An Empirical Algorithm for Estimating Agricultural and Riparian Evapotranspiration Using MODIS Enhanced Vegetation Index and Ground Measurements of ET. I. Description of Method


Abstract
:1. Introduction
1.1. The Vegetation Index Approach for Scaling ET by Remote Sensing
1.2. Potential Problems in Applying VI Methods to Arid Zone Plant Communities
1.3. Goal and Objectives
2. Methods
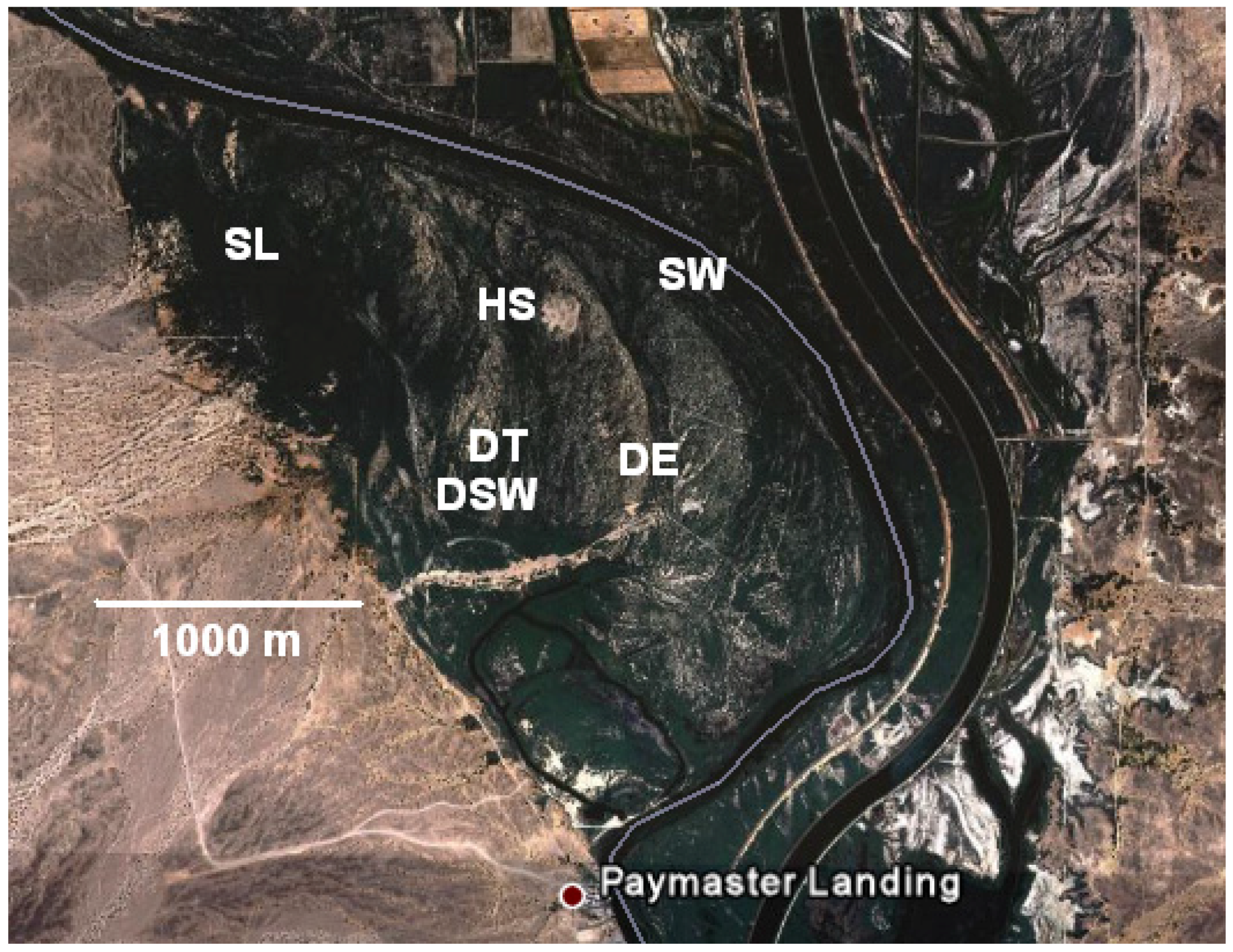
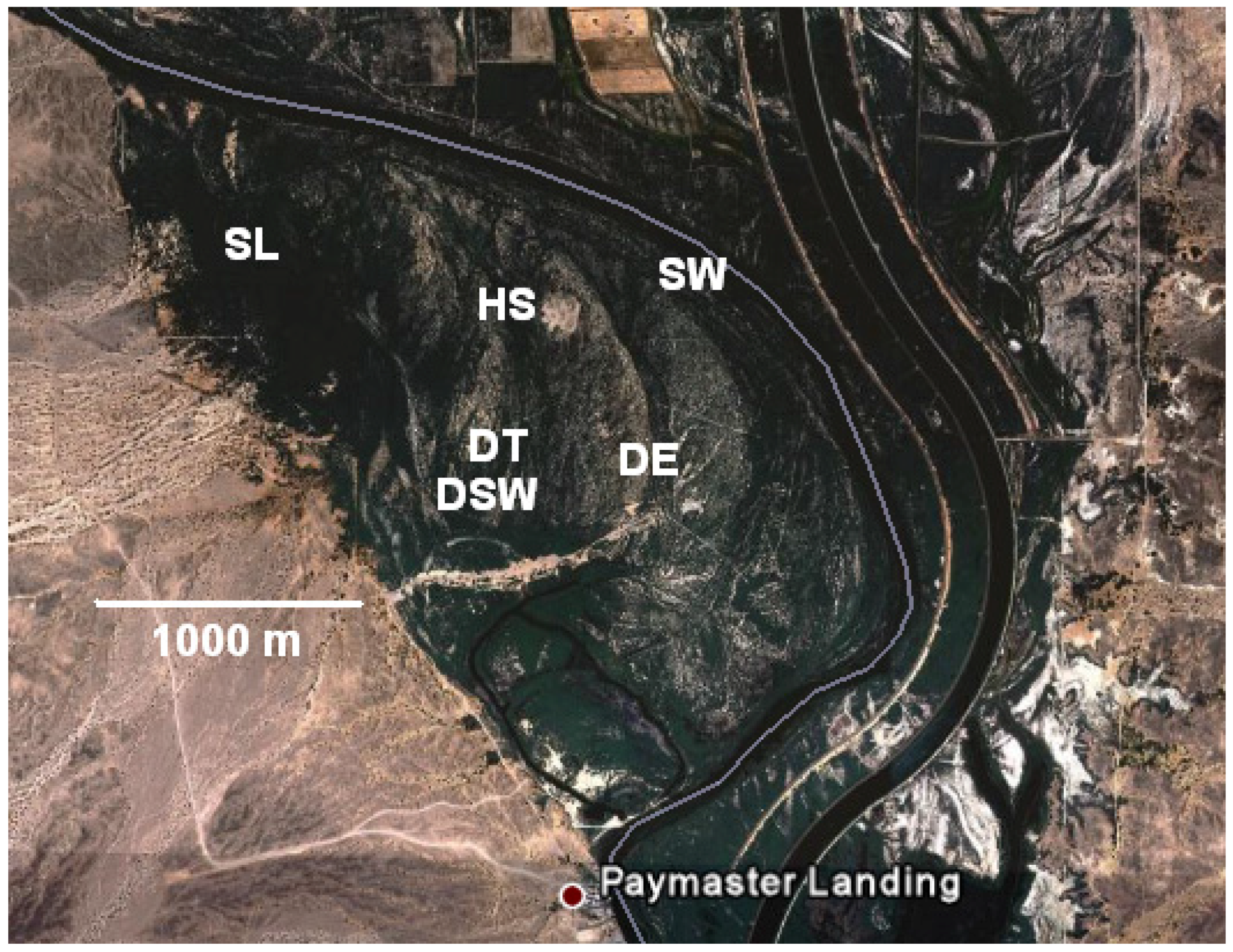
2.1. Site Description

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Lat/Lon (o North, o West) | Distance From River (m) | Soil Texture (S-Si-Cl) | Depth to Aquifer (m) | Aquifer Salinity (dS m−1) | Aquifer Temperature (oC) |
|---|---|---|---|---|---|---|
