1. Introduction
Capturing the three-dimensional (3D) structure and the spatial heterogeneity of vegetation canopies is crucial for characterizing ecosystems mass and energy fluxes. 3D leaf distribution has become a critical parameter for modelling radiative transfers [
1] and for eco-physiological models incorporating photosynthesis and transpiration processes [
2,
3]. Moreover, the vegetative element distribution is an important characteristic of forest stand conditions, which affects carbon cycling, wildlife habitat [
4], or disturbances and stresses such as wildfire [
5]. It is hence a key parameter to monitor.
Foliage density or Leaf Area Density (
LAD, m
2/m
3), defined as the one-sided area of leaves per unit volume [
6], can theoretically be retrieved from direct measurement of the surface area of leafy elements in reference volumes of interest. However, such an approach is not practical and remains volume limited, as it is extremely time consuming and has obvious destructive consequences on vegetation. As a result, foliage area density is often quantified through a vertical integration, namely the Leaf Area Index (
LAI), which can be indirectly measured using ground-based methods [
6,
7,
8]. However, these methods do not provide the 3D foliage distribution and require empirical corrections to account for clumping [
9].
With a dense and regular laser sampling, Terrestrial LiDAR (Light Detection and Ranging) scanning, hereafter referred to as TLS, has a great potential to characterize vegetation 3D structure. TLS technology relies on a high frequency emission/reception of low divergent laser beams with registration of the 3D coordinates associated to each laser hit. As a result, TLS acquires dense 3D point clouds, providing a high-resolution representation of forest canopies. Several attempts to retrieve foliage attributes from TLS have led to encouraging results, based either on the measurement of gap fractions [
10,
11] or on the estimation of
LAD in voxels [
12,
13,
14,
15]. Gap fraction approaches generally assume homogeneous layers to estimate effective LAD profile and use a factor to correct for clumping effect [
11,
15]. In contrast to gap fraction approaches, the voxel-based methods, which are studied in the present paper, explicitly account for vegetation heterogeneity (i.e., clumping effect) at scales larger than voxel size.
These voxel methods use various metrics to estimate the
LAD, based on different estimators of the attenuation coefficient, which is the rate at which vegetation attenuates beam transmission. The Relative Density Index (
RDI) is the ratio of number of hits inside a voxel to the number of beams entering the voxel [
16] and is an empirical measurement of vegetation absorbance [
15]. Voxel-based methods rely on different functional forms of the RDI or other statistics such as the mean path length (mean distance potentially explored by beams within a voxel assuming no element intercepted them) and the mean free path length (mean distance actually explored by beams within a voxel) [
15]. These indices can be readily applied or combined with field measurements through a calibration phase [
14]. The voxel-based canopy profiling method [
17] was developed for very small voxel sizes (below 10 cm) and uses ratios of full
versus empty voxels to compute RDI. For larger voxel sizes, intra-voxel LAD estimators can be computed, using the relative density index [
14,
16], the contact frequency [
18], the modified contact frequency [
12] and the Beer-Lambert law [
13,
19,
20]. Intra-voxel estimators were recently compared using a theoretical framework and numerical experiments [
15].
A key characteristic of vegetation when retrieving
LAD in these formulations is the leaf orientation. Indeed, the estimated attenuation coefficient is the projected leaf area density of the voxel in the plane perpendicular to the direction of emission of the laser beam. This projection ratio depends on leaf orientation and is accounted for with the
G factor [
12]. The value attributed to
G for random leaf orientation is 0.5 (i.e., the projected area is half the one-sided leaf area). Determining the exact
G value in voxels with TLS is challenging [
13] but is now possible for large leaves [
20]. Regarding small elements, no method has gained a consensus yet but it has been shown that branches (and by extension leaves) orientation has a limited impact on the quality of results, even for planophile and erectophile distributions [
13,
14,
21], supporting the validity of an assumption of
G = 0.5. Another critical aspect is the differentiation between leaf and wood return, as the final objective of these methods is to determine the
LAD and not the
PAD (Plant Area Density, which includes both leaf and wood surface areas).
The theoretical bases supporting these methods generally assume that vegetation elements are small and randomly distributed within a voxel [
12,
15]. As leaf clumps are surrounded by empty volumes, voxels should be small enough to discretize large gaps between branches [
22]. Indeed, aggregating empty spaces and leaf clumps in a single large voxel leads to a systematic underestimation of predicted leaf area, as a consequence of Jensen’s inequality [
23,
24], as suggested in Reference [
15]. However, smaller voxels do not necessarily increase the accuracy of the estimation. Indeed, the voxels should be large enough with respect to the size of vegetation elements, otherwise leading to an overestimation of the
LAD [
22]. Also, smaller voxels cause a sharp rise in the variability of
LAD estimates, as it decreases the number of beams and the number of vegetation elements in voxels [
15]. It should be noted, however, that theoretical corrections can be implemented to account for element size and number of beams [
15].
In addition to biases inherent to vegetation structure, the instrument can introduce deviations from actual
LAD values, mostly as a consequence of the finite cross-sectional diameter of the scanner beam, in particular with single return phase shift scanners. A laser beam can indeed partly hit a leaf, while a remaining—not intercepted—fraction of laser energy continues its path before possibly interacting with another object located in the background. In this case, a single return phase-shift scanner registers a unique point which can be anywhere between the two objects, leading to a hit misplacement (mixed points, [
25]). As a result, different backgrounds (vegetated vs. open) may affect any partial hit of vegetation elements and in turn,
LAD predictions. In addition, as the footprint (cross-sectional area) of a laser beam increases with distance to scanner (because of beam divergence), the probability to hit vegetation elements increases, which induces an increase in the
LAD estimation with distance to scanner. Such bias can be corrected by weighting estimated
RDI, with distance-dependent calibrated function of return intensity [
12] but to date, this approach was only applied at fixed distance only (scanning individual plants at constant distance to the scanner). Finally, the limited number of beams effectively reaching a voxel is a last source of bias and uncertainty in
TLS measurement [
15]. Low beam number arises from both increasing distance to scanner and vegetation occlusion.
Hence, the three sources of bias –theoretical, vegetation and instrument- of LAD estimates are sensitive to both voxel size and distance to scanner. Indeed, variations in voxel size affect the number of beams, the relative size of vegetation element (compared to voxel size) and the discretization of vegetation structure (i.e., how clumps and gaps are discretized). Also, increased distance to scanner decreases the number of beams, increases the probability of hit and decreases returned energy (so detection ability). Among the different sources of bias, some of them can be theoretically corrected, as suggested in Reference [
15].
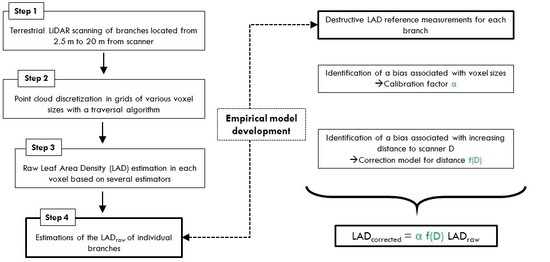
This study aims to (i) disentangle the different sources of bias and to evaluate their magnitude, (ii) focus on the specific biases associated with voxel size and distance to scanner and (ii) propose generic calibration models to retrieve correct LAD estimates from TLS measurements. To achieve these objectives, we worked on tree branches in a controlled environment with negligible occlusion. Accurate destructive LAD reference values were available for 46 branches. We first investigated the performances of five different voxel-based methods for estimating the LAD of vegetation samples. In particular, these estimators were tested on three species with distinct foliage shapes, sizes and clumping patterns. Secondly, we investigated the sensitivity of the predictions to discretization (voxel-size effect) and distance to scanner (distance effect). These analyses were carried out with two scanners (one phase shift and one time of flight scanner), which expands the scope of our results.
3. Results
Results are organized in four subsections, as described in
Section 2.5. First, we compare the raw predictions arising from the different estimators. Second, we compare the raw
LAD to the actual (reference)
LAD and investigate how the relationship was affected by the voxel size, for short distance measurements (distance to scanner of 2.5 m). Third, we analyse the sensitivity of predictions to the distance to scanner. Finally, we present some calibration functions correcting for both voxel size and distance biases, as well as metrics to evaluate the performance of the corresponding models.
