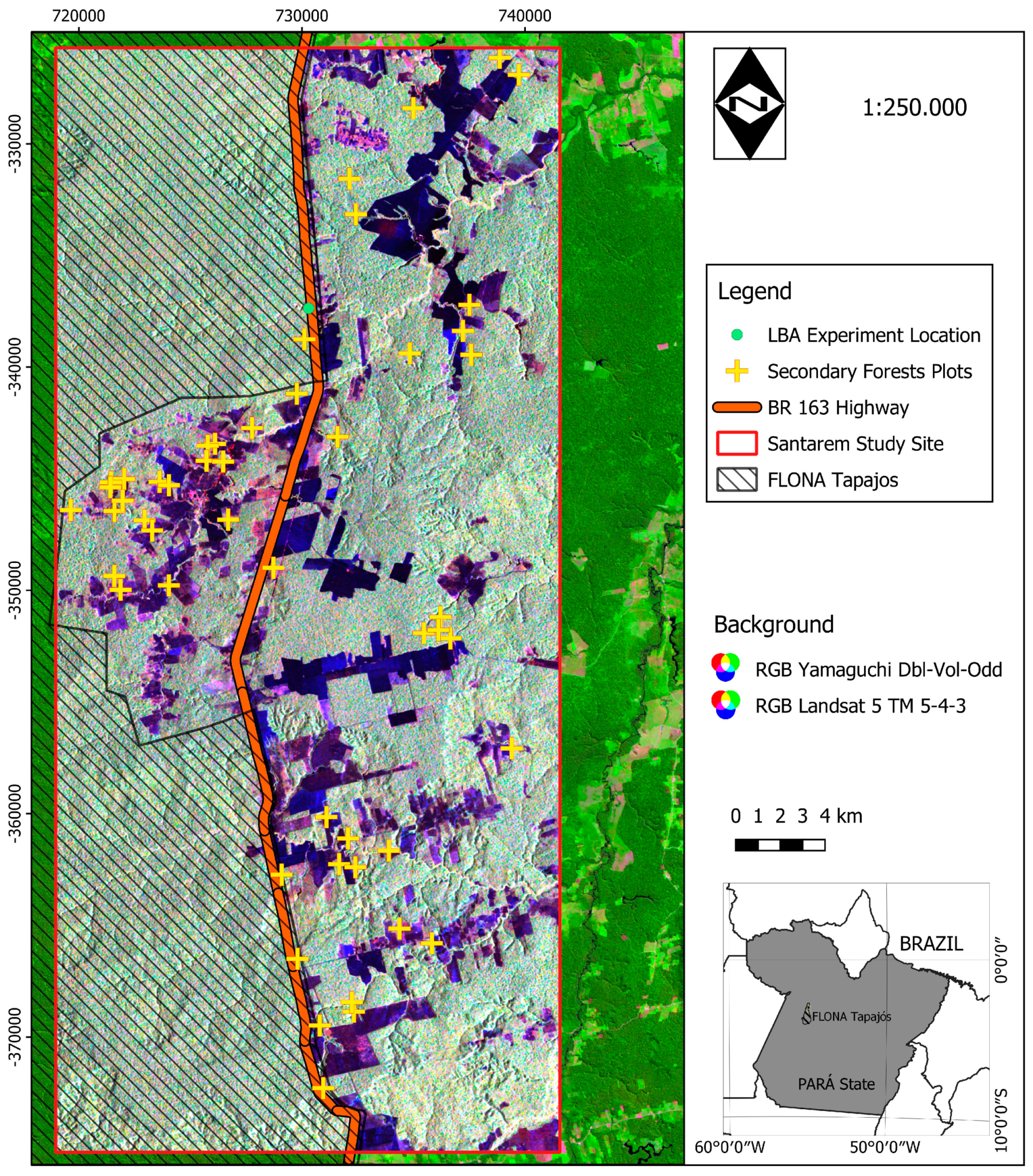
2.1. A Brief History of the Land-Use in the Santarém Region
2.2. Site Description
2.3. Field and Satellite Imagery Data
2.4. Aboveground Biomass (AGB) Allometric Equations
2.5. Growth Models for Biophysical Variables
2.6. Landsat Time-Series Classification
2.7. ALOS-2 PALSAR-2 Data Processing
2.8. Correlation Analysis
2.9. Models to Retrieve Aboveground Biomass
2.9.1. Non-Linear Regression Model
2.9.2. Multiple Linear Regression Model
2.10. Evaluation of Model’s Performance
3. Results
3.1. Structural Parameters of the Secondary Forests
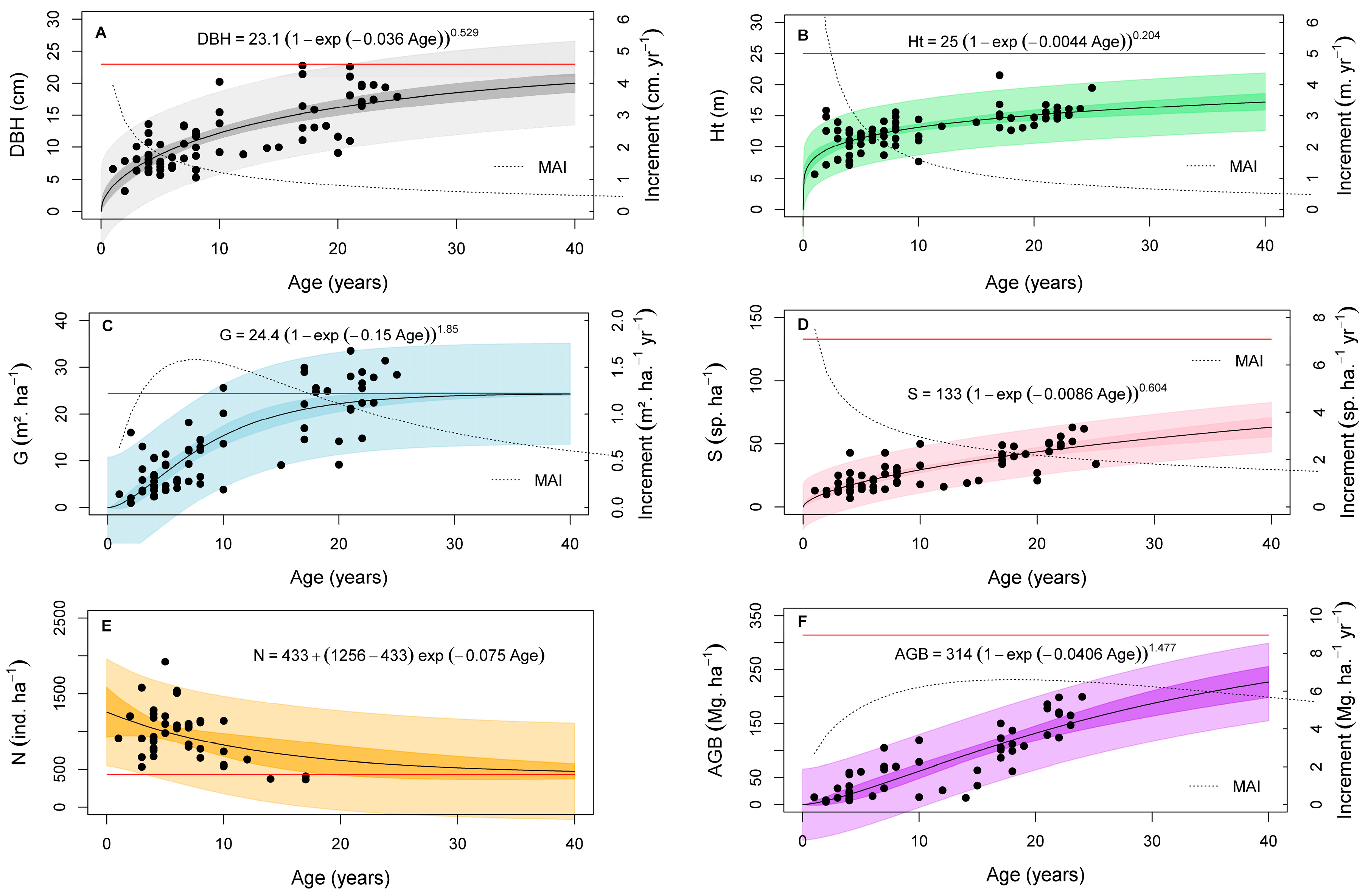
3.2. Growth Models and Accumulation Rates of the Secondary Forests Biophysical Variables
3.3. Characterization of Secondary Forest with Polarimetric Data
3.4. Modeling AGB
4. Discussion
4.1. Growth Models for Biophysical Variables
4.2. Polarimetric Data for SF Characterization
4.3. Regression Models
4.4. Uncertainty Report
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AdvSS | Advanced Secondary Succession |
|---|---|
| AIC | Akaike information criterion |
| ALOS-2 | Advanced Land Observation Satellite-2 |
| C, [C], [C]3 | Covariance 3 × 3 matrix |
| CDB | Convention on Biological Diversity |
| CF | Calibration coefficient factor (dB) |
| CFS | Correlation-based feature selection |
| DEM | Digital elevation model |
| FC | Frequency of cuts (× times) |
| FLONA | National forest, in Portuguese |
| FR | Faraday rotation (Ω) |
| FRA | Forest resources assessment |
| I | Phase image |
| IntSS | Intermediate secondary succession |
| ISS | Initial secondary succession |
| LULUCF | Land use, land-use change, and forestry |
| MAI | Mean annual increment |
| MCSM | Multiple-component scattering model |
| MLR | Multiple linear regression |
| NL | Non-linear |
| OLS | Ordinary least squares |
| PALSAR-2 | Phased Array L-band Synthetic Aperture Radar-2 |
| PALU | Period of active land use (years) |
| PF | Primary forest |
| PRODES | Program for Monitoring Deforestation of the Amazon by Satellite |
| Q | Complex image phase quadrature |
| R | R Foundation for statistical computing |
| SAR | Synthetic aperture radar |
| SF | Secondary forest |
| SLC | Single look complex |
| T, [T], [T]3 | Coherency 3 × 3 matrix |
| TanDEM | TerraSAR-X add-on for digital elevation measurement |
| TEC | Total electron content (electrons m−2) |
| TSVM | Target scattering vector model |
| VIF | Variance inflation factor |
| BP | Breusch–Pagan test |
| C | Organic carbon |
| CO2 | Carbon dioxide |
| Mg | Mega grams (106 g) |
| Pg | Pentagrams (1015 g) |
| R2 | Coefficient of determination |
| R2adj. | Coefficient of determination adjusted |
| RMSE | Root mean square error |
| |ρhh-vv| | Magnitude of complex coherency between linear polarizations (linear units) ([0, 1]; 0 = depolarized, 1 = completely polarized) |
| A | Asymptote |
| A | Anisotropy from Cloude & Pottier decomposition, related to the relative importance of secondary and tertiary scattering types A = (λ2 − λ3)/(λ2 + λ3) |
| AGB | Aboveground Biomass (kg or Mg ha−1) |
| alpha, α | Dominant scattering type angle in the trihedral-dihedral basis ([0, 90]; 0 = surface scattering, 45 = isotropic scattering (volumetric), 90 = double-bounce) |
| BMI | Biomass Index is an indicator of the relative amount of woody compared to leafy biomass |
| c | Shape of the curve (inflection point) |
| CSI | Canopy structure index is the relative importance of vertical versus horizontal structure in the vegetation |
| DBH | Diameter at the breast height (cm) |
| DERD, DERD_norm | Double-bounce Eigenvalue relative difference, and normalized DERD ([0, 1]; 0 = 0 = multiple scattering mechanism, 1 = dominant double-bounce scattering) |
| Forest | Forest backscattering coefficient |
| G | Basal area (m2 ha−1) |
| h | Tree height (m) |