| Swamp | 33.2754, 114.6873 | 200 | 25-55-20 | 2.35 (0.03) | 3.90 (0.18) | 23.1 (0.3) |
| Slitherin | 33.2746, 114.7098 | 750 | 40-45-15 | 3.51 (0.06) | 4.72 (0.48) | 22.5 (0.2) |
| Diablo Tower | 33.2659, 114.6992 | 1500 | 50-35-15 | 2.61 (0.06) | 15.49 (0.18) | 22.8 (0.3) |
| Diablo Southwest | 33.2663, 114.7002 | 1550 | 90-7-3 | 2.79 (0.06) | 24.0 (0.55) | 22.2 (0.2) |
| Diablo East | 33.2687, 114.6895 | 870 | 95-2-3 | 2.40 (0.04) | 7.1 (0.19) | 24.9 (0.2) |
| Hot Springs | 33.2783, 114.6924 | 557 | 85-10-5 | 2.36 (0.03) | 5.39 (0.07) | 51.0 (0.5) |
| Hayday Farms Alfalfa | 33.4683, 114.6938 | 8,439 | - | 1.81 (0.05) | 1.85 (0.05) | 23.0 (0.35) |
2.2. Sap Flow Measurements
2.3. Scaling ETactual to Whole Plants and Stands of Plants Based on Leaf Area Index
2.4. Measurement of Alfalfa ETactual