3.1. Comparison between Predictions of the Different Estimates
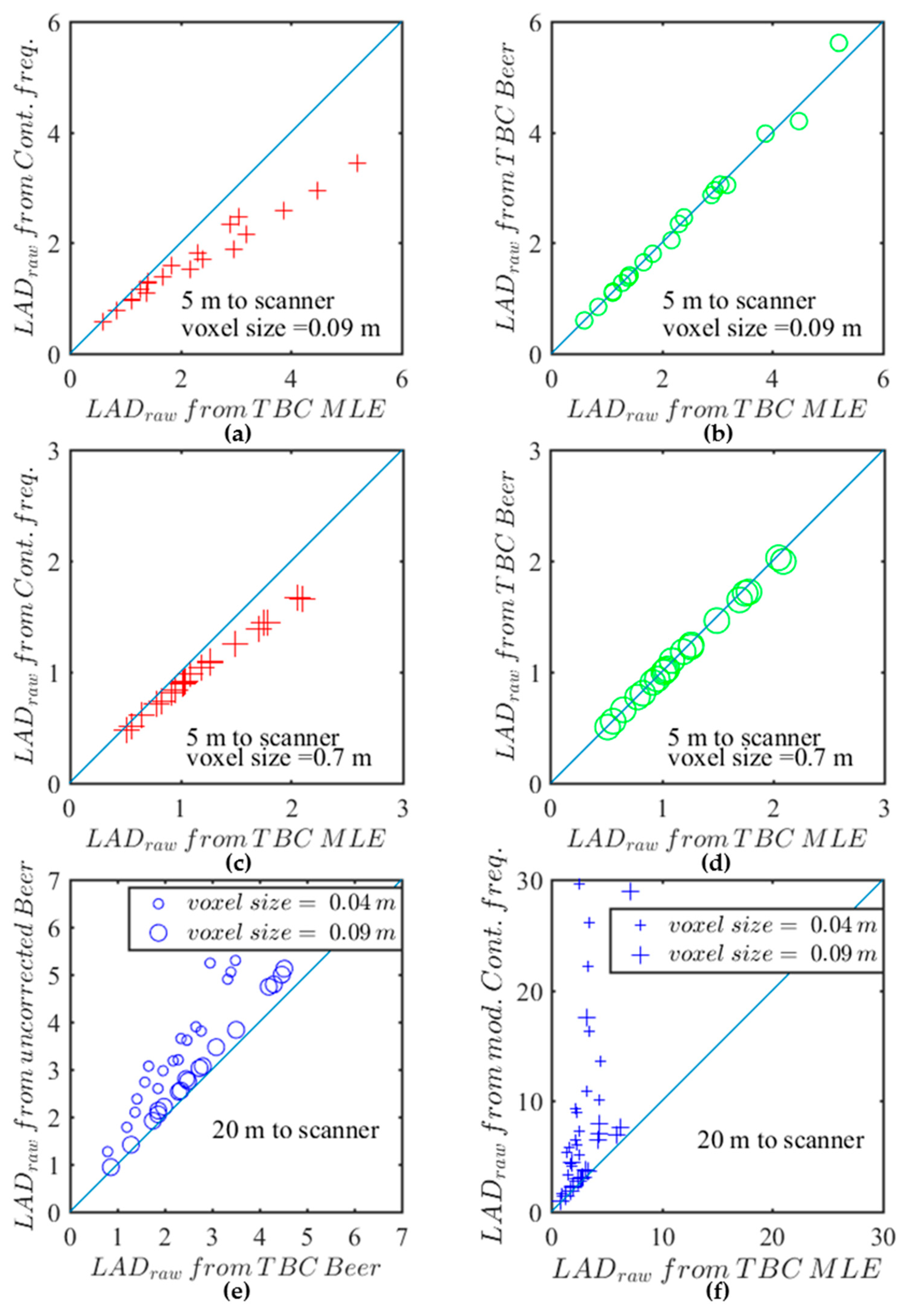
First, both TBC estimators (from Beer-Lambert law or MLE) led to very similar predictions. This is illustrated in
Figure 3a,b which show both predictions for
Quercus pubescens, at a distance to scanner of 5 m and for respectively Beer-Lambert law or MLE at 9 cm and 70 cm voxel sizes, revealing that they were in close agreement (green circles close to the 1:1 line). Second,
Figure 3c,d show that predictions arising from the Contact Frequency were significantly lower than those from the bias-corrected MLE (red crosses). This difference can be explained by the systematic underestimation arising from the Contact Frequency method that was previously mentioned. Third, we found that there may be significant differences between predictions arising from TBC and corresponding uncorrected estimators when the vegetation is far from the scanner (
Figure 3e,f). TBC estimators were systematically lower, especially for small voxel sizes. For example, the
computed with the TBC Beer-Lambert law were roughly 40% lower than the
computed with the uncorrected Beer-Lambert law for a 4 cm voxel size, whereas the estimates based on the TBC MLE were up to 100% lower than those based on the Modified Contact Frequency, even for a 9 cm voxel size. These differences arose from shifts in distributions of free path lengths and effective free path lengths (
Appendix D).
More generally, for distances lower than 10 m and voxel sizes larger than 9 cm, we found that bias corrections were negligible, as predictions of the corrected and uncorrected estimators were similar. One noticeable exception was the estimator derived from the Contact Frequency approach, which systematically led to lower estimates than the other estimators.
Hence, when implemented, the theoretical bias corrections induced a systematic decrease in predictions of . The differences arising from the corrections were significant in the most extreme cases (voxels smaller than 5 cm and distances larger than 15 m), especially for the TBC MLE, for which the range of predictions was close to the range of reference values (between 0 and 4 m2/m3), whereas the Modified Contact Frequency led to value as high as 30 m2/m3.
For brevity, most results in the following sections are limited to predictions arising from bias-corrected MLE and the contact frequency, which exhibited the most significant differences.
When computing the correlations between raw estimates and reference measurements of
LAD, we found similar correlation coefficients for most raw estimators, as they were almost linearly related. Performances will be analysed in detail in the next subsections. We found, however, that the MCF for small voxel sizes exhibited significantly lower correlation coefficients than the others, which was explained by the overestimations observed in
Figure 3f (the range of reference measurement is 0–4 m
2/m
3), highlighting the importance of bias correction for this estimator.
3.2. Influence of the Voxel Size on Short Distance Measurements and Subsequent Calibration
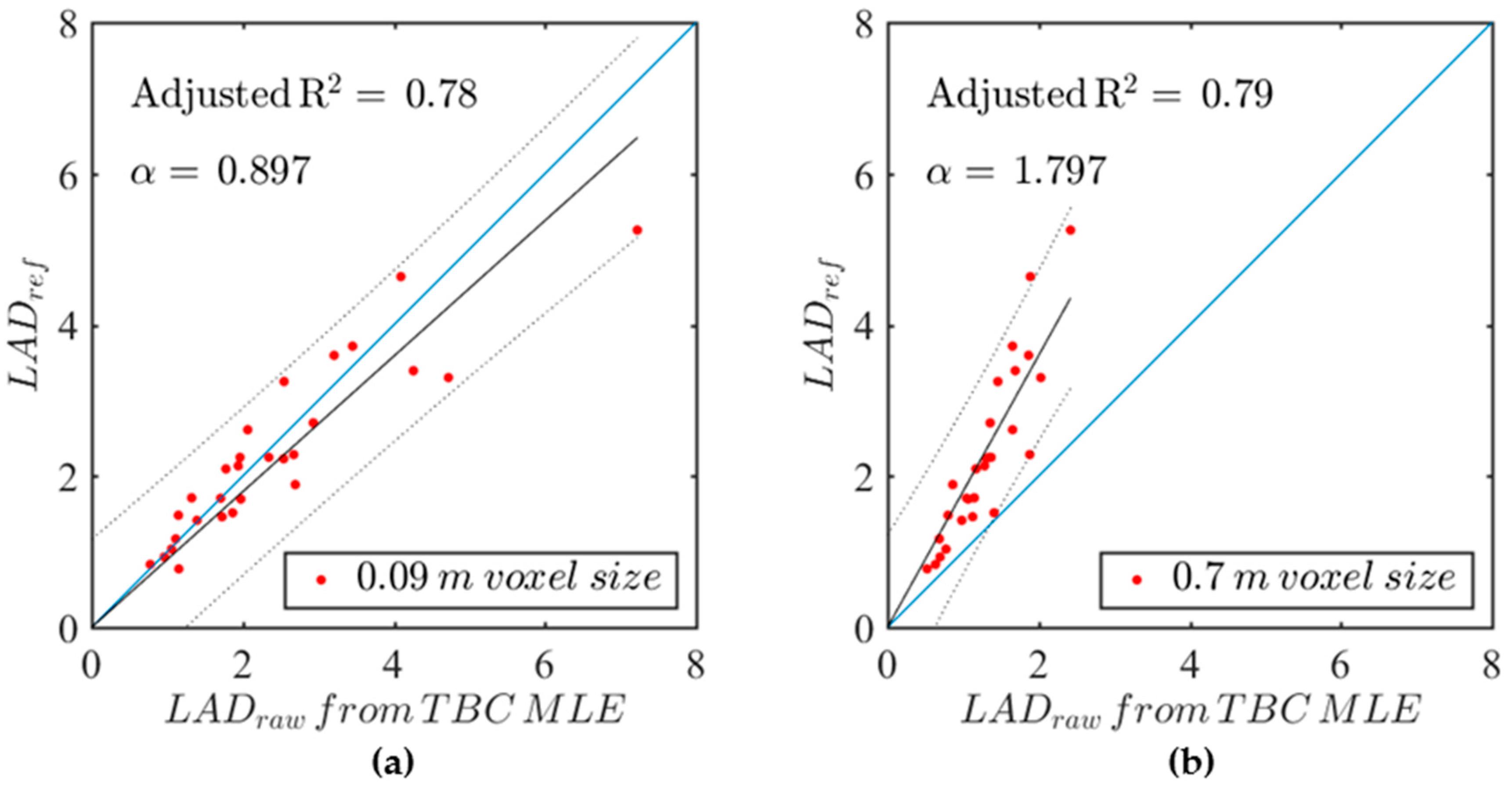
Figure 4 shows some example correlations between raw leaf area densities derived from the FARO data using the TBC MLE estimator at
d = 2.5 m and reference leaf area densities for
Q. ilex. Subplots (a) and (b) correspond to 9 cm and 70 cm voxel sizes, respectively. A linear fit with a null intercept is reasonable for both voxel sizes, showing that reference and raw LAD were proportional. A regression coefficient
smaller or larger than 1 respectively corresponds to over and underestimation, whereas
equal to 1 corresponds to an unbiased estimator. We found that the 0.09 m voxel size exhibited a small positive bias, whereas the 0.7 m voxel size exhibited a strong negative bias.