| H | Entropy from Cloude & Pottier decomposition ([0, 1]; 0 = dominant scattering mechanism targets, 1 = random distributed targets) |
| H_A | H(1–A) highlights a random scattering process (high H and low A, λ1, λ2, λ3 |
| HA | H(A) highlights the presence of two scattering mechanisms with same pseudo-probabilities, high H and A, as forest targets ~λ3 |
| Ht | Mean total tree height (m) |
| I_Cnm_imag, Cnm_imag | Imaginary term nm (row × column) of the covariance matrix (linear units) |
| I_Cnm_real, Cnm_imag | Real term nm (row × column) of the covariance matrix (linear units) |
| k | Growth rate |
| N | Tree density (ind ha−1) |
| N0 | Tree density at t = 0 |
| Na | Tree density at t = ∞ |
| Neumann_mDelta | Scattering anisotropy magnitude from Neumann decomposition ([0, 2]; 0 = isotropic sphere, 1 = dipole) |
| Neumann_phDelta | |
| Neumann_tau | |
| P | |
| Ps | Proportion of odd-bounce or superficial scattering mechanism |
| Pd | Proportion of double-bounce scattering mechanism |
| Pv | Proportion of volumetric scattering mechanism |
| p1, p2, p3 | Pseudo-probabilities of eigenvalues λi from Cloude & Pottier decomposition ([0, 1]) |
| PF | Polarization fraction given by the difference between completely depolarized waves ([0, 1]; 0 = random distributed targets, 1 = dominant (pure) targets) |
| PH | |
| phi, φ | Scattering type phase difference between trihedral-dihedral ([−90, 90]; 0 = phase ambiguity, −90° = trihedral, 90° dihedral) |
| psi, ψ | Target tilt angle related to orientation angle of SAR line-of-sight ([−45, 45]; ±45 = preferred orientation direction, 0 = completely random orientation) |
| Rcp | |
| RFDI | Radar Forest Degradation Index, is the normalized difference between σ0 hh and σ0 hv |
| Rpp | |
| RVI | |
| S | Species richness (sp ha−1) |
| SE | Shannon entropy, sum of the contribution of intensity (SE_I) and polarimetry (SE_P), in dB |
| SE_I, SE_I_norm | |
| SE_P, SE_P_norm | Contribution of SE polarimetry normalized ([0, 1]; 0 = depolarized entropy, 1 = polarized entropy) , |
| SERD, SERD_norm | Single-bounce Eigenvalue relative difference, and normalized SERD ([0, 1]; 0 = multiple scattering mechanism, 1 = dominant single-bounce scattering) |
| Span | |
| t | Age |
| tau, τ | Helicity angle related to assess target symmetry ([−45, 45] 0 = symmetric target angle, ±45 = non-symmetric target angle) |
| Tnm_imag_Z | Imaginary term nm (row × column) of the coherency matrix from the Z decomposition, in linear units |
| Tnm_real_Z | Real term nm (row × column) of the coherency matrix from the Z decomposition, in linear units |
| VSI | Vegetation Scattering Index is an indicator of canopy thickness or density |
| Xi | Explanatory variable of the regression model |
| Yi | Average of the AGB i-th observation of the regression model |
| Yt | Size of the variable at age t |
| Z_Dbl | Proportion of double-bounce scattering mechanism from the Z decomposition |
| Z_Hlx | Proportion of helix scattering mechanism from the Z decomposition |
| Z_Odd | Proportion of odd (surface) scattering mechanism from the Z decomposition |
| Z_Vol | Proportion of volumetric scattering mechanism from the Z decomposition |
| Z_Wire | Proportion of wire scattering mechanism from the Z decomposition |
| α, β, λ, γ, δ | Terms of the Cloude eigenvectors decomposition, respectively: dominant scattering type angle, target tilt angle, mean target eigenvalue, and phase differences between channels |
| ε | Experimental error of the regression model |
| ρ | Wood density in g cm−3 |
| ρhh-vv | |
| σ° | Sigma-naught, in dB |
| σ° for | Total backscatter coefficient of the forest, in dB |
| χ2 | Chi-square test |
References
- CDB. Definitions: Indicative Definitions Taken from the Report of the ad hoc Technical Expert Group on Forest Biological Diversity. 2014. Available online: https://www.cbd.int/forest/definitions.shtml (accessed on 18 September 2014).