2.5. MODIS Data
2.6. Meteorological Data and Other Calculations
2.7. Other Sources of EG and ETactual Data
3. Results
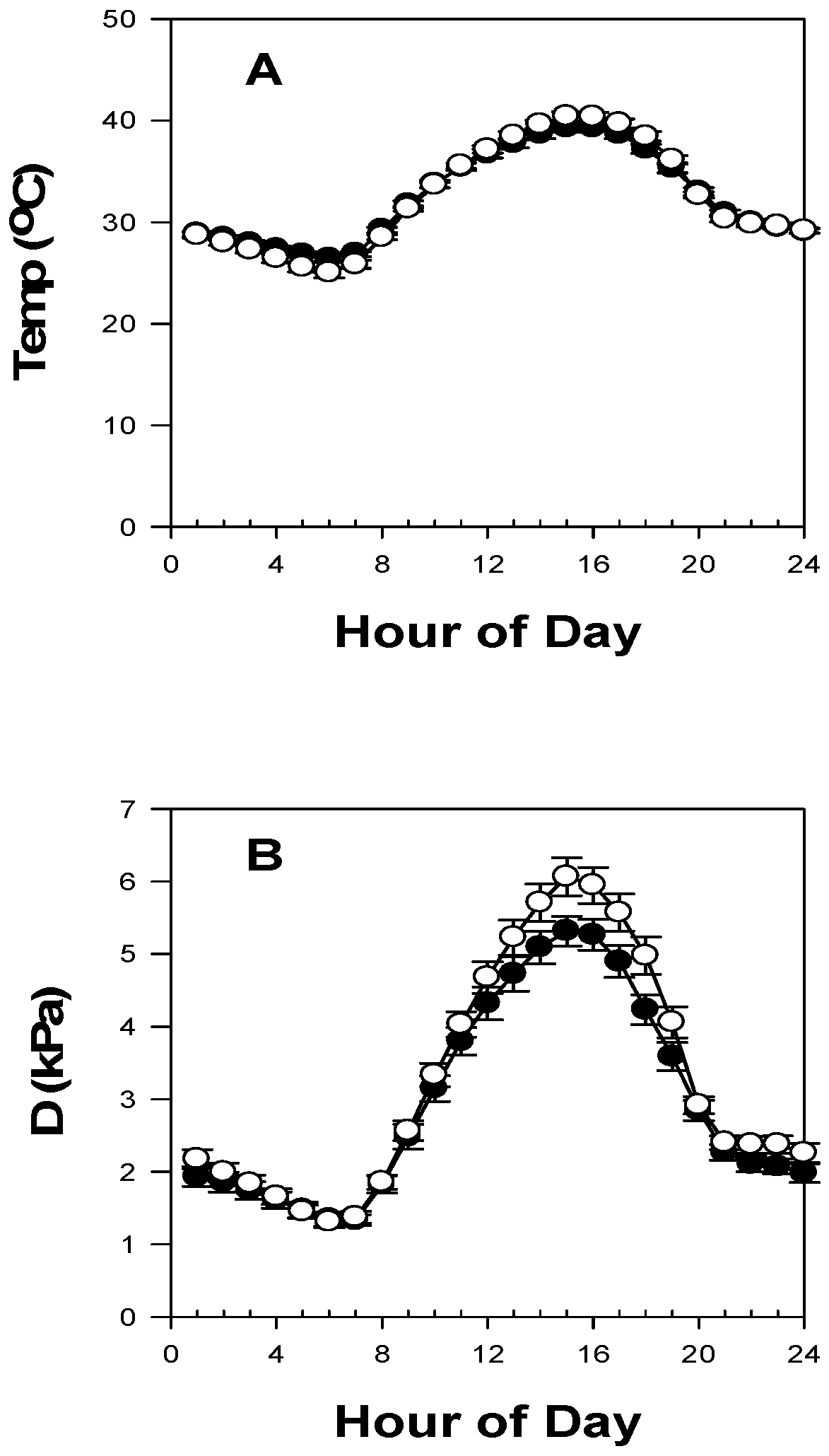
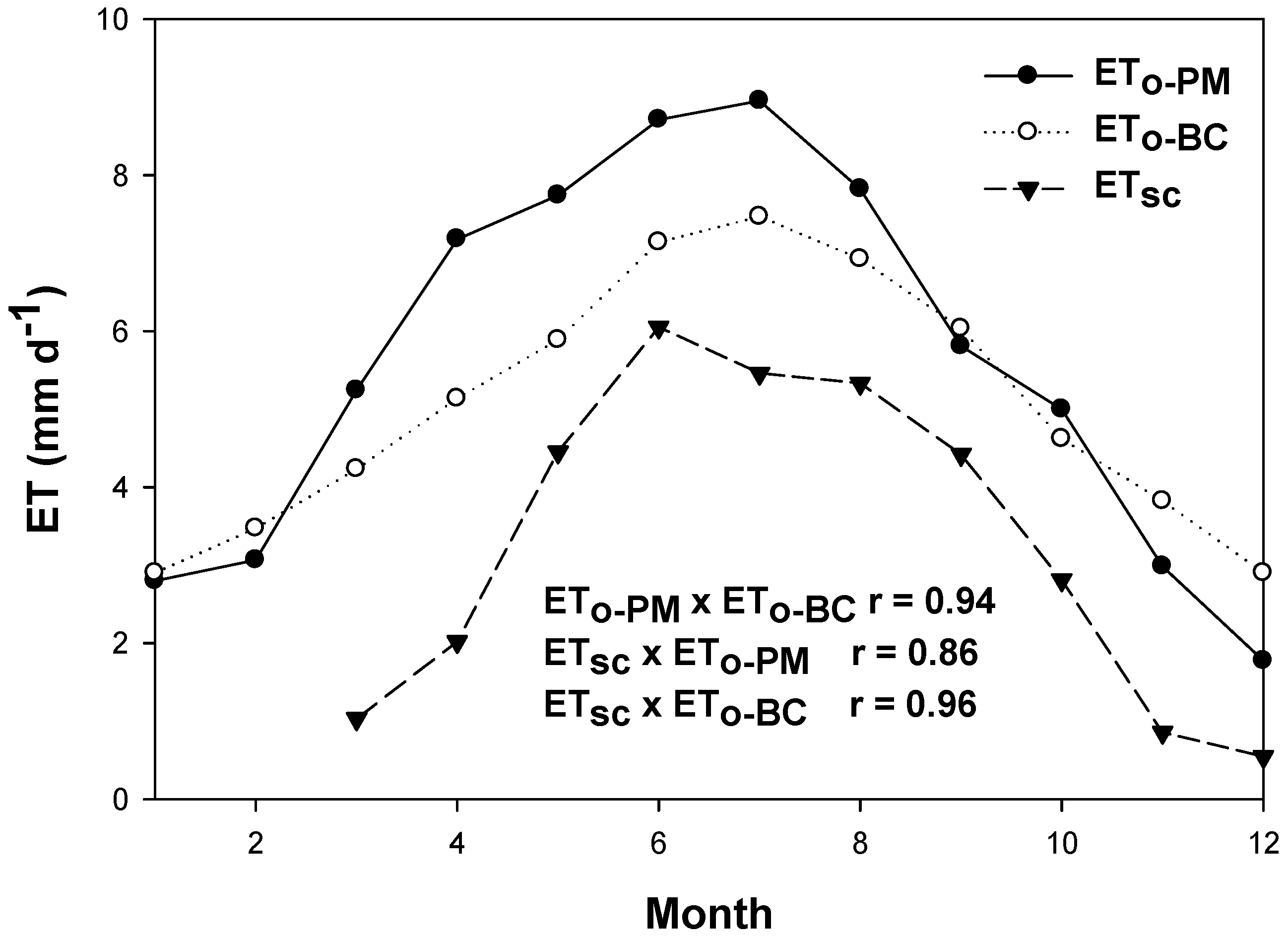
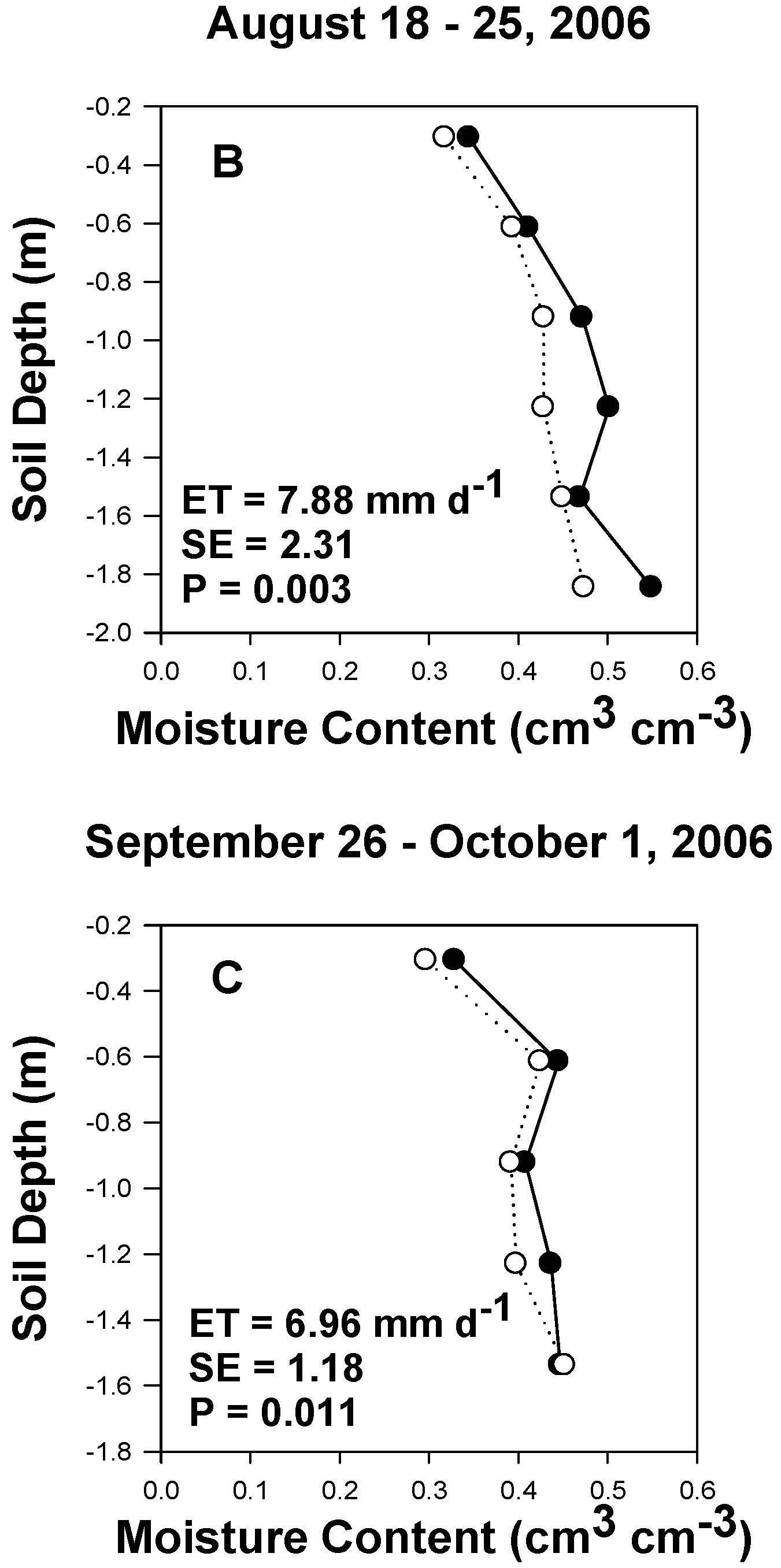
3.1. Meteorological, Soil and Aquifer Conditions at Saltcedar and Alfalfa Sites

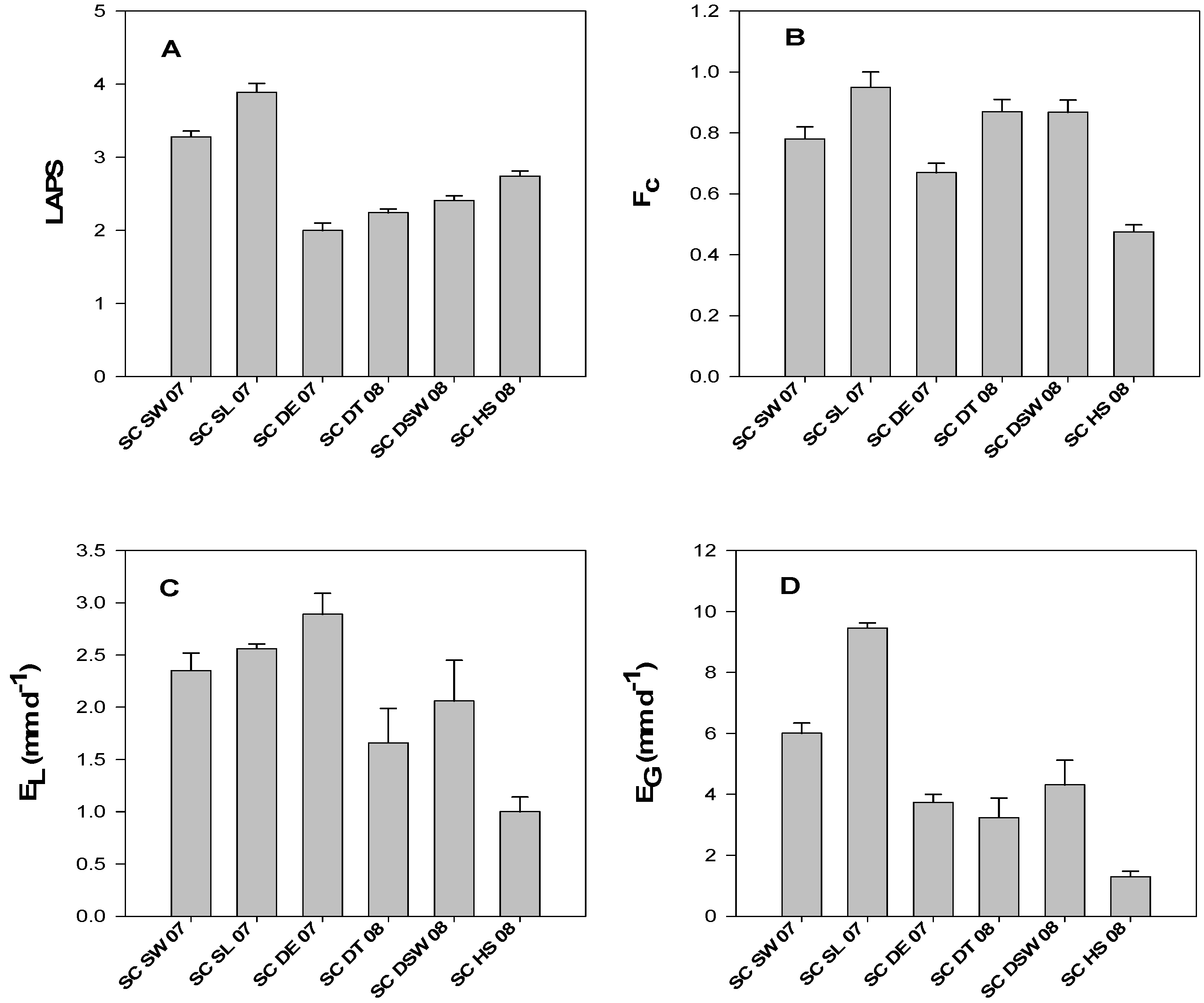