Similar models were fitted for all studied voxel sizes and species, which were the only factors significantly affecting the relationship between raw and reference
LAD (
Appendix C.2). As the coefficient
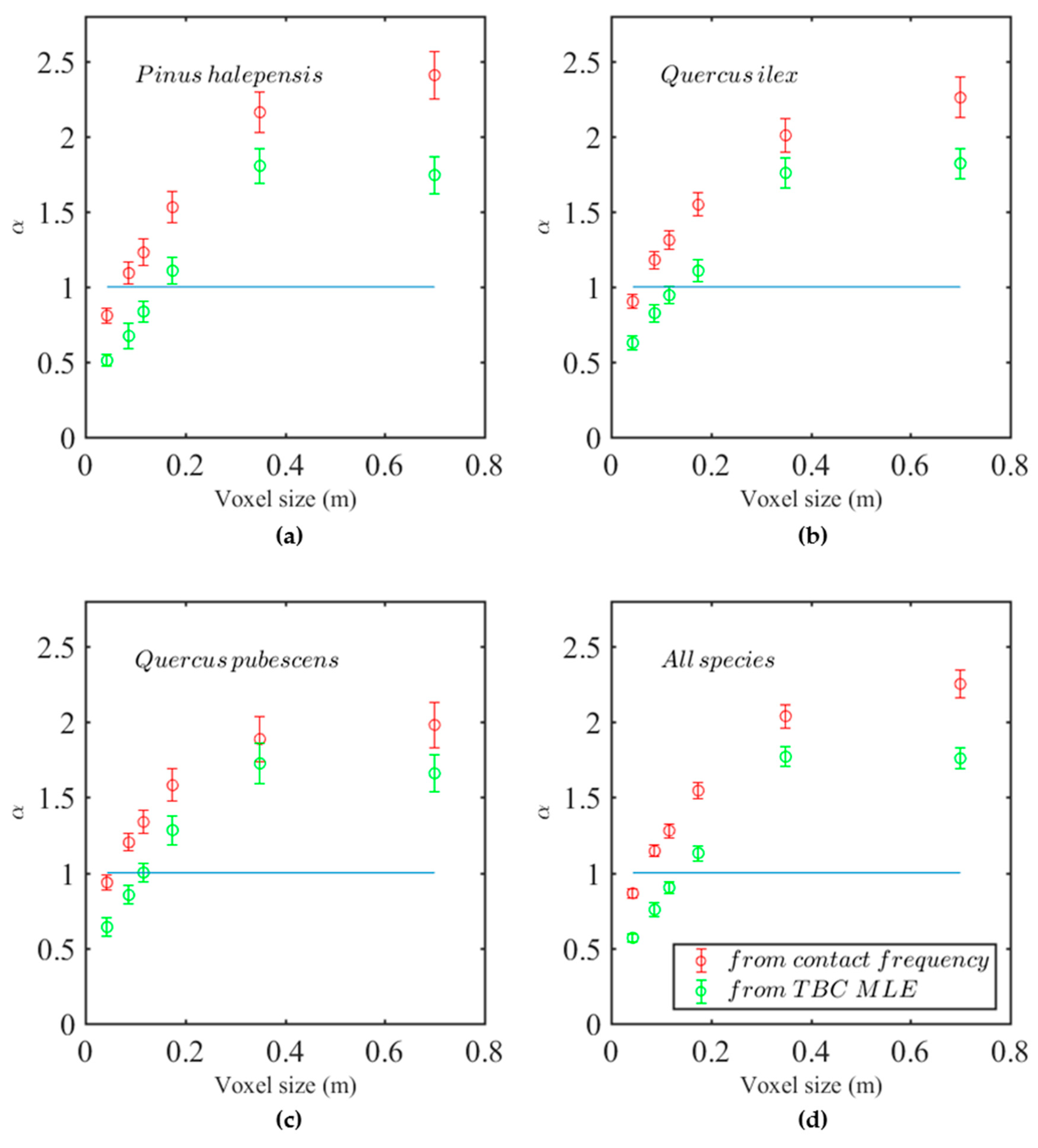
often significantly differed from 1, it is later referred to as “calibration coefficient.” The calibration coefficients
are shown in
Figure 5 for raw estimators deriving from both Contact Frequency (in red) and TBC MLE (in green). In all cases, the predicted calibration coefficient increased with voxel size until a saturation was reached for voxels larger than 0.35 m, corresponding to a decrease in raw predictions compared to references as voxel size increases.
Figure 5a–c shows the calibration coefficients for the three species, whereas all species were combined in
Figure 5d. Although statistically significant (
Appendix C.2), the differences between species were quantitatively marginal, as the different species exhibited very similar sensitivity to voxel size.
The voxel size was hence clearly the main effect, with a strong increase in calibration coefficient with voxel size for both Contact frequency and TBC MLE.
3.3. Influence of the Distance to Scanner and Subsequent Calibration
Figure 6 illustrates the influence of the distance to scanner on the raw
LAD, for the same case as
Figure 4a (i.e.,
Q. ilex -0.09 m voxel size), the differently coloured dots corresponding to the different distances to scanner. The coloured lines show the slopes of the relationship obtained at the different distances. Obviously, a strong effect of the distance of measurement is observed with the FARO scanner (up to +100% of 2.5 m value for some of the branches), leading to a strong overestimation of the
LAD at greater distances.
The statistical analysis (
Appendix C.3) suggested that this effect of distance to scanner was much lower for the RIEGL than for the FARO scanner and that it was affected by factors such as voxel size and species.