- Brown, S.; Lugo, A.E. Tropical secondary forests. J. Trop. Ecol. 1990, 6, 1–32. [Google Scholar] [CrossRef]
- Corlett, R.T. What is Secondary Forest? J. Trop. Ecol. 1994, 10, 445–447. [Google Scholar] [CrossRef]
- Whitmore, T.C. An Introduction to Tropical Rainforests, 2nd ed.; Oxford University Press: New York, NY, USA, 1998; ISBN 978-0-19-850147-3. [Google Scholar]
- FAO. Workshop on Tropical Secondary Forest Management in Africa: Reality and Perspectives. 2003. Available online: http://www.fao.org/docrep/006/j0628e/J0628E10.htm#P572_41806 (accessed on 15 May 2014).
- FAO. Global Forest Resources Assessment 2000. FAO Forestry Paper, n. 140. Roma: Main Report. 2001. Available online: http://www.fao.org/forestry/fra/fra2010/en/ (accessed on 10 September 2014).
- FAO. Global Forest Resources Assessment 2010. FAO Forestry Paper, n. 163. Roma: Main Report. 2010. Available online: http://www.fao.org/forestry/fra/86624/en/ (accessed on 10 September 2014).
- Achard, F. Determination of Deforestation Rates of the World’s Humid Tropical Forests. Science 2002, 297, 999–1002. [Google Scholar] [CrossRef] [PubMed]
- Chazdon, R.L. Tropical forest recovery: Legacies of human impact and natural disturbances. Perspect. Plant Ecol. Evol. Syst. 2003, 6, 51–71. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Gibson, L.; Lee, T.M.; Koh, L.P.; Brook, B.W.; Gardner, T.A.; Barlow, J.; Peres, C.A.; Bradshaw, C.J.A.; Laurance, W.F.; Lovejoy, T.E.; et al. Erratum: Corrigendum: Primary forests are irreplaceable for sustaining tropical biodiversity. Nature 2014, 505, 710. [Google Scholar] [CrossRef]
- Chazdon, R.L. Second Growth: The Promise of Tropical Forest Regeneration in an Age of Deforestation; Chicago Press: Chicago, IL, USA, 2014; ISBN 978-0-22-611807-9. [Google Scholar]
- Harris, N.L.; Brown, S.; Hagen, S.C.; Saatchi, S.S.; Petrova, S.; Salas, W.; Hansen, M.C.; Potapov, P.V.; Lotsch, A. Baseline Map of Carbon Emissions from Deforestation in Tropical Regions. Science 2012, 336, 1573–1576. [Google Scholar] [CrossRef]
- Chazdon, R.L.; Broadbent, E.N.; Rozendaal, D.M.A.; Bongers, F.; Zambrano, A.M.A.; Aide, T.M.; Balvanera, P.; Becknell, J.M.; Boukili, V.; Brancalion, P.H.S.; et al. Carbon sequestration potential of second-growth forest regeneration in the Latin American tropics. Sci. Adv. 2016, 2, e1501639. [Google Scholar] [CrossRef] [Green Version]
- Aragão, L.E.O.C.; Poulter, B.; Barlow, J.B.; Anderson, L.O.; Malhi, Y.; Saatchi, S.; Phillips, O.L.; Gloor, E. Environmental change and the carbon balance of Amazonian forests. Biol. Rev. 2014, 89, 913–931. [Google Scholar] [CrossRef]
- Houghton, R.A.; Skole, D.L.; Nobre, C.A.; Hackler, J.L.; Lawrence, K.T.; Chomentowski, W.H. Annual fluxes of carbon from deforestation and regrowth in the Brazilian Amazon. Nature 2000, 403, 301–304. [Google Scholar] [CrossRef]
- Gehring, C.; Denich, M.; Vlek, P.L.G. Resilience of secondary forest regrowth after slash-and-burn agriculture in central Amazonia. J. Trop. Ecol. 2005, 21, 519–527. [Google Scholar] [CrossRef]
- Uhl, A.C.; Buschbacher, R.; Serrao, E.A.S.; Uhl, C.; Buschbachertt, R. Abandoned Pastures in Eastern Amazonia. I. Patterns of Plant Succession. J. Ecol. 1988, 76, 663–681. [Google Scholar] [CrossRef]
- Tucker, J.M.; Brondizio, E.S.; Moran, E.F. Rates of forest regrowth in Eastern Amazonia: A comparision of Altamira and Bragantina regions, Para state, Brazil. Interciencia 1998, 23, 64–73. [Google Scholar]
- Junqueira, A.B.; Shepard, G.H.; Clement, C.R. Secondary forests on anthropogenic soils in Brazilian Amazonia conserve agrobiodiversity. Biodivers. Conserv. 2010, 19, 1933–1961. [Google Scholar] [CrossRef]
- Prates-Clark, C.D.C.; Lucas, R.M.; dos Santos, J.R. Implications of land-use history for forest regeneration in the Brazilian Amazon. Can. J. Remote Sens. 2009, 35, 534–553. [Google Scholar] [CrossRef]
- Steininger, M.K. Secondary forest structure and biomass following short and extended land-use in central and southern Amazonia. J. Trop. Ecol. 2000, 16, 689–708. [Google Scholar] [CrossRef]
- Alves, D.; Soares, J.V.; Amaral, S.; Mello, E.; Almeida, S.; Silva, O.F.; Silveira, A. Biomass of primary and secondary vegetation in Rondonia, Western Brazilian Amazon. Glob. Chang. Biol. 1997, 3, 451–461. [Google Scholar] [CrossRef]
- Vieira, I.C.G.; de Paiva Salomão, R.; de Araujo Rosa, N.; Nepstad, D.C.; Roma, J.C. O renascimento da floresta no rastro da agricultura. Ciênc. Hoje 1996, 20, 38–44. [Google Scholar]
- Neeff, T.; dos Santos, J.R. A growth model for secondary forest in Central Amazonia. For. Ecol. Manag. 2005, 216, 270–282. [Google Scholar] [CrossRef]
- Wandelli, E.V.; Fearnside, P.M. Secondary vegetation in central Amazonia: Land-use history effects on aboveground biomass. For. Ecol. Manag. 2015, 347, 140–148. [Google Scholar] [CrossRef]
- Carreiras, J.M.B.; Pereira, J.M.C.; Campagnolo, M.L.; Shimabukuro, Y.E. Assessing the extent of agriculture/pasture and secondary succession forest in the Brazilian Legal Amazon using SPOT VEGETATION data. Remote Sens. Environ. 2006, 101, 283–298. [Google Scholar] [CrossRef]
- Carreiras, J.M.B.; Jones, J.; Lucas, R.M.; Gabriel, C. Land Use and Land Cover Change Dynamics across the Brazilian Amazon: Insights from Extensive Time-Series Analysis of Remote Sensing Data. PLoS ONE 2014, 9, e104144. [Google Scholar] [CrossRef] [PubMed]
- Almeida, C.A.; Valeriano, D.M.; Escada, M.I.S.; Rennó, C.D. Estimativa de área de vegetação secundária na Amazônia Legal Brasileira. Acta Amaz. 2010, 40, 289–301. [Google Scholar] [CrossRef] [Green Version]
- INPE. Projeto PRODES: Monitoramento da Floresta Amazônica Brasileira por Satélite. 2014. Available online: http://www.obt.inpe.br/prodes/index.php (accessed on 20 September 2014).