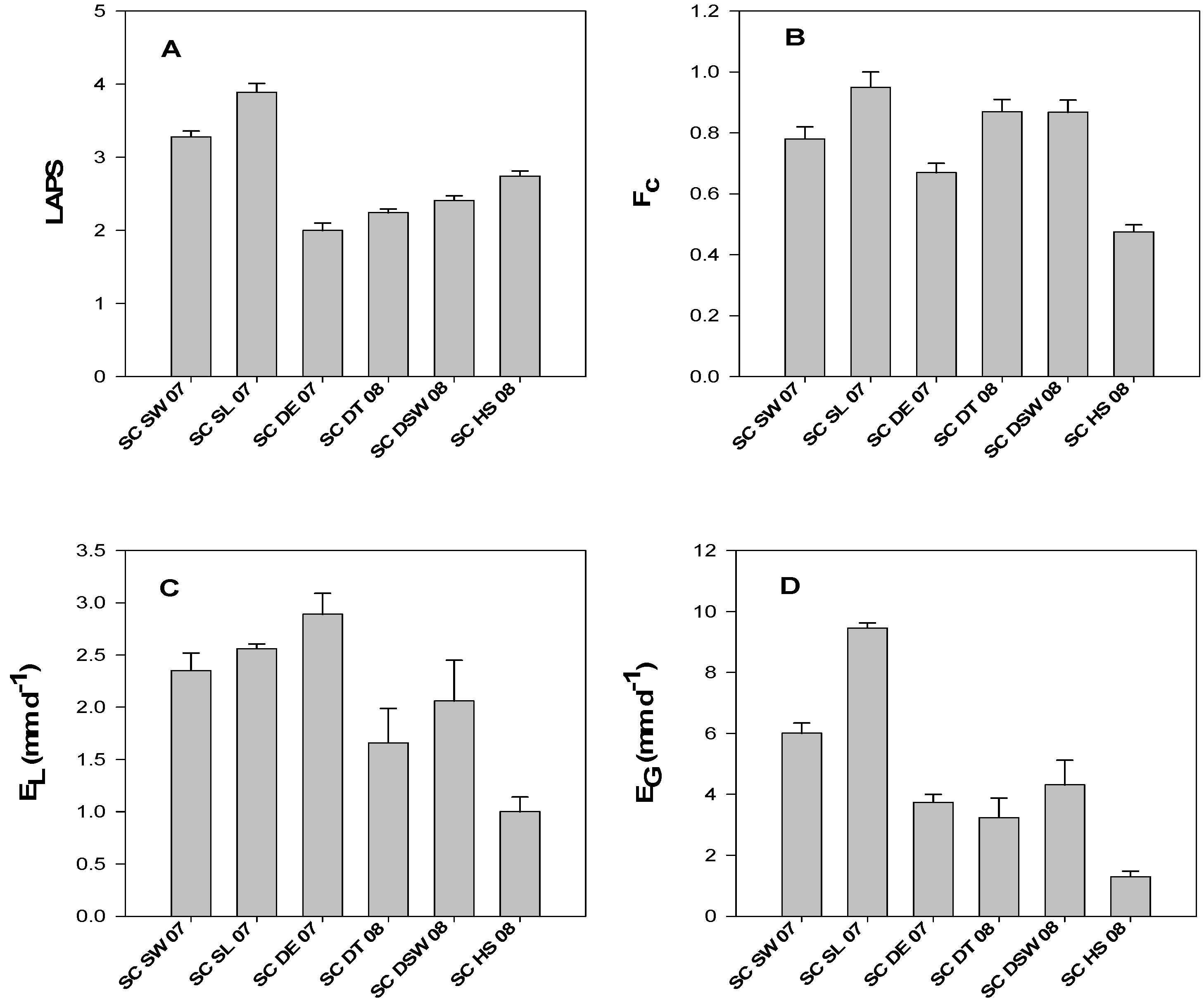
3.2. LAPS, fc, EL, and EG of Saltcedar Stands
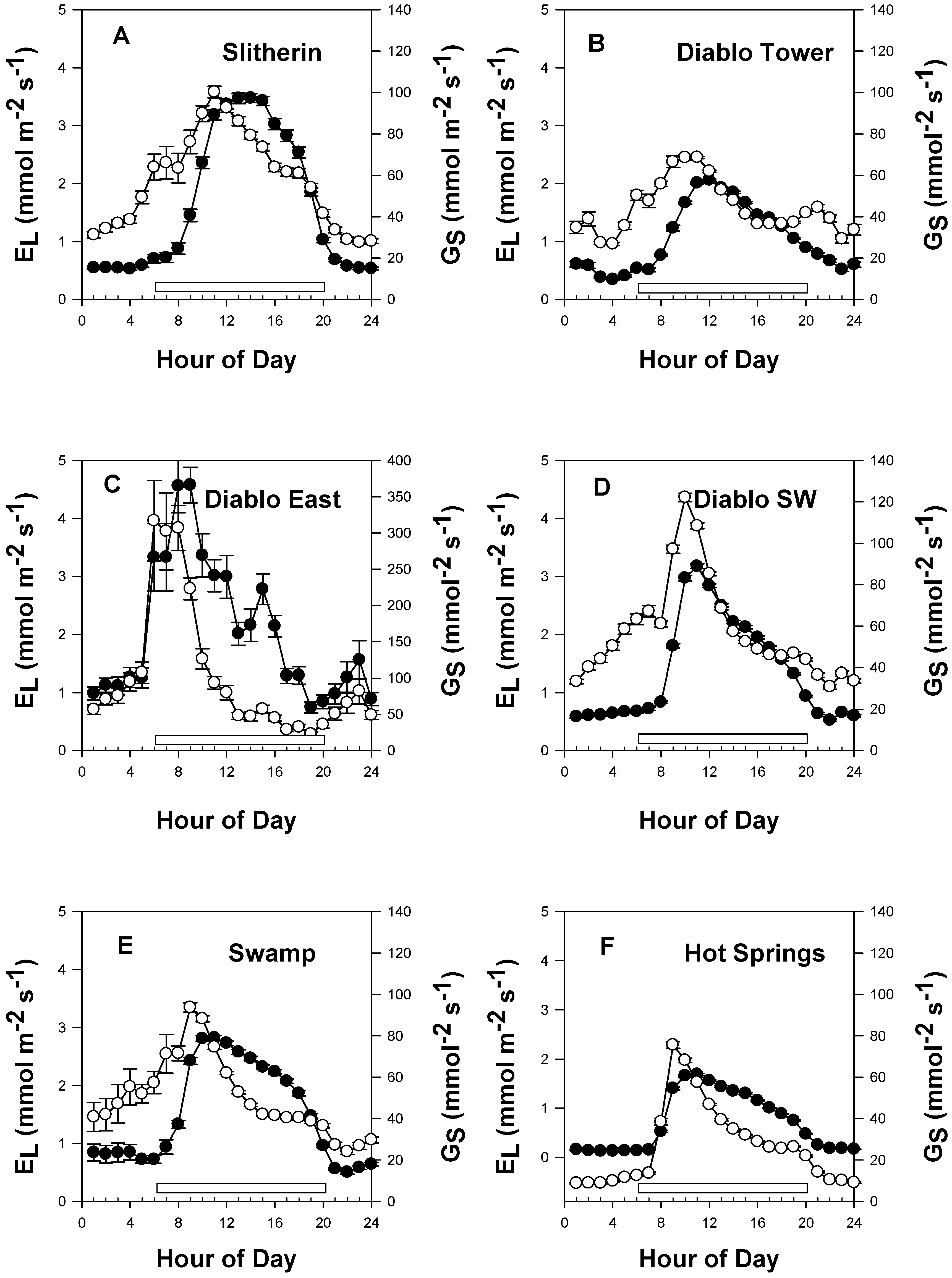
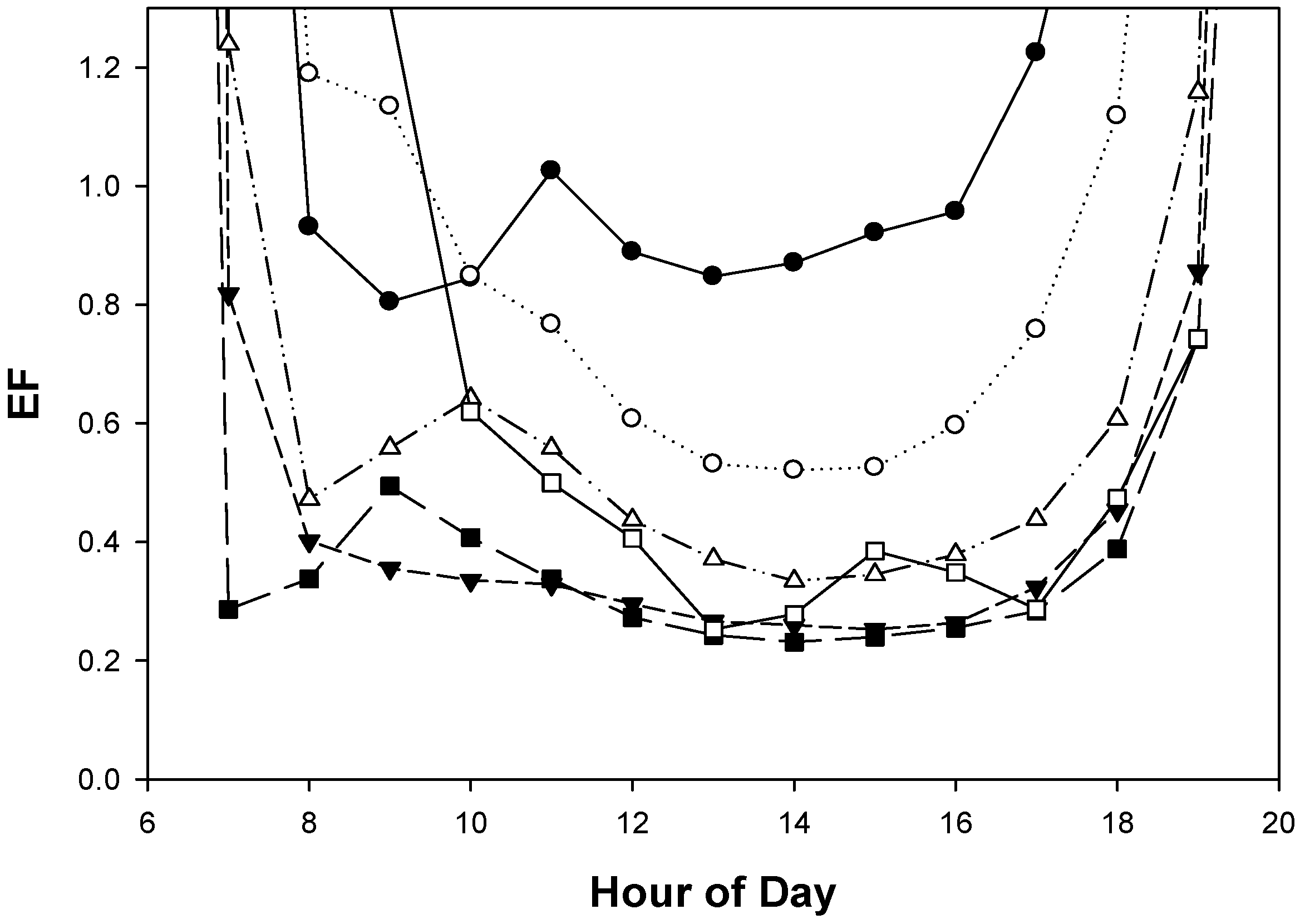
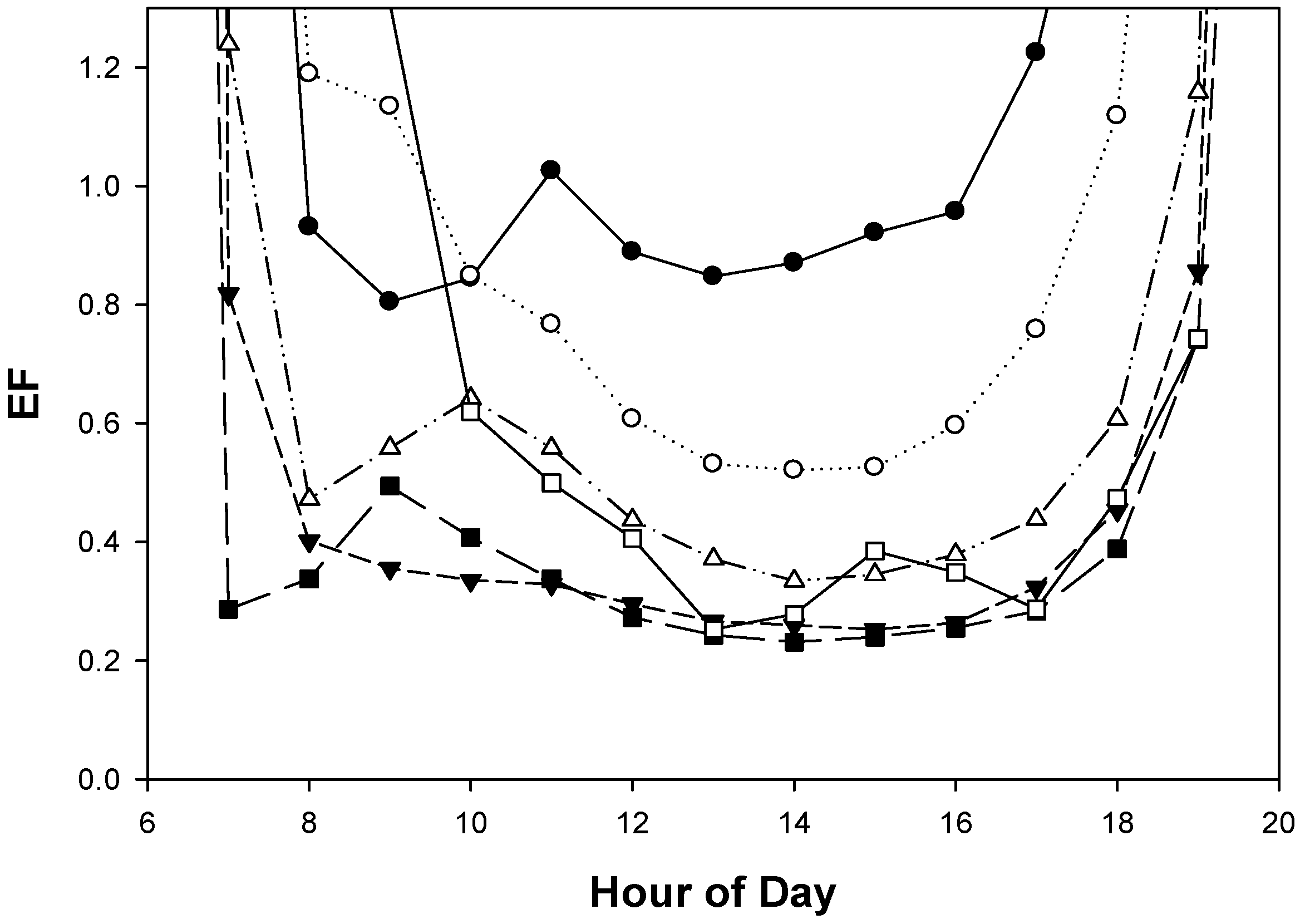
3.3. Diurnal Variation in EL, GS and EF

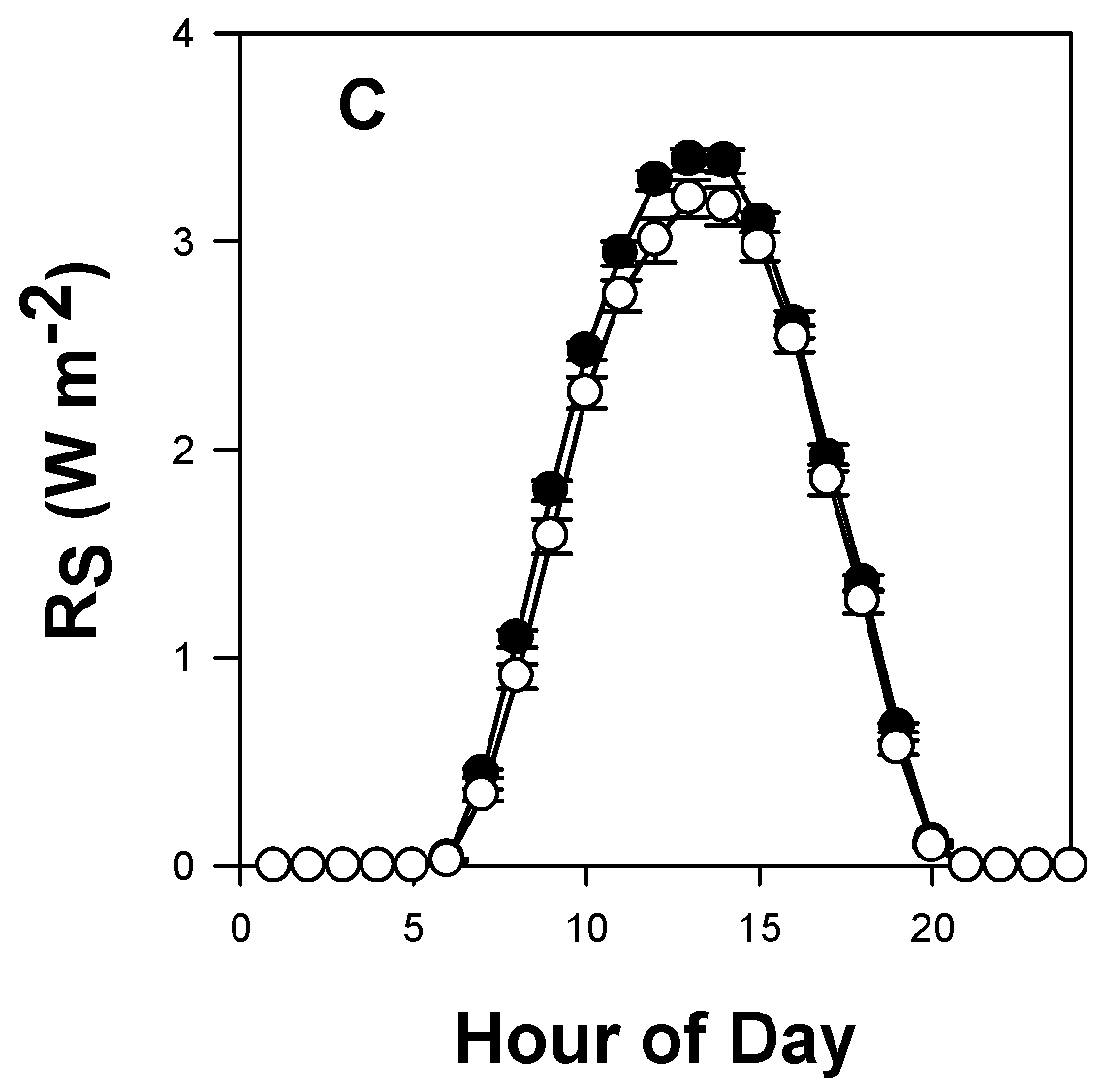
3.4. Alfalfa ETactual

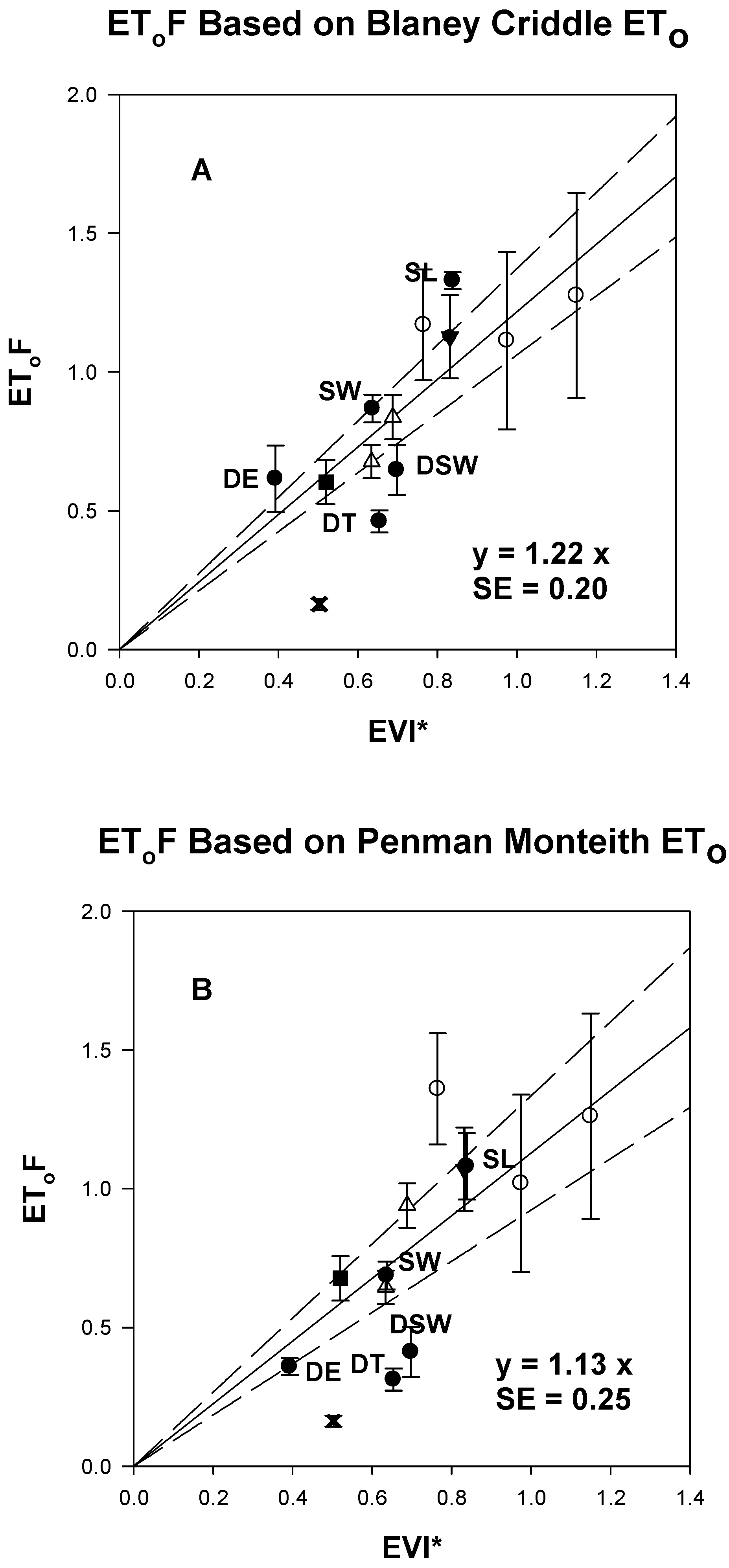
3.5. Linear Equations for EGF and EToF Based on EVI*