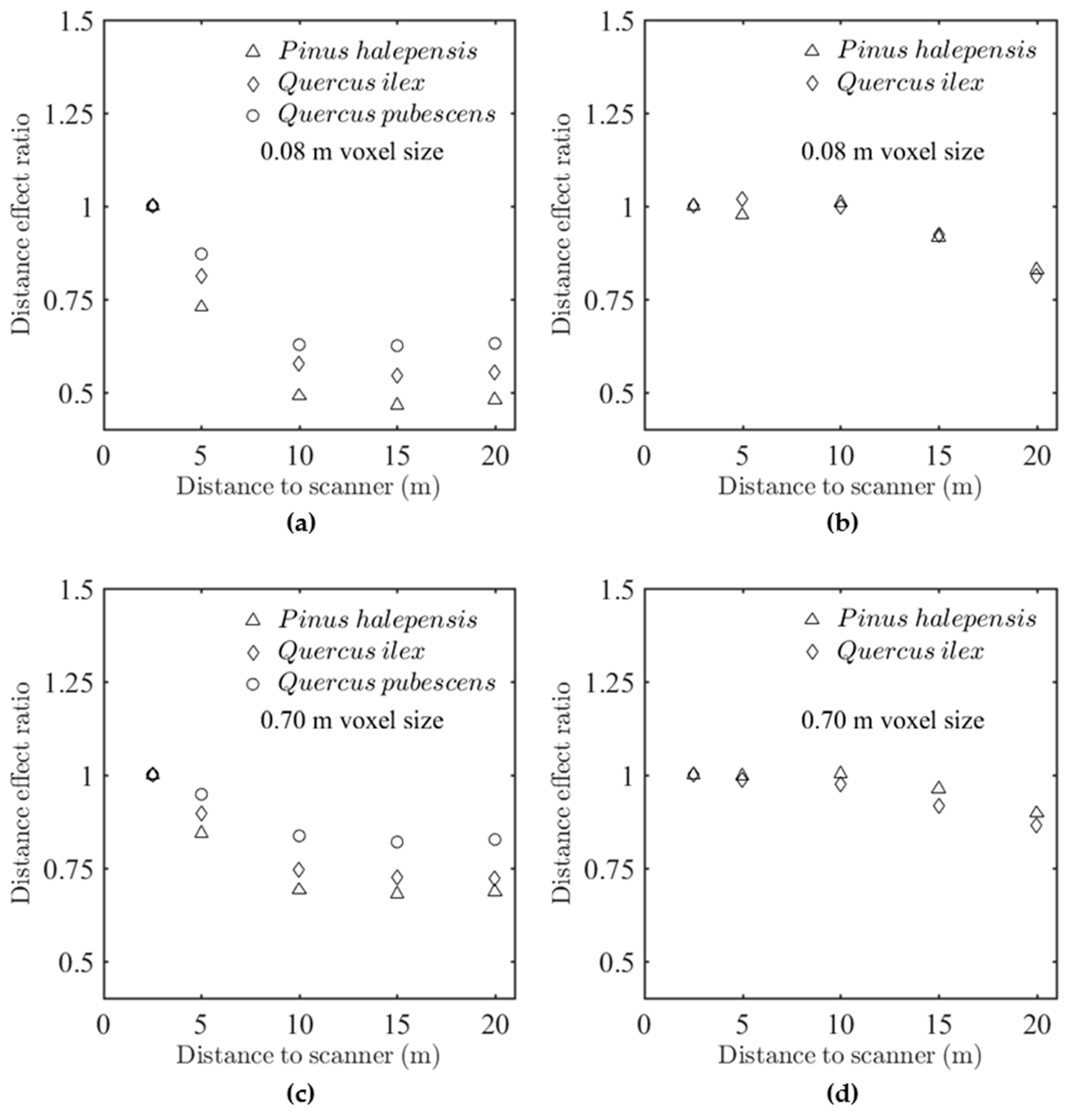
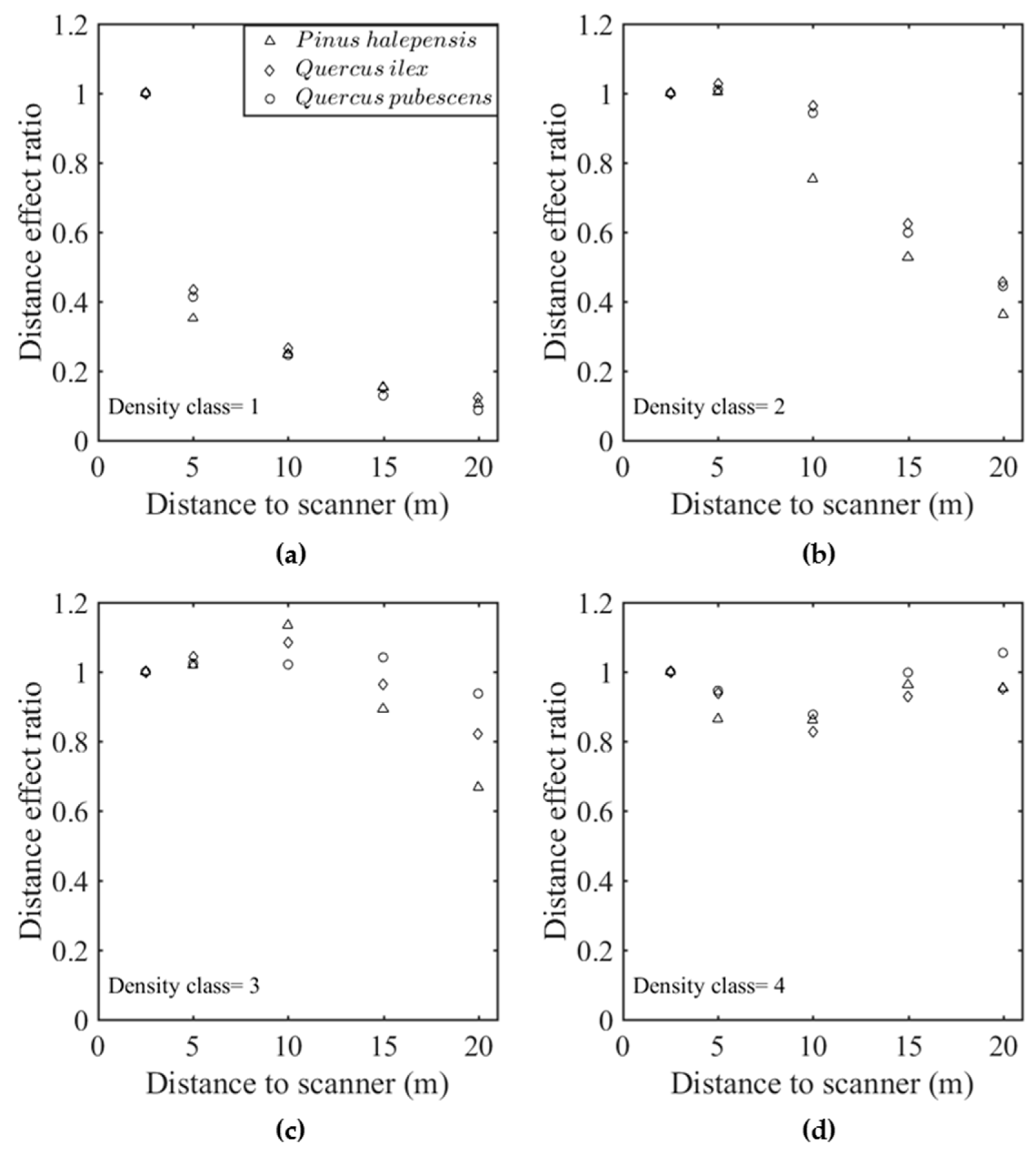
Figure 7 shows the “distance effect” ratio, which was defined as the ratio between 2.5 m predictions and the predictions at a given distance
d. This ratio was also the calibration factor by which predictions should be multiplied to correct the distance bias (Equation (7)). Subplots (a) and (c) correspond to FARO scanner for two voxel sizes (0.09 and 0.7m), whereas subplots (b) and (d) correspond to the RIEGL scanner for the same voxel sizes. The comparison between the two scanners confirmed that the distance effect was much weaker with the RIEGL than with the FARO system, as suggested in the statistical analysis. However,
Figure 7b,d suggest that even the RIEGL system exhibited a small positive bias beyond 10 m, especially for small voxel sizes. For the FARO scanner, the increasing positive bias suggested from
Figure 6 was confirmed for all species and the two resolutions. The distance effect thus followed an exponential attenuation for larger voxel sizes. We can observe that
Q. pubescens and
P. halepensis were the less and the most sensitive species, respectively. The differences between species, although significant (
Appendix C.3), were of limited magnitude. The differences induced by voxel size were, much more important, as suggested by the comparison between distance effects in subplot (a) and (c).
Similar trends were observed with the other estimators, with the exception of the estimator derived from the Contact Frequency for which the magnitude of distance effect was slightly smaller.
An in-depth analysis, reported in
Appendix E, shows that the bias associated with distance decreased when the vegetation density increased at voxel scale, since the distance effect on estimated
LAD was stronger when elements were sparse in a voxel. Such an effect could have been accounted for in the distance model but it would have induced a complex formulation, as the bias correction would have been a function of the overall prediction. This effect was thus neglected in the overall calibration presented in the next subsection.