- Lucas, R.M.; Xiao, X.; Hagen, S.; Frolking, S. Evaluating TERRA-1 MODIS data for discrimination of tropical secondary forest regeneration stages in the Brazilian Legal Amazon. Geophys. Res. Lett. 2002, 29, 42-1–42-4. [Google Scholar] [CrossRef]
- Asner, G.P.; Rudel, T.K.; Aide, T.M.; Defries, R.; Emerson, R. A Contemporary Assessment of Change in Humid Tropical Forests. Conserv. Biol. 2009, 23, 1386–1395. [Google Scholar] [CrossRef] [PubMed]
- INPE. Projeto Terra Class: Levantamento de Informações de uso e Cobertura da Terra na Amazônia. 2010. Available online: http://www.inpe.br/cra/projetos_pesquisas/terraclass2010.php (accessed on 20 September 2014).
- Lucas, R.M.; Honzák, M.; Do Amaral, I.; Curran, P.J.; Foody, G.M. Forest regeneration on abandoned clearances in central Amazonia. Int. J. Remote Sens. 2002, 23, 965–988. [Google Scholar] [CrossRef]
- Zarin, D.J.; Davidson, E.A.; Brondizio, E.; Vieira, I.C.G.; Sá, T.; Schuur, E.A.G.; Mesquita, R.; Moran, E.; Delamonica, P.; Mark, J.; et al. Legacy of fire slows carbon accumulation in Amazonian forest regrowth. Ecology 2005, 3, 365–369. [Google Scholar] [CrossRef] [Green Version]
- Bonner, M.T.L.; Schmidt, S.; Shoo, L.P. A meta-analytical global comparison of aboveground biomass accumulation between tropical secondary forests and monoculture plantations. For. Ecol. Manag. 2013, 291, 73–86. [Google Scholar] [CrossRef]
- Bispo, P.C.; Santos, J.R.; Valeriano, M.M.; Touzi, R.; Seifert, F.M. Integration of Polarimetric PALSAR Attributes and Local Geomorphometric Variables Derived from SRTM for Forest Biomass Modeling in Central Amazonia. Can. J. Remote Sens. 2014, 40, 26–42. [Google Scholar] [CrossRef]
- Vafaei, S.; Soosani, J.; Adeli, K.; Fadaei, H.; Naghavi, H.; Pham, T.; Tien Bui, D. Improving Accuracy Estimation of Forest Aboveground Biomass Based on Incorporation of ALOS-2 PALSAR-2 and Sentinel-2A Imagery and Machine Learning: A Case Study of the Hyrcanian Forest Area (Iran). Remote Sens. 2018, 10, 172. [Google Scholar] [CrossRef]
- Saatchi, S.; Halligan, K.; Despain, D.G.; Crabtree, R.L. Estimation of Forest Fuel Load From Radar Remote Sensing. IEEE Trans. Geosci. Remote Sens. 2007, 45, 1726–1740. [Google Scholar] [CrossRef]
- Vaglio Laurin, G.; Pirotti, F.; Callegari, M.; Chen, Q.; Cuozzo, G.; Lingua, E.; Notarnicola, C.; Papale, D. Potential of ALOS2 and NDVI to Estimate Forest Above-Ground Biomass, and Comparison with Lidar-Derived Estimates. Remote Sens. 2016, 9, 18. [Google Scholar] [CrossRef]
- Sinha, S.; Jeganathan, C.; Sharma, L.K.; Nathawat, M.S. A review of radar remote sensing for biomass estimation. Int. J. Environ. Sci. Technol. 2015, 12, 1779–1792. [Google Scholar] [CrossRef] [Green Version]
- Woodhouse, I.H. Predicting backscatter-biomass and height-biomass trends using a macroecology model. IEEE Trans. Geosci. Remote Sens. 2006, 44, 871–877. [Google Scholar] [CrossRef]
- Ghasemi, N.; Sahebi, M.R.; Mohammadzadeh, A. Biomass estimation of a temperate deciduous forest using wavelet analysis. IEEE Trans. Geosci. Remote Sens. 2013, 51, 765–776. [Google Scholar] [CrossRef]
- Avtar, R.; Suzuki, R.; Sawada, H. Natural Forest Biomass Estimation Based on Plantation Information Using PALSAR Data. PLoS ONE 2014, 9, e86121. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.R.; Silva, C.V.D.J.; Galvão, L.S.; Treuhaft, R.; Mura, J.C.; Madsen, S.; Gonçalves, F.G.; Keller, M.M. Determining aboveground biomass of the forest successional chronosequence in a test-site of Brazilian Amazon through X- and L-band data analysis. In Proceedings of the Second International Conference on Remote Sensing and Geoinformation of the Environment (RSCy2014), Paphos, Cyprus, 7–10 April 2014; Hadjimitsis, D.G., Themistocleous, K., Michaelides, S., Papadavid, G., Eds.; SPIE: Bellingham, WA, USA, 2014; Volume 9229, p. 92291E. [Google Scholar]
- Pereira, L.O.; Furtado, L.F.A.; Novo, E.M.L.M.; Anna, S.J.S.S. Multifrequency and Full-polarimetric SAR Assessment for estimating Above Ground Biomass and Leaf Area Index in the Amazon Várzea Wetlands. Remote Sens. 2018, 10, 1355. [Google Scholar] [CrossRef]
- Scatena, F.N.; Walker, R.T.; Homma, A.K.O.; de Conto, A.J.; Ferreira, C.A.P.; de Amorim Carvalho, R.; da Rocha, A.C.P.N.; dos Santos, A.I.M.; de Oliveira, P.M. Cropping and fallowing sequences of small farms in the “terra firme” landscape of the Brazilian Amazon: A case study from Santarem, Para. Ecol. Econ. 1996, 18, 29–40. [Google Scholar] [CrossRef]
- Roosevelt, A.C.; Housley, R.A.; Da Silveira, M.I.; Maranca, S.; Johnson, R. Eighth Millennium Pottery from a Prehistoric Shell Midden in the Brazilian Amazon. Science 1991, 254, 1621–1624. [Google Scholar] [CrossRef]
- Fearnside, P.M. Deforestation in Brazilian Amazonia: History, Rates, and Consequences. Conserv. Biol. 2005, 19, 680–688. [Google Scholar] [CrossRef]
- Aragão, L.E.O.C.; Anderson, L.O.; Fonseca, M.G.; Rosan, T.M.; Vedovato, L.B.; Wagner, F.H.; Silva, C.V.J.; Silva Junior, C.H.L.; Arai, E.; Aguiar, A.P.; et al. 21st Century drought-related fires counteract the decline of Amazon deforestation carbon emissions. Nat. Commun. 2018, 9, 536. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, F.G.; Santos, J.R. dos Composição florística e estrutura de uma unidade de manejo florestal sustentável na Floresta Nacional do Tapajós, Pará. Acta Amaz. 2008, 38, 229–244. [Google Scholar] [CrossRef]
- Vieira, S.; de Camargo, P.B.; Selhorst, D.; da Silva, R.; Hutyra, L.; Chambers, J.Q.; Brown, I.F.; Higuchi, N.; dos Santos, J.; Wofsy, S.C.; et al. Forest structure and carbon dynamics in Amazonian tropical rain forests. Oecologia 2004, 140, 468–479. [Google Scholar] [CrossRef] [Green Version]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef]
- Quesada, C.A.; Lloyd, J.; Schwarz, M.; Patiño, S.; Baker, T.R.; Czimczik, C.; Fyllas, N.M.; Martinelli, L.; Nardoto, G.B.; Schmerler, J.; et al. Variations in chemical and physical properties of Amazon forest soils in relation to their genesis. Biogeosciences 2010, 7, 1515–1541. [Google Scholar] [CrossRef] [Green Version]
- Silver, W.L.; Neff, J.; McGroddy, M.; Veldkamp, E.; Keller, M.; Cosme, R. Effects of Soil Texture on Belowground Carbon and Nutrient Storage in a Lowland Amazonian Forest Ecosystem. Ecosystems 2000, 3, 193–209. [Google Scholar] [CrossRef]
- Telles, E.D.; de Camargo, P.B.; Martinelli, L.A.; Trumbore, S.E.; da Costa, E.S.; Santos, J.; Higuchi, N.; Oliveira, R.C. Influence of soil texture on carbon dynamics and storage potential in tropical forest soils of Amazonia. Glob. Biogeochem. Cycles 2003, 17. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.; Moran, E.; Mausel, P. Linking Amazonian secondary succession forest growth to soil properties. Land Degrad. Dev. 2002, 13, 331–343. [Google Scholar] [CrossRef]
- De Castilho, C.V.; Magnusson, W.E.; de Araújo, R.N.O.; Luizão, R.C.C.; Luizão, F.J.; Lima, A.P.; Higuchi, N. Variation in aboveground tree live biomass in a central Amazonian Forest: Effects of soil and topography. For. Ecol. Manag. 2006, 234, 85–96. [Google Scholar] [CrossRef]
- IBGE. Manual Técnico da Vegetação Brasileira: Sistema Fitogeográfico. Inventário das Formações Florestais e Campestres. Técnicas e Manejo de Coleções Botânicas. Procedimentos para Mapeamentos, 2nd ed.; IBGE: Brasília, Brazil, 2012; ISBN 978-8-52-404272-0.