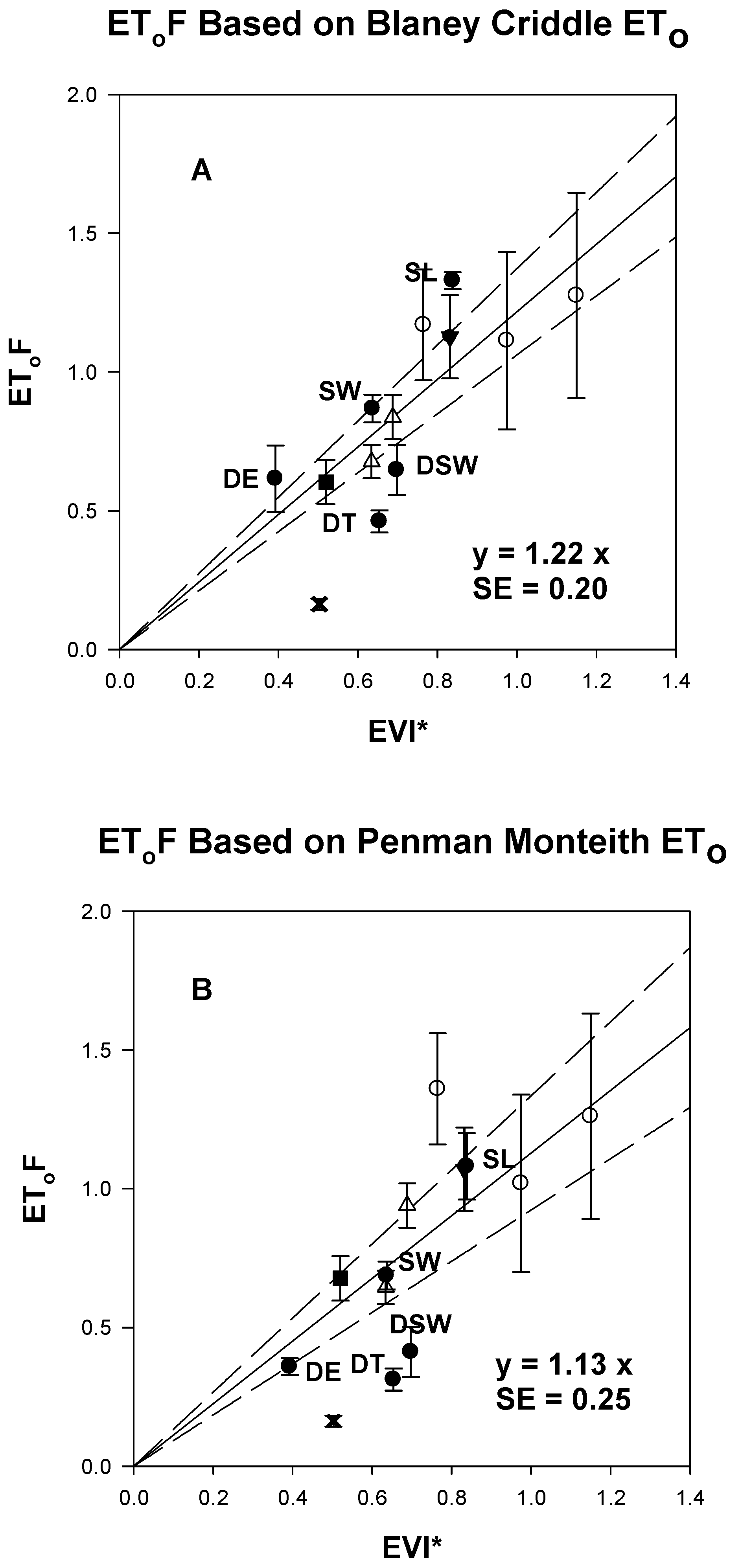
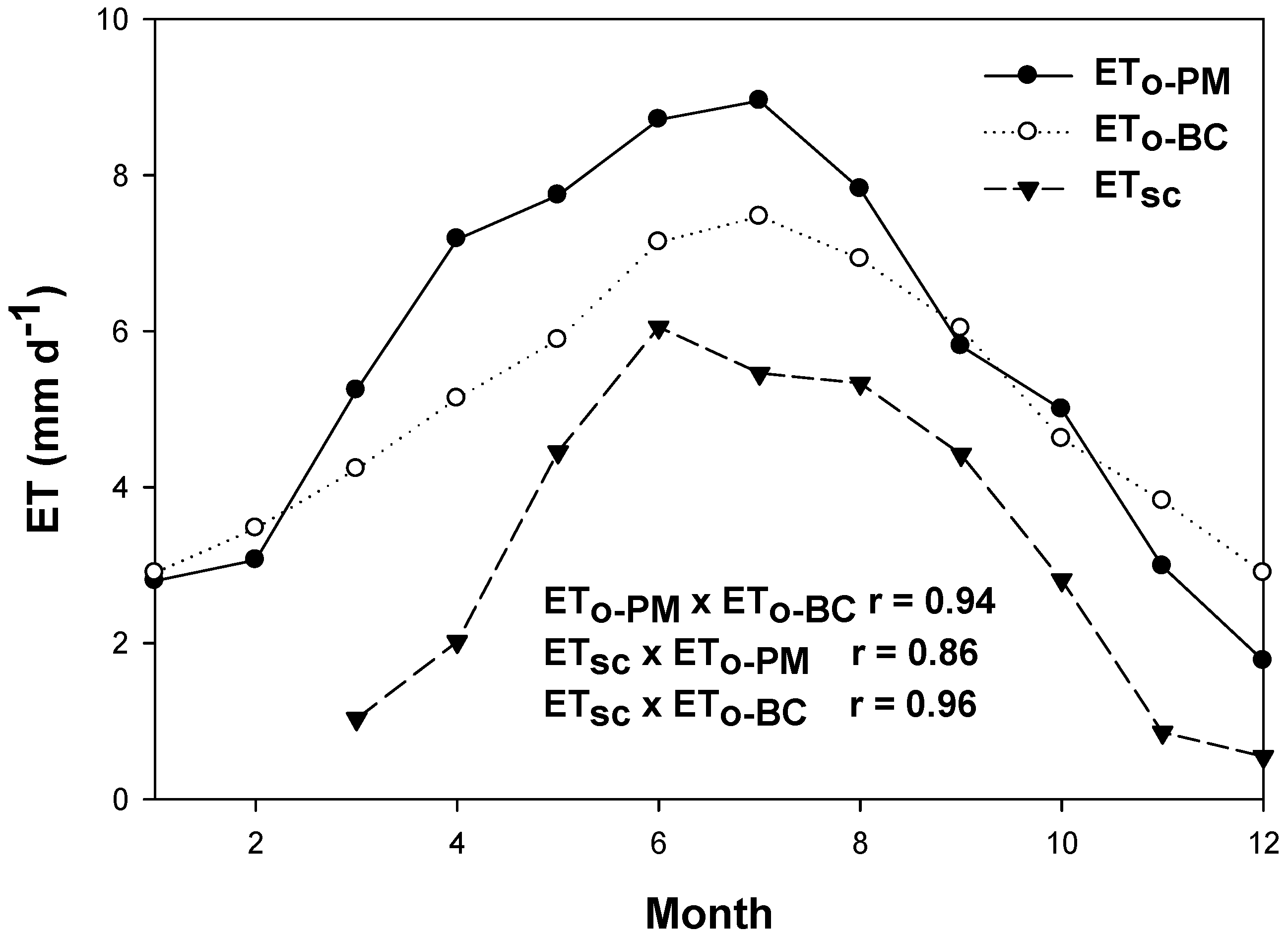
4. Discussion
4.1. Diurnal Patterns of Saltcedar EL, GS and EF Indicate Stress
4.2. Remote Sensing Algorithm for Scaling ETactual
4.3. Comparison with Other Remote Sensing Methods for ET
4.4. Sources of Error and Uncertainty
5. Conclusions
Acknowledgements
References and Notes
- Groeneveld, D.P.; Baugh, W.M.; Sanderson, J.S.; Cooper, D.J. Annual groundwater evapotranspiration mapped from single satellite scenes. J. Hydrol. 2007, 344, 146–156. [Google Scholar] [CrossRef]
- Glenn, E.; Morino, K.; Didan, K.; Jordan, F.; Nagler, P.; Waugh, J.; Carroll, K.; Sheader, L. Scaling sap flux measurements of grazed and ungrazed shrub communities with fine and coarse-resolution remote