3.4. Example Calibrated Estimators
In this subsection, we present a selection of calibrated models
developed to correct the raw estimators according to Equations (7)–(9). The models presented in
Table 3 were developed for both scanners and two voxel sizes (9 and 70 cm) to correct raw estimates obtained using contact frequency method, TBC Beer’s law and TBC MLE approach. The resulting calibrated
LAD values, referred to as
, included a correction for distance effect when computed for the FARO scanner, whereas this effect was neglected when processing data acquired with the RIEGL scanner (i.e.,
). The mean absolute percentage errors (MAPE) of the models were systematically smaller with the RIEGL instrument than with the FARO scanner. For the RIEGL scanner, the errors were found to be generally higher for
P. halepensis than for
Q. ilex.
The calibrated models arising from the most sophisticated attenuation coefficient estimators ( and ) exhibited slightly lower errors than the model deriving from the basic contact frequency for the largest voxel size (70 cm) but differences were generally small. When shifting to the small voxel size (9 cm), the errors of the FARO scanner increased for estimators derived from and and remained stable for the contact frequency (CF). Results were more contrasted with the RIEGL scanner, since errors were stable for Q. ilex but increased with P. halepensis when decreasing voxel size.
Overall, the most sophisticated estimators did not perform better than the basic contact frequency (with the exception of the
Q. ilex with the RIEGL scanner and the small voxel). This result, which may seem counterintuitive, could be explained by the fact that unbiased estimators often exhibit a larger variability than simpler biased estimators [
15], resulting in some cases to a lower residual error at the scale of individual measurements (bias-variance trade-off). This crucial point is discussed in the next section.
Fitting the models on subsets corresponding to species and/or scanners enabled to reduce MAPE values by around 5%. However, corresponding models required a much higher number of coefficients and were thus not considered for further analysis.
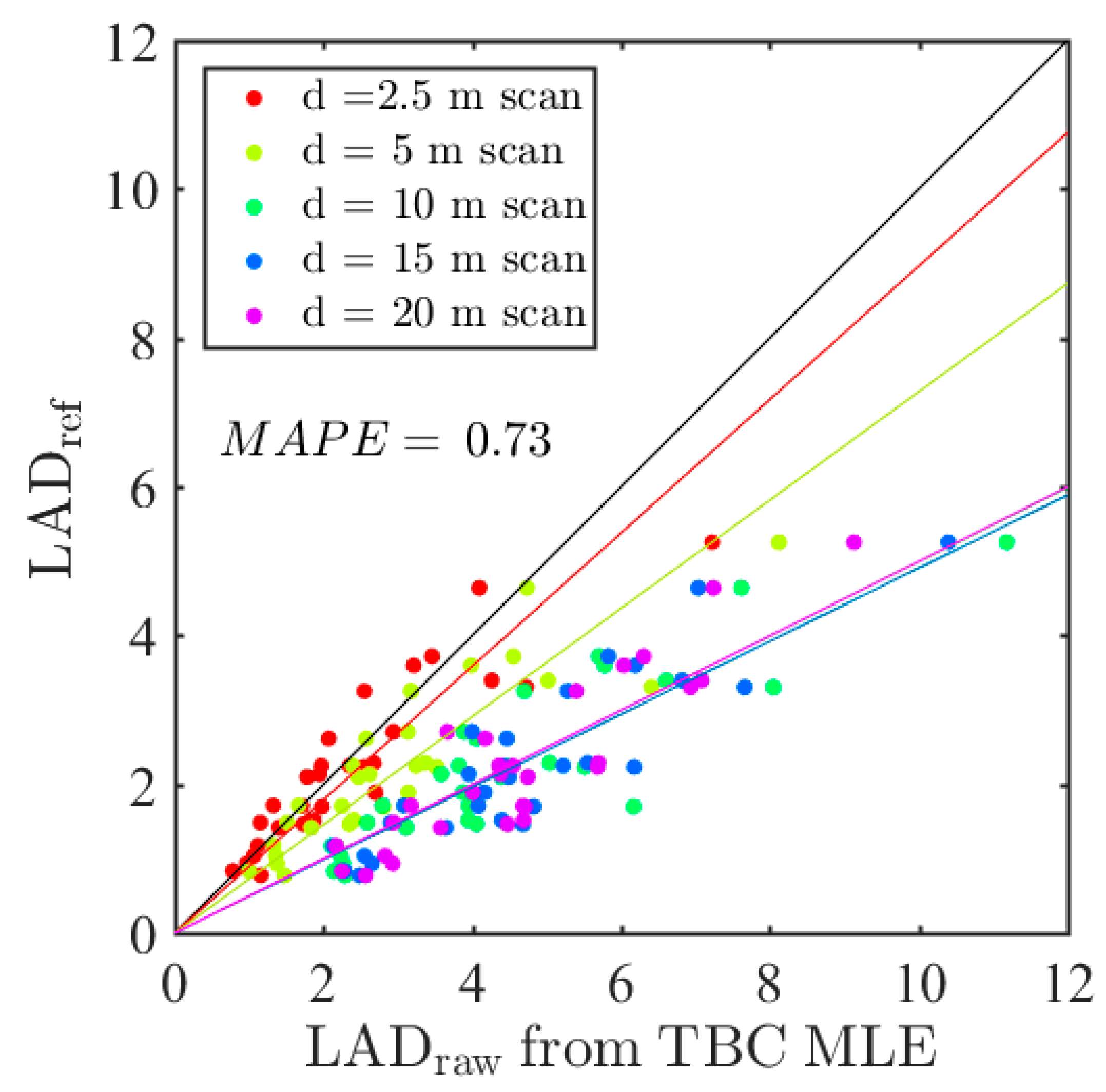
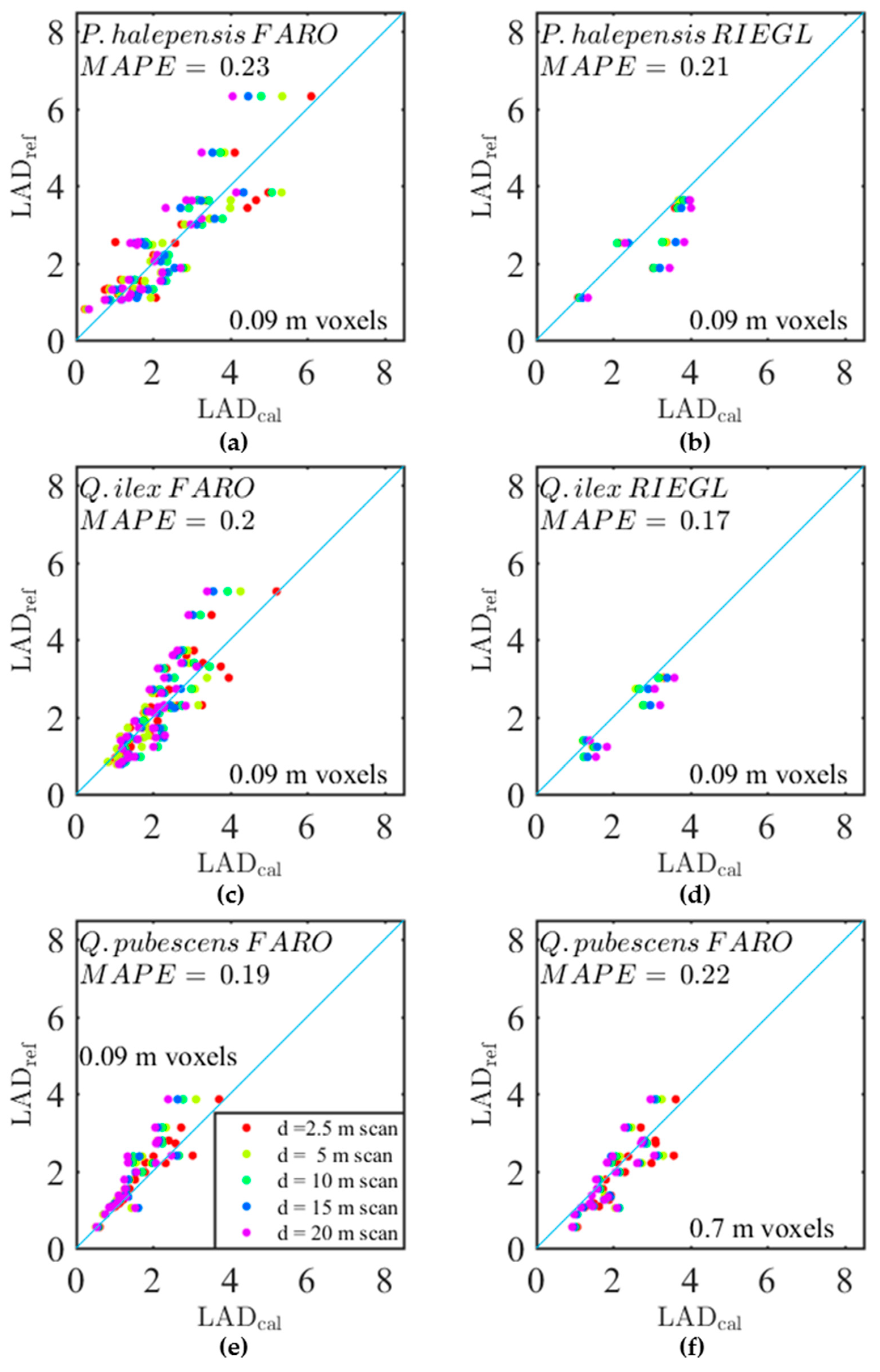
Figure 8 shows
LAD predictions after calibration (
) for some of the models presented in
Table 3. Residual biases of calibrated estimates were low for the FARO instrument, whatever the species or the resolution, with the exception of the branches with highest densities, which were generally underestimated. This underestimation was explained by the fact that the correction for the distance effect was too high in dense branches, because the sensitivity to voxel density of the distance effect was neglected in the calibration model, as explained in
Section 3.3. The RIEGL instrument appeared to be more prone to overestimations. This is consistent with the fact that no correction was applied for the positive bias associated with distance in the case of the RIEGL instrument.