- Vieira, S.; Trumbore, S.; Camargo, P.B.; Selhorst, D.; Chambers, J.Q.; Higuchi, N.; Martinelli, L.A. Slow growth rates of Amazonian trees: Consequences for carbon cycling. Proc. Natl. Acad. Sci. USA 2005, 102, 18502–18507. [Google Scholar] [CrossRef] [Green Version]
- Pyle, E.H.; Santoni, G.W.; Nascimento, H.E.M.; Hutyra, L.R.; Vieira, S.; Curran, D.J.; van Haren, J.; Saleska, S.R.; Chow, V.Y.; Carmago, P.B.; et al. Dynamics of carbon, biomass, and structure in two Amazonian forests. J. Geophys. Res. Biogeosci. 2008, 113. [Google Scholar] [CrossRef] [Green Version]
- Hunter, M.O.; Keller, M.; Morton, D.; Cook, B.; Lefsky, M.; Ducey, M.; Saleska, S.; de Oliveira, R.C.; Schietti, J. Structural Dynamics of Tropical Moist Forest Gaps. PLoS ONE 2015, 10, e0132144. [Google Scholar] [CrossRef] [PubMed]
- Cassol, H.L.G.; Shimabukuro, Y.E.; Carreiras, J.M.D.B.; Moraes, E.C. Improved tree height estimation of secondary forests in the Brazilian Amazon. Acta Amaz. 2018, 48, 179–190. [Google Scholar] [CrossRef]
- Silva, C.V.D.J.; dos Santos, J.R.; Galvão, L.S.; da Silva, R.D.; Moura, Y.M. Floristic and structure of an Amazonian primary forest and a chronosequence of secondary succession. Acta Amaz. 2016, 46, 133–150. [Google Scholar] [CrossRef] [Green Version]
- Cassol, H.L.G. Aplicação Dos Dados Polarimétricos Alos/Palsar-2 Para Modelagem De Biomassa. Ph.D. Thesis, Instituto Nacional de Pesquisas Espaciais, São José dos Campos, Brazil, 2017. [Google Scholar]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.C.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.; Gillespie, A.J.R.; Lugo, A.E. Biomass estimation methods for tropical forests with applications to forest inventory data. For. Sci. 1989, 35, 881–902. [Google Scholar]
- Zanne, A.E.; Lopez-Gonzalez, G.; Coomes, D.A.; Ilic, J.; Jansen, S.; Lewis, S.L.; Miller, R.B.; Swenson, N.G.; Wiemann, M.C.; Chave, J. Global wood density database. Dryad 2009, 235, 33. [Google Scholar] [CrossRef]
- Cummings, D.L.; Boone Kauffman, J.; Perry, D.A.; Flint Hughes, R. Aboveground biomass and structure of rainforests in the southwestern Brazilian Amazon. For. Ecol. Manag. 2002, 163, 293–307. [Google Scholar] [CrossRef]
- Pienaar, L.V.; Turnbull, K.J. The Chapman-Richards generalization of von Bertalanffy’s growth model for basal area growth and yield in even-aged stands. For. Sci. 1973, 19, 2–22. [Google Scholar]
- Draper, N.R.; Smith, H. Applied Regression Analysis, 2nd ed.; John Wiley: New York, NY, USA, 1981; 709p, ISBN 978-0-471-17082-2. [Google Scholar]
- Luckman, A.; Baker, J.; Kuplich, T.M.; Freitas, C.C.; Frery, A.C. A study of the relationship between radar backscatter and regenerating tropical forest biomass for spaceborne SAR instruments. Remote Sens. Environ. 1997, 60, 1–13. [Google Scholar] [CrossRef]
- Prates-Clark, C.C. Remote Sensing of Tropical Regenerating Forests in the Brazilian Amazon. Ph.D. Thesis, University of Wales, Aberystwyth, UK, 2004. [Google Scholar]
- Santos, J. Airborne P-band SAR applied to the aboveground biomass studies in the Brazilian tropical rainforest. Remote Sens. Environ. 2003, 87, 482–493. [Google Scholar] [CrossRef]
- Robinson, A.P.; Hamann, J.D. Forest Analytics with R; Gentleman, R., Hornik, K., Parmigiani, G., Eds.; Springer: New York, NY, USA, 2011; ISBN 978-1-4419-7761-8. [Google Scholar]
- Neter, J.; Kutner, N.H.; Nachtssheim, C.J.; Wasserman, W. Applied Linear Statistical Models, 4th ed.; McGraw-Hill: Boston, MA, USA, 1996; 1408p. [Google Scholar]
- Bates, D.M.; Watts, D.G. Nonlinear Regression Analysis and Its Applications, 2nd ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1990; ISBN 978-0-470-13900-4. [Google Scholar]
- Pretzsch, H. Forest Dynamics, Growth and Yield; Springer: Berlin/Heidelberg, Germany, 2010; ISBN 978-3-540-88306-7. [Google Scholar]
- Baker, T.R.; Phillips, O.L.; Malhi, Y.; Almeida, S.; Arroyo, L.; Di Fiore, A.; Erwin, T.; Killeen, T.J.; Laurance, S.G.; Laurance, W.F.; et al. Variation in wood density determines spatial patterns in Amazonian forest biomass. Glob. Chang. Biol. 2004, 10, 545–562. [Google Scholar] [CrossRef]
- Silva, C.V.D.J. Caracterização Florístico-Estrutural e Modelagem de Biomassa na Floresta Amazônica a Partir de Dados ALOS/PALSAR e TERRASAR/TANDEM-X. Ph.D. Thesis, Instituto Nacional de Pesquisas Espaciais, São José dos Campos, Brazil, 2014. [Google Scholar]
- Chambers, J.Q.; dos Santos, J.; Ribeiro, R.J.; Higuchi, N. Tree damage, allometric relationships, and above-ground net primary production in central Amazon forest. For. Ecol. Manag. 2001, 152, 73–84. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, A.A.; Mori, S.A. A central Amazonian terra firme forest. I. High tree species richness on poor soils. Biodivers. Conserv. 1999, 8, 1219–1244. [Google Scholar] [CrossRef]
- Espírito-Santo, F.D.B. Caracterização e Mapeamento da Vegetação da Região da Floresta Nacional do Tapajós Através de Dados Óticos, Radar e Inventários Florestais. Ph.D. Thesis, Instituto Nacional de Pesquisas Espaciais, São José dos Campos, Brazil, 2003. [Google Scholar]
- Holm, J.A.; Chambers, J.Q.; Collins, W.D.; Higuchi, N. Forest response to increased disturbance in the central Amazon and comparison to western Amazonian forests. Biogeosciences 2014, 11, 5773–5794. [Google Scholar] [CrossRef] [Green Version]
- Hunter, M.O.; Keller, M.; Victoria, D.; Morton, D.C. Tree height and tropical forest biomass estimation. Biogeosciences 2013, 10, 8385–8399. [Google Scholar] [CrossRef] [Green Version]
- Marra, D.M.; Higuchi, N.; Trumbore, S.E.; Ribeiro, G.H.; Santos, J.D.; Carneiro, V.M.; Lima, A.J.; Chambers, J.Q.; Negrón-Juárez, R.I.; Holzwarth, F.; et al. Predicting biomass of hyperdiverse and structurally complex central Amazonian forests—A virtual approach using extensive field data. Biogeosciences 2016, 13, 1553–1570. [Google Scholar] [CrossRef]