4. Discussion
The present work aimed at providing new insights regarding the measurement of leaf area with TLS, from a comparison with destructive laboratory measurements. Our experimental set-up was designed to investigate the impact of several factors affecting LAD estimations through voxel-based methods, which are still poorly understood.
Overall, we observed a strong correlation between the different estimates for LAD and reference measurements. However, these relations were found to be dependent on the type of theoretical estimator (“raw” estimator), the voxel size, the distance to scanner, the scanner type and in a lesser extent, the species. The predictions of the LAD raw estimators were corrected thanks to empirical calibrations.
4.1. Raw Estimations of the LAD
Our results showed that the predictions were sensitive to the type of estimator of the attenuation coefficient. First, we confirmed that the Contact Frequency estimator was negatively biased, at least for short distance to the scanner, as already reported by [
20] from measurements, suggested by [
12] and theoretically demonstrated by [
15]. However, we point out that such bias can be largely overwhelmed by the negative bias associated with large voxels, which might explain the results obtained in Reference [
20], who used large voxels (2 m). Second, we found that most bias corrections suggested by [
15] were only significant for very small voxels (<9 cm) and long distances (
d > 10 m). In these cases, estimators implementing bias corrections exhibited smaller overestimations and higher correlations with reference measurements than the uncorrected estimators, especially the Modified Contact Frequency. This result was promising for further applications of these corrections in the field, where occlusion—which was very limited in our laboratory experiment—leads to numerous voxels which are sampled by a small number of beams, even at short distance to the scanner, especially when fine grids are used for scene discretization (i.e., small voxels).
With the exception of these extreme cases, all raw estimators performed similarly once calibrated, including the Contact Frequency. In particular, our analysis showed a linear relationship between the reference measurements and the Contact Frequency and thus the RDI (because the mean free path is expected to be constant with vegetation density). Such a finding, already reported by [
14] in forestry plots, was somehow counterintuitive, as the response function of the
LAD to the
RDI is expected to follow the Beer-Lambert law (
), which is not linear. This departure from the Beer-Lambert law—which assumes a random distribution of vegetation elements within a voxel—could indicate that the organization of plant elements at small scale would not be random but would rather exhibit some regularity. Indeed, the fact that the absorption (i.e., RDI) was proportional to the leaf area suggests that element clumps tend to be regularly distributed and that self-shading is limited. Hence, this result suggests that TLS voxel-based methods can provide critical data to improve our understanding of plant organization at small scale, which is critical in the context of light interception and transpiration modelling [
1,
2,
3]. It should be recognized, however, that branches were not scanned under a specific orientation (i.e., from above), meaning that scanner beams do not meet the actual direct sun exposure pattern inside vegetation, which limits the interpretation in terms of light interception.
4.2. A Strong Sensitivity of LAD Prediction to Voxel Size, Which Could Mostly Arise from Vegetation Element Distribution
The first calibration step (on short-distance measurements) accounted for remaining sources of biases and uncertainties, once the distance to scanner and voxel sizes were fixed, namely leaf orientation, species, detection capacity of TLS instruments with regards to vegetation structure (beam diameter, element reflectance, sensor sensitivity, partial hit and mixed points). The resulting calibration coefficient was mostly affected by the voxel size. The other factors, such as the species, the instrument or the type of background, were of secondary importance and often not significant. Such a sensitivity to voxel size was already reported in References [
19,
20,
22].
The bias corrections for element size described in Reference [
15] slightly reduced this sensitivity when voxel were very small (5–9 cm), when compared to uncorrected estimators. Such finding was in agreement with [
22] who described similar element-size effects in small voxels (<10 cm). However, this sensitivity remained strong for both scanners, all species and all estimators, even when theoretical bias corrections were included. This hence supports the assumption that the effect of the voxel size and the subsequent underestimation of
LAD observed in largest voxels arose most likely from vegetation heterogeneity, as it was the last remaining source of (negative) bias identified at fixed distance to scanner. It should be noted that [
29] found, on the contrary, that
LAD predictions increased with voxel size, with variations from 1 to 10 in integrated
LAD predictions using the voxel-based canopy profiling method (VCP). This finding can largely be explained by the fact that the VCP method assimilates voxels containing at least one hit to “fully-vegetated” voxels, neglecting the actual vegetation densities in these voxels. As a consequence, the increase in predictions with voxel size observed in Reference [
29] may simply arise from a coarser discretization of vegetated volumes. It thus cannot be compared to the voxel-based approaches used in the present paper, as they explicitly accounted for the vegetation density inside voxels.
Noteworthy, it appeared that the short-distance calibration factors reported in the present study were remarkably stable among scanners and backgrounds. Also, this stability holds among species, even for the coniferous species, once a rigorous projection of the pine needles was applied (
Appendix A). This suggests that these calibration factors could be used in other studies, provided that the leaf morphology and the TLS instrument did not exhibit large differences with those of the present study.
Finally, this potentially critical influence of vegetation distribution on
LAD prediction highlights the problem of methods based on gap fraction, which generally assume that the vegetation distribution is homogeneous in a horizontal layer, to inverse the transmission equation [
10,
11,
30]. Hence, such an approach is theoretically equivalent to the application of a voxel-based method to thin “pancake” voxels with a horizontal extent which encompasses the forest plot, thus potentially leading to
LAD underestimations.
4.3. The Sensitivity to Distance to Scanner Led to the Notion of Effective Footprint and Revealed the Existence of Spatial Bias in LiDAR Plot-Scale Estimations
In addition to these variations at 2.5 m, a strong effect of distance was revealed for the FARO instrument and in a smaller extent for the RIEGL instrument. The farther the scanner was from the vegetation, the higher the estimated LAD was. We would like to highlight the importance of this finding, which reveals the presence of equally large spatial biases in measurements at the scale of a vegetation scene (e.g., forest plot) when raw estimates are not corrected for the distance effect, which is the case of most studies. Indeed, plot-scale or tree-scale measurements combine various estimations done at several distances to scanner, especially when the canopy is high. As a result, an estimated LAI of 2 in a 5-m or in a 20-m high canopy might correspond to considerably different actual LAI. As for the voxel-size effect, such a distance effect also leads to bias in gap fraction estimates, as their computation rely on the same theoretical basis. Hence, they should also be corrected for distance effect.