- Milliken, W. Structure and Composition of One Hectare of Cental Amazonian Terra Firme Forest. Biotropica 1998, 30, 530–537. [Google Scholar] [CrossRef]
- Narvaes, I.D.S. Avaliação de Dados SAR Polarimétricos Para Estimativa de Biomassa em Diferentes Fitofisionomias de Florestas Tropicais. Ph.D. Thesis, Instituto Nacional de Pesquisas Espaciais, São José dos Campos, Brazil, 2010. [Google Scholar]
- Nascimento, H.E.M.; Laurance, W.F. Total aboveground biomass in central Amazonian rainforests: A landscape-scale study. For. Ecol. Manag. 2002, 168, 311–321. [Google Scholar] [CrossRef]
- Rankin-de-Mérona, J.M.; Prance, G.T.; Hutchings, R.W.; Freitas da Silva, M.; Rodrigues, W.A.; Uehling, M.E. Preliminary results of a large-scale tree inventory of upland rain forest in the central Amazon. Acta Amaz. 1992, 22, 493–534. [Google Scholar] [CrossRef]
- Suwa, R.; Sakai, T.; Dos Santos, J.; Da Silva, R.P.; Kajimoto, T.; Ishizuka, M.; Higuchi, N. Significance of Topographic Gradient in Stem Diameter—Height Allometry for Precise Biomass Estimation of a Tropical Moist Forest in the Central Amazon. Jpn. Agric. Res. Q. 2013, 47, 109–114. [Google Scholar] [CrossRef]
- Keller, M.; Palace, M.; Hurtt, G. Biomass estimation in the Tapajos National Forest, Brazil. For. Ecol. Manag. 2001, 154, 371–382. [Google Scholar] [CrossRef]
- Shimabukuro, Y.E.; Smith, J.A. The least-squares mixing models to generate fraction images derived from remote sensing multispectral data. IEEE Trans. Geosci. Remote Sens. 1991, 29, 16–20. [Google Scholar] [CrossRef]
- Chander, G.; Markham, B.L.; Helder, D.L. Summary of current radiometric calibration coefficients for Landsat MSS, TM, ETM+, and EO-1 ALI sensors. Remote Sens. Environ. 2009, 113, 893–903. [Google Scholar] [CrossRef] [Green Version]
- JAXA. Japan Aerospace Exploration Agency ALOS/PALSAR-2. Tokyo, Japan, 2015. Available online: http://global.jaxa.jp/projects/sat/ALOS2/pdf/daichi2_e.pdf (accessed on 15 April 2015).
- Instituto Nacional de Meteorologia (INMET). Historic Data Precipitation. Available online: http://www.inmet.gov.br/portal/index.php?r=home2/index (accessed on 20 May 2017).
- Freeman, A.; Saatchi, S.S. On the detection of Faraday rotation in linearly polarized L-band SAR backscatter signatures. IEEE Trans. Geosci. Remote Sens. 2004, 42, 1607–1616. [Google Scholar] [CrossRef]
- EMBRACE/INPE. TEC Map. Available online: http://www2.inpe.br/climaespacial/portal/tec-map-home/ (accessed on 20 February 2017).
- Wright, P.A.; Quegan, S.; Wheadon, N.S.; Hall, C.D. Faraday rotation effects on L-band spaceborne sar data. IEEE Trans. Geosci. Remote Sens. 2003, 41, 2735–2744. [Google Scholar] [CrossRef]
- Curlander, J.C. Location of Spaceborne Sar Imagery. IEEE Trans. Geosci. Remote Sens. 1982, GE-20, 359–364. [Google Scholar] [CrossRef]
- Small, D.; Schubert, A. Guide to ASAR Geocoding; University of Zürich: Zurich, Switzerland, 2008. [Google Scholar]
- Small, D. Flattening gamma: Radiometric terrain correction for SAR imagery. IEEE Trans. Geosci. Remote Sens. 2011, 49, 3081–3093. [Google Scholar] [CrossRef]
- Mermoz, S.; Réjou-Méchain, M.; Villard, L.; Le Toan, T.; Rossi, V.; Gourlet-Fleury, S. Decrease of L-band SAR backscatter with biomass of dense forests. Remote Sens. Environ. 2015, 159, 307–317. [Google Scholar] [CrossRef]
- Woodhouse, I.H. Introduction to Microwave Remote Sensing; Taylor & Francis Group CRC Press: Boca Raton, FL, USA, 2006; p. 370. ISBN 0-415-27123-1. [Google Scholar]
- Lee, J.-S.; Pottier, E. Polarimetric Radar Imaging: From Basics to Applications; Taylor & Francis Group CRC Press: New York, NY, USA, 2009; 440p, ISBN 978-1-4200-5497-2. [Google Scholar]
- Zhang, L.; Zou, B.; Cai, H.; Zhang, Y. Multiple-Component Scattering Model for Polarimetric SAR Image Decomposition. IEEE Geosci. Remote Sens. Lett. 2008, 5, 603–607. [Google Scholar] [CrossRef]
- Freeman, A.; Durden, S.L. A three-component scattering model for polarimetric SAR data. IEEE Trans. Geosci. Remote Sens. 1998, 36, 963–973. [Google Scholar] [CrossRef]
- Neumann, M. Remote Sensing of Vegetation Using Multi-Baseline Polarimetric SAR Interferometry: Theoretical Modeling and Physical Parameter Retrieval. Ph.D. Thesis, Université de Rennes, Rennes, France, 2009. [Google Scholar]
- Van Zyl, J.J. Application of Cloude’s target decomposition theorem to polarimetric imaging radar data. In Proceedings of the SPIE Conference on Radar Polarimetry (San Diego’92), San Diego, CA, USA, 22 July 1992; Mott, H., Boerner, W.-M., Eds.; SPIE: San Diego, CA, USA, 1993; Volume 1748, p. 184. [Google Scholar]
- Yamaguchi, Y.; Moriyama, T.; Ishido, M.; Yamada, H. Four-component scattering model for polarimetric SAR image decomposition. IEEE Trans. Geosci. Remote Sens. 2005, 43, 1699–1706. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Muhuri, A.; De, S.; Manickam, S.; Frery, A.C. Modifying the Yamaguchi Four-Component Decomposition Scattering Powers Using a Stochastic Distance. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2015, 8, 3497–3506. [Google Scholar] [CrossRef]
- Singh, G.; Yamaguchi, Y.; Park, S.-E. General Four-Component Scattering Power Decomposition with Unitary Transformation of Coherency Matrix. IEEE Trans. Geosci. Remote Sens. 2013, 51, 3014–3022. [Google Scholar] [CrossRef]
- Touzi, R. Target scattering decomposition of one-look and multi-look SAR data using a new coherent scattering model: The TSVM. In Proceedings of the IEEE International Geoscience and Remote Sensing Symposium (IGARSS’04), Anchorage, AK, USA, 20–24 September 2004; Volume 4, pp. 2491–2494. [Google Scholar]
- Cloude, S.; Pottier, E. An entropy based classification scheme for land applications of polarimetric SAR. IEEE Trans. Geosci. Remote Sens. 1997, 35, 68–78. [Google Scholar] [CrossRef]