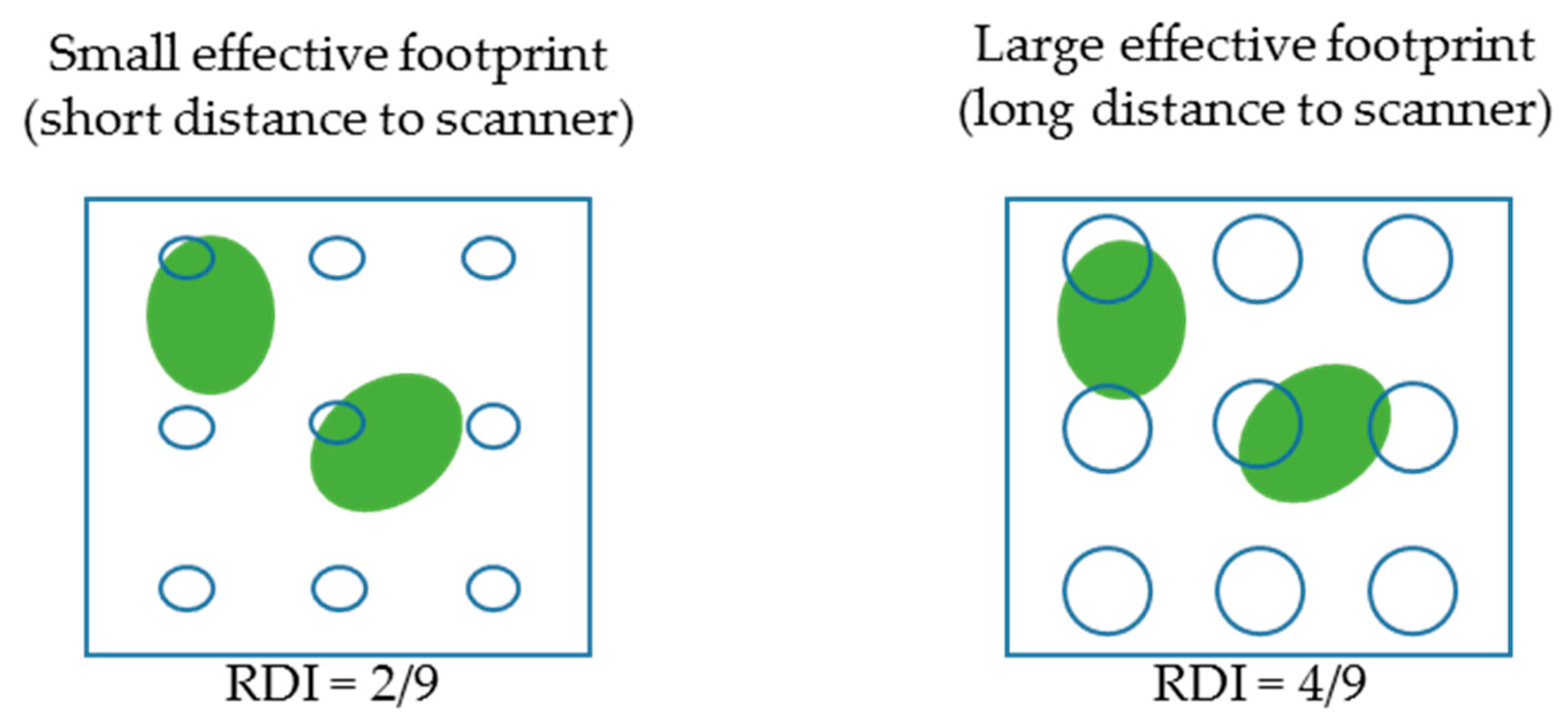
As explained in the introduction, this distance effect could be explained by an increase in the hit probability resulting from the increase of laser footprint with distance to scanner, induced by beam divergence ([
12]). Indeed, when beam diameters increase with distance to scanner, beams lose their ability to pass through vegetation elements without triggering a hit, leading to higher
RDI and hence,
LAD estimates (
Figure 9). In other word, vegetation elements are “seen” by the scanner bigger than they actually are, as returns can be registered even when the beam centre lines do not intercept any elements. Another explanation to the sensitivity to distance might have been the decay of beam number in voxels, which causes a positive theoretical bias in uncorrected estimates [
15]. However, we observed similar increases with distance when this theoretical bias was corrected, suggesting that the beam divergence was the main cause of observed sensitivity to distance. The distance effect was found to slightly vary among species (
Figure 7), the effect being stronger for species with smaller vegetation elements (pine needles) than with larger leaves. This was consistent with the above explanation, as the relative increase in probability of interception was found to be bigger for fine than for large elements. The specifications provided by the instrument manufacturers entail to estimate a theoretical footprint at the exit (beam diameters equal to 6.5 mm and 2.25 mm for the RIEGL and the FARO, respectively) and at further distances (beam divergence equal to 0.35 mrad and 0.19 mrad for the RIEGL and the FARO, respectively). From these theoretical specifications, the footprint of the RIEGL at 2.5 m (42.7 mm
2) should be much larger than the one of the FARO (5.8 mm
2), so that the raw estimates of the FARO should have been much lower than those of the RIEGL. This contradicts the results observed at 2.5 m (calibration coefficient not being sensitive to the instrument) and suggests that the raw estimates do not only reflect the size of the theoretical footprint but also other characteristics of the scanner and how beams interact with vegetation elements (element reflectance, sensor sensitivity, partial hit management).
In practice, a beam diameter of a given size does not mean that a scanner can detect vegetation within this whole footprint. Indeed, a hit is registered only when the intensity of the return is higher than a given threshold [
11], so that not all partial hits are detected. To facilitate data interpretation, we suggest the notion of effective footprint (or equivalently effective beam size), which would be the mean footprint area within which vegetation elements are actually detected in a given voxel. This effective footprint would depend of the beam diameter but also of the sensor detection threshold, the return intensity and vegetation element properties and is expected to be smaller than the actual footprint of the scanner. Our short-distance analyses suggest that both the FARO and the RIEGL exhibited similar effective footprints at
d = 2.5 m, since the calibration coefficient obtained with both instruments were similar, despite of the differences in theoretical footprints (possibly because of a lower detection threshold of the RIEGL, which might have offset its larger footprint). The distance effect pattern suggested that the effective footprint of the RIEGL was more or less constant with distance up to 10 m, before slightly increasing at further distances. Conversely, the effective footprint of the FARO strongly increased before 10 m and was constant at further distances. A physical modelling of such an effective footprint is not straightforward, even if some physical bases were provided in Reference [
11]. However, the physical approach is limited with scanner such as the FARO, as several factors affect the registered value of phase-shift scanner intensity, such as temperature [
31] and shooting direction in the different phases. The methodology presented here provided a promising alternative to calibrate the sensitivity to distance.
The magnitude of the distance effect decreases with voxel size (
Figure 7), which was somehow counterintuitive. This observation could arise from the heterogeneous structure of the vegetation, as suggested by the following mechanism. Smaller voxels lead to more frequent “empty” voxels, as their size approaches the size of gaps between shoots in a branch. Hence, the raw
LAD distribution is wider on a finer grid. As shown in
Appendix E, the distance effect was much stronger in voxels with low density, whereas it was smaller and fairly constant at medium and high densities. As a result, the changes in raw
LAD distribution due to smaller voxels (in particular the increase in frequency of low density voxels) induce a higher frequency of voxels with stronger distance effect and hence a slightly stronger overall distance effect (at branch scale) with small voxels.
4.4. Calibrated Estimator Performance
Overall, calibrated raw LAD estimates exhibited lower error with the RIEGL than with the FARO scanner. Hence, the RIEGL obtained the lowest errors with Q. ilex for all voxel sizes (about 15%). The better performance of the RIEGL instrument (despite no correction for distance was applied) might be explained by a better management of mixed pixels and a lower level of noise with time-of-flight than with phase-shift scanners. Indeed, the RIEGL scanner registers the 3D coordinates of the location where the return intensity was the highest, whereas the FARO scanner combines the information arising from multiple phase shifts (3 wavelengths for the FARO), leading to the registration of a hit position located within the shortest and longest distances at which intensity was returned, which does not necessarily correspond to the location where the actual vegetation element is. Hence, the FARO might be more subject to hit misplacement. However, the performance of the RIEGL scanner was generally lower with P. halepensis compared to the FARO scanner, especially for the smallest voxel sizes (<15 cm).
Overall, with the exception of
Q. ilex, a decrease in voxel size led to higher errors in
LAD predictions with both scanners, even with theoretically TBC estimators (
Table 3). It has been demonstrated [
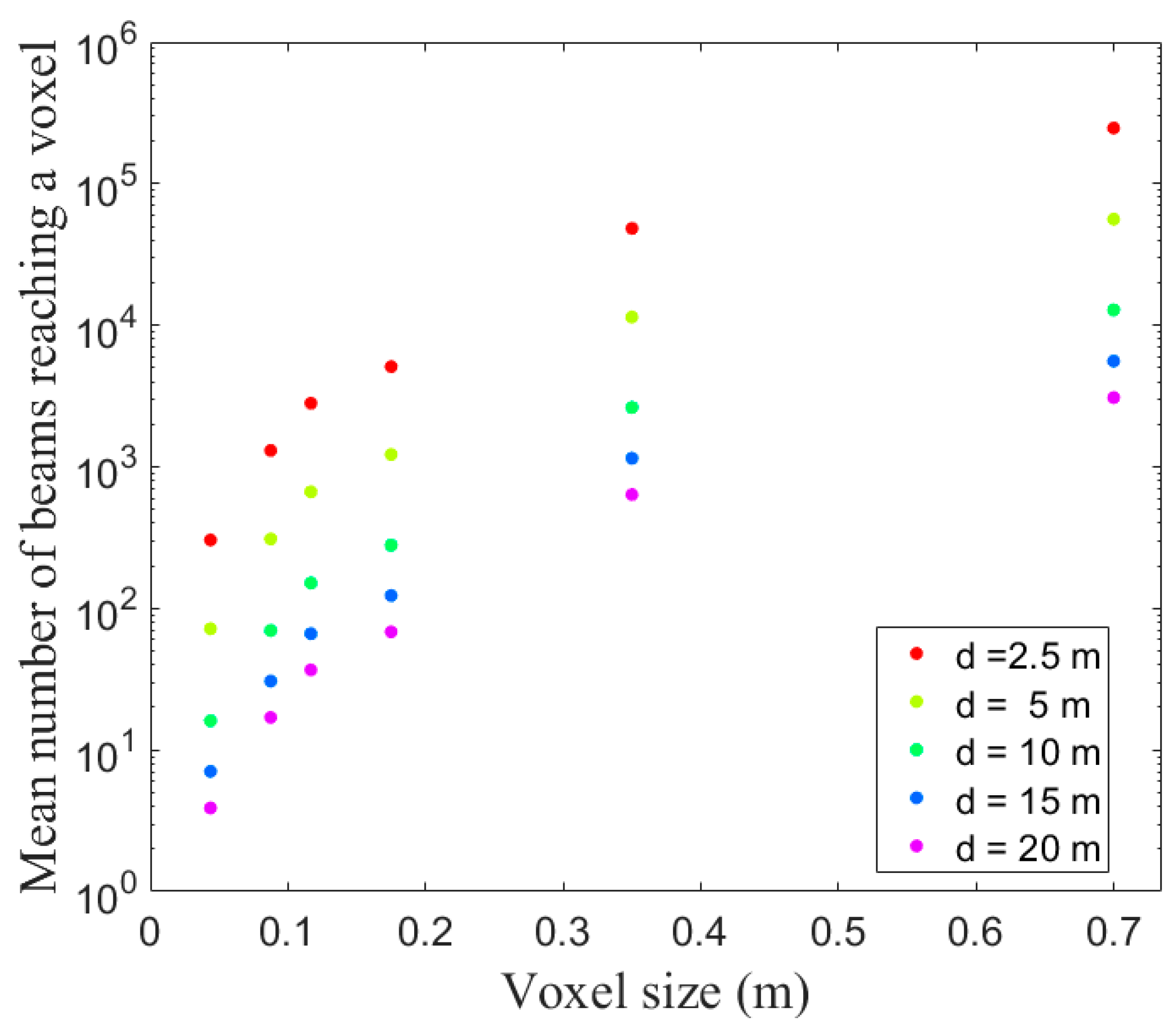
15] that the variability in raw estimates sharply increased with decreasing number of beams and number of vegetation elements within a voxel, leading to large measurements errors. The increase in errors observed below 10 cm suggests that the overall measurement accuracy at branch scale was affected by the measurement errors resulting from random effects occurring at small scale. Indeed, 10 cm appeared as a good compromise between limited error arising from uncertainties of measurements in small voxels and an adequate representation of spatial heterogeneity. It is important to notice, however, that similar measurement error can be achieved in larger voxels, provided that a relevant calibration was applied.
The lower performance of the more sophisticated (TBC) estimators when applied to the FARO scanner at small voxel size suggests that these estimators were more sensitive to measurement error and noise than simple estimators (i.e., CF). This could be interpreted as an example of the bias-variance trade-off. Indeed, sophisticated estimators were theoretically unbiased, meaning that their expectation over a large number of trials was closer to the actual value. [
15] showed that their variance was the smallest that could be expected from any unbiased estimators (since they reached the Cramer-Rao bound, see [
15]) but also that this variance was high (leading to 95% confident interval as large as 100% of the expectation when beam number was low and vegetation elements were large). Moreover, the TBC estimators do not account for vegetation heterogeneity, a major source of bias as suggested in the present paper. As a result, it is not surprising that once calibrated at the scale of interest (here the branch), more basic estimators with smaller variance (such as the basic contact frequency approach) were more robust to noise and led to smaller errors than the TBC estimator. Another potential cause of limited performance of the bias-corrected estimator, might be the fact that the spatial arrangement of vegetation would not be random at small scale, as suggested in
Section 4.1.
These findings, based on detailed laboratory data, highlight the critical need for detailed field data and actual TLS data to validate or invalidate finding arising from theory and simulations.
4.5. Recommendations and Further Work
Our study confirmed a strong sensitivity of the LAD estimation to discretization arising from vegetation heterogeneity and revealed a strong distance effect which, if not corrected, can lead to large spatial biases in LiDAR plot-scale estimates. The observed variations among tested scanners showed that emission pattern and properties of beams from LiDAR instruments play an important role in results and in spatial robustness of estimators. We thus recommend that these mechanisms are accounted for in further studies addressing the estimation of LAD with TLS. These recommendations apply not only to voxel-based methods but also to gap fraction approaches.
The method presented in the present study (calibration based on experimental tests on branches) could be applied to other species, estimators and instruments as a first step to improve LAD estimations by the use of calibration functions. The evaluation/calibration procedure developed in this study can be replicated because it is easily reproducible and reasonably time-consuming (1 week per species). In addition, the same calibration could be applied to species exhibiting close vegetation structure. This method can be seen as a fast approach to characterize bias occurring in LAD estimations.
It is however important to recognize that such laboratory studies do not replace experimental approaches at plant [
12] or stand scales [
14], as additional issues, such as determining the leaf fraction or accounting for occlusion should be tackled to assess reliable
LAD [
19]. Occlusion was avoided on purpose in this study to ensure detection of vegetation elements and achieve corrections of biases related to vegetation heterogeneity and scanner properties only. The occurrence of occlusion on field might hence limit reliability of our estimations, even though this work deals with some occlusion issues. Indeed, tested TBC estimators theoretically account for decrease in number of beams used for voxel sampling, which is a major drawback of occlusion. In addition, small voxel sizes used in this study (e.g., 0.09 m) enable an accurate pinpointing of potentially occluded areas. Nevertheless, other biases (i.e., heterogeneous sampling of voxel or totally occluded areas inside tree crowns) are not addressed as it was not the purpose of this work. Large scale specific experiments at tree or stand scale are also required to evaluate the calibration derived from laboratory experiments.
Finally, the RIEGL scanner was used in this experiment similarly as a single return instrument, that is, neglecting further impacts (second, third and fourth returns) which only represent 4% of total registered returns. This low percentage of multiple returns was mainly due to the background characterized by an obstacle-free distance of at least 100 m in the scan direction when the RIEGL scanner was used (see
Section 2.1). However, in field measurements, a significant increase in the number of multiple returns is expected due to the surrounding vegetation. Considering first returns only results in an underestimation of voxel transmittance, that is, an overestimation of RDI and LAD. Indeed, in that case, for each return, the vegetation is implicitly assumed to fully intercept the laser beam or, in other words, to cover the whole beam area. Most of the time, part of the beam energy is not intercepted and the leaf area responsible for an echo detection is smaller than the whole footprint area. It has been demonstrated that using multiple returns information enhanced LAD estimates and that the smaller the leaves, the more significant the improvement is [
13]. To take into account this information several strategies have been proposed to weight echoes when computing the RDI. Weighting can be based either on echo intensities [
19] or on the number [
13] or on the number and rank [
32], of the echoes belonging to the same laser beam. However, taking into account partial interceptions remains challenging due to the numerous factors that influence echo intensity and also because information about the amount of energy remaining in the laser beam after the last echo, if any, is always missing.
When dealing with partial interceptions by using multiple returns information, raw LAD estimates and, consequently, appropriate calibration coefficients can thus depend on voxel environment. For these reasons, it should be more effective and accurate to assess and use the calibration coefficients obtained with the proposed approach, considering first returns only when working with field measurements.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}