- Huynen, J.R. Phenomenological Theory of Radar Targets. Ph.D. Thesis, Delft University of Technology, Rotetterdam, The Netherlands, 1970. [Google Scholar]
- Cloude, S.R. Target decomposition theorems in radar scattering. Electron. Lett. 1985, 21, 22–24. [Google Scholar] [CrossRef]
- Holm, W.A.; Barnes, R.M. On radar polarization mixed target state decomposition techniques. In Proceedings of the 1988 IEEE National Radar Conference, Ann Arbor, MI, USA, 20–21 April 1988; pp. 249–254. [Google Scholar]
- Réfrégier, P.; Morio, J. Shannon entropy of partially polarized and partially coherent light with Gaussian fluctuations. J. Opt. Soc. Am. A Opt. Image Sci. Vis. 2006, 23, 3036–3044. [Google Scholar] [CrossRef] [PubMed]
- Allain, S.; Ferro-Famil, L.; Pottier, E. New Eigenvalue-Based Parameters for Natural Media Characterization. In Proceedings of the European Radar Conference 2005 (EURAD 2005), Paris, France, 3–4 October 2005; Volume 2005, pp. 197–200. [Google Scholar]
- Durden, S.L.; van Zyl, J.J.; Zebker, H.A. The unpolarized component in polarimetric radar observations of forested areas. IEEE Trans. Geosci. Remote Sens. 1990, 28, 268–271. [Google Scholar] [CrossRef]
- Ainsworth, T.L.; Lee, J.S.; Schuler, D.L. Multi-frequency polarimetric SAR data analysis of ocean surface features. In Proceedings of the IGARSS 2000. IEEE 2000 International Geoscience and Remote Sensing Symposium. Taking the Pulse of the Planet: The Role of Remote Sensing in Managing the Environment. Proceedings (Cat. No.00CH37120), Honolulu, HI, USA, 24–28 July 2000; Volume 3, pp. 1113–1115. [Google Scholar]
- Henderson, F.M.; Lewis, A.J. Manual of Remote Sensing: Principles and Applications of Imaging Radar, 3rd ed.; John Wiley Sons: New York, NY, USA, 1998; 896p. [Google Scholar]
- Pope, K.O.; Rey-Benayas, J.M.; Paris, J.F. Radar remote sensing of forest and wetland ecosystems in the Central American tropics. Remote Sens. Environ. 1994, 48, 205–219. [Google Scholar] [CrossRef]
- Saatchi, S.S.; Dubayah, R.; Clark, D.; Chazdon, R.; Hollinger, D. Estimation of forest biomass change from fusion of radar and lidar measurements. In Seminário de Atualização em Sensoriamento Remoto E Sistemas de Informações Geográficas Aplicados À Engenharia Florestal, 9 (Sengef, 2010); Personal Communication; SENGEF: Curitiba, Brasil; Available online: http://www.slideshare.net/grssieee/estimation-of-forest-biomass (accessed on 13 May 2016).
- Nguyen, L.V.; Tateishi, R.; Nguyen, H.T.; Sharma, R.C.; To, T.T.; Le, S.M. Estimation of Tropical Forest Structural Characteristics Using ALOS-2 SAR Data. Adv. Remote Sens. 2016, 5, 131–144. [Google Scholar] [CrossRef]
- Hall, M. Correlation-based Feature Selection for Machine Learning. Ph.D. Thesis, University of Waikato, Hamilton, New Zealand, 1999. [Google Scholar]
- Romanski, P.; Kotthoff, L. FSelector: Selecting Attributes. R Package Version 0.31. 2015. Available online: https://CRAN.R-project.org/package=FSelector (accessed on 05 July 2017).
- Yang, Y.; Pedersen, J.O. A comparative study on feature selection in text categorization. In Proceedings of the Fourteenth International Conference on Machine Learning (ICML’97), Nashville, TN, USA, 8–12 July 1997; Morgan Kaufmann Publishers Inc.: San Francisco, CA, USA, 1997; pp. 412–420. [Google Scholar]
- Parimala, R.; Nallaswamy, R. A Study of Spam E-mail classification using Feature Selection package. Glob. J. Comput. Sci. Technol. 2011, 11, 45–54. [Google Scholar]
- Calcagno, V. Glmulti: Model Selection and Multimodel Inference Made Easy. R Package Version 1.0.7. 2013. Available online: https://CRAN.R-project.org/package=glmulti (accessed on 18 February 2016).
- Hastie, T.; Tibshirani, R.; Friedman, J. The Elements of Statistical Learning: Data Mining, Inference, and Prediction, 2nd ed.; Springer: Stanford, CA, USA, 2009; 745p. [Google Scholar]
- Lebrija-Trejos, E.; Bongers, F.; Pérez-García, E.A.; Meave, J.A. Successional Change and Resilience of a Very Dry Tropical Deciduous Forest Following Shifting Agriculture. Biotropica 2008, 40, 422–431. [Google Scholar] [CrossRef]
- Foody, G.M.; Green, R.M.; Lucas, R.M.; Curran, P.J.; Honzak, M.; Do Amaral, I. Observations on the relationship between SIR-C radar backscatter and the biomass of regenerating tropical forests. Int. J. Remote Sens. 1997, 18, 687–694. [Google Scholar] [CrossRef]
- Kuplich, T.M.; Curran, P.J.; Atkinson, P.M. Relating SAR image texture to the biomass of regenerating tropical forests. Int. J. Remote Sens. 2005, 26, 4829–4854. [Google Scholar] [CrossRef]
- Sai Bharadwaj, P.; Kumar, S.; Kushwaha, S.P.S.; Bijker, W. Polarimetric scattering model for estimation of above ground biomass of multilayer vegetation using ALOS-PALSAR quad-pol data. Phys. Chem. Earth Parts A/B/C 2015, 83–84, 187–195. [Google Scholar] [CrossRef]
- Jakovac, C.C.; Peña-Claros, M.; Kuyper, T.W.; Bongers, F. Loss of secondary-forest resilience by land-use intensification in the Amazon. J. Ecol. 2015, 103, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Poorter, L.; Bongers, F.; Aide, T.M.; Almeyda Zambrano, A.M.; Balvanera, P.; Becknell, J.M.; Boukili, V.; Brancalion, P.H.S.; Broadbent, E.N.; Chazdon, R.L.; et al. Biomass resilience of Neotropical secondary forests. Nature 2016, 530, 211–214. [Google Scholar] [CrossRef]
- Ahmed, R.; Siqueira, P.; Hensley, S. Analyzing the uncertainty of biomass estimates from l-band radar backscatter over the harvard and howland forests. IEEE Trans. Geosci. Remote Sens. 2014, 52, 3568–3586. [Google Scholar] [CrossRef]
- Joshi, N.; Mitchard, E.; Schumacher, J.; Johannsen, V.; Saatchi, S.; Fensholt, R. L-Band SAR Backscatter Related to Forest Cover, Height and Aboveground Biomass at Multiple Spatial Scales across Denmark. Remote Sens. 2015, 7, 4442–4472. [Google Scholar] [CrossRef]









| N | Plot Size (m) | DBH min (cm) | Age (Years) | Inventory Date (Year) | Succession | Area (ha) | Reference |
|---|---|---|---|---|---|---|---|
| 16 | 10 × 100 | 5 | 16–28 | 2015 | Advanced (Adv) | 1.60 | Our study |
| 16 | 20 × 100 | 10 | 16–28 | 2015 | Advanced (Adv) | 3.20 | Our study |
| 16 | 60 × 100 | 20 | 16–28 | 2015 | Advanced (Adv) | 9.60 | Our study |
| 14 | 20 × 50 | 5 | 1–7 | 2012/13 | Initial (ISS) | 1.75 | Silva et al. [64] |
| 9 | 20 × 50 | 5 | 7–16 | 2012/13 | Intermediate (IntSS) | 2.40 | Silva et al. [64] |
| 3 | 25 × 100 | 10 | >16 | 2012/13 | Advanced (Adv) | 1.35 | Silva et al. [64] |
| Biophysical Variables | Asymptote | Units | References |
|---|---|---|---|
| Mean diameter (DBH) | 23.1 | cm | [25,58,79,80,81,82,83,84] |
| Mean total tree height (Ht) | 25.0 | m | [25,79,82,85,86] |
| Basal area (G) | 24.4 | m2 ha−1 | [25,79,82,85,87,88,89,90,91] |
| Species richness (S) | 133.00 | sp ha−1 | [60] |
| Tree density (N) | 433.00 | individuals ha−1 | [25,60,80,82,88,90,91] |
| Aboveground biomass (ABG) | 314.00 | Mg ha−1 | [25,60,61,79,92] |
| Date | Sensor | Date | Sensor | Date | Sensor |
|---|---|---|---|---|---|
| 24 August 1984 | TM | 20 October 1993 | TM | 29 August 2003 | TM |
| 26 July 1985 | TM | 10 October 1995 | TM | 1 July 2005 | TM |
| 29 July 1986 | TM | 25 August 1996 | TM | 5 August 2006 | TM |
| 16 July 1987 | TM | 27 July 1997 | TM | 21 June 2007 | TM |
| 3 August 1988 | TM | 15 August 1998 | TM | 30 November 2008 | TM |
| 22 August 1989 | TM | 3 September 1999 | TM | 12 July 2009 | TM |
| 9 August 1990 | TM | 5 September 2000 | TM | 29 June 2010 | TM |
| 11 July 1991 | TM | 16 September 2001 | ETM+ |
| Input Matrix | N°Attr. | Polarimetric Attributes | Reference |
|---|---|---|---|
| C3 | 9 | I_C11, I_C12imag, I_C12real, I_C13imag, I_C13real, I_C22, I_C23imag, I_C23real, I_C33 | [104] |
| C3 | 3 | Freeman_Dbl, Freeman_Odd, Freeman_Vol | [105] |
| C3 | 2 | ρhh-vv, |ρhh-vv| | [104] |
| C3 | 3 | Neumann_mDelta, Neumann_phDelta, Neumann_tau | [106] |
| C3 | 3 | VanZyl_Dbl, VanZyl_Odd, VanZyl_Vol | [107] |
| C3 | 4 | Yamaguchi_Dbl, Yamaguchi_Hlx, Yamaguchi_Odd, Yamaguchi_Vol | [108] |
| C3 | 4 | Bhattacharya_Dbl, Bhattacharya_Hlx, Bhattacharya_Odd, Bhattacharya_Vol | [109] |
| C3 | 5 | MCSM_Dbl, MCSM_DblHlx, MCSM_Odd, MCSM_Vol, MCSM_Wire | [110] |
| C3 | 4 | Singh_Dbl, Singh_Hlx, Singh_Odd, Singh_Vol | [111] |
| S, T3 | 16 | TSVM_alpha_s, TSVM_alpha_s1, TSVM_alpha_s2, TSVM_alpha_s3, TSVM_phi_s, TSVM_phi_s1, TSVM_phi_s2, TSVM_phi_s3, TSVM_psi_s, TSVM_psi_s1, TSVM_psi_s2, TSVM_psi_s3, TSVM_tau_s, TSVM_tau_s1, TSVM_tau_s2, TSVM_tau_s3 | [112] |
| T3 | 9 | T11, T12imag, T12real, T13imag, T13real, T22, T23imag, T23real, T33 | [104] |
| T3 | 15 | A, H, α, β, λ, γ, δ, p1, p2, p3, HA, H_A, λ1, λ2, λ3 | [113] |
| T3 | 9 | T11_H, T12imag_H, T12real_H, T13imag_H, T13real_H, T22_H, T23imag_H, T23real_H, T33_H | [114] |
| T3 | 9 | T11_C, T12imag_C, T12real_C, T13imag_C, T13real_C, T22_C, T23imag_C, T23real_C, T33_C | [115] |
| T3 | 9 | T11_B, T12imag_B, T12real_B, T13imag_B, T13real_B, T22_B, T23imag_B, T23real_B, T33_B | [116] |
| T3 | 6 | SE, SE_norm, SE_I, SE_I_norm, SE_P, SE_P_norm | [117] |
| T3 | 4 | SERD, SERD_norm, DERD, DERD_norm | [118] |
| T3 | 1 | PH—pedestal height | [119] |
| T3 | 1 | PF—polarization fraction | [120] |
| T3 | 1 | RVI—radar vegetation index | [107] |
| C3 | 3 | VSI—vegetation scattering index, BMI—biomass index, CSI—canopy structure index | [121] |
| C3 | 1 | RFDI—radar forest degradation index | [122] |
| C3 | 1 | Span (Total Power) | [104] |
| C3 | 2 | Rpp—parallel polarization ratio, Rcp—cross-polarization ratio | [123] |
| C3 | 1 | Forest | [124] |
| Plot | n | Age * | FC | PALU | DBH | Ht | G | S | N | AGB |
|---|---|---|---|---|---|---|---|---|---|---|
| (Plots) | (Years) | (x) | (Years) | (cm) | (m) | (m2 ha−1) | (sp ha−1) | (ind ha−1) | (Mg ha−1) | |
| ISS | 14 | 3.3(1.1) | 2.8(0.5) | 3.1(1.7) | 11.9(2.8) | 13.0(1.4) | 14.0(5.6) | 29(9) | 841(145) | 83.2(42.2) |
| IntSS | 9 | 9.2(2.3) | 2.2(1.1) | 2.8(1.9) | 12.9(1.1) | 13.5(0.5) | 16.7(2.3) | 31(4) | 785(57) | 99.3(17.8) |
| Adv | 19 | 20.2(2.6) | 1.9(0.9) | 1.4(0.5) | 17.0(2.3) | 15.0(1.1) | 24.9(3.8) | 49(7) | 1274(424) | 151.5(30.4) |
| Mean | 42 † | 12.2(7.9) | 2.3(1.1) | 2.3(1.5) | 14.4(3.3) | 14(1.5) | 19.5(6.5) | 39(12) | 1025(373) | 117.2(45.2) |
| AIC | R2 Validation | RMSE (Mg ha−1) | RMSE ISS | RMSE IntSS | RMSE Adv | RMSE PALU 1–2 years | RMSE PALU 3–4 years | RMSE PALU >4 years | Bias (Mg ha−1) | |
|---|---|---|---|---|---|---|---|---|---|---|
| NL | 417.1 | 0.28 | 39.7 | 50.9 | 34.6 | 38.2 | 45.4 | 33.2 | 31.3 | 0.3 |
| MLR | 408.1 | 0.37 | 38.7 | 52.6 | 37.7 | 33.8 | 37.6 | 43.6 | 21.8 | 2.1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cassol, H.L.G.; Carreiras, J.M.d.B.; Moraes, E.C.; Aragão, L.E.O.e.C.d.; Silva, C.V.d.J.; Quegan, S.; Shimabukuro, Y.E. Retrieving Secondary Forest Aboveground Biomass from Polarimetric ALOS-2 PALSAR-2 Data in the Brazilian Amazon. Remote Sens. 2019, 11, 59. https://doi.org/10.3390/rs11010059
Cassol HLG, Carreiras JMdB, Moraes EC, Aragão LEOeCd, Silva CVdJ, Quegan S, Shimabukuro YE. Retrieving Secondary Forest Aboveground Biomass from Polarimetric ALOS-2 PALSAR-2 Data in the Brazilian Amazon. Remote Sensing. 2019; 11(1):59. https://doi.org/10.3390/rs11010059
Chicago/Turabian StyleCassol, Henrique Luis Godinho, João Manuel de Brito Carreiras, Elisabete Caria Moraes, Luiz Eduardo Oliveira e Cruz de Aragão, Camila Valéria de Jesus Silva, Shaun Quegan, and Yosio Edemir Shimabukuro. 2019. "Retrieving Secondary Forest Aboveground Biomass from Polarimetric ALOS-2 PALSAR-2 Data in the Brazilian Amazon" Remote Sensing 11, no. 1: 59. https://doi.org/10.3390/rs11010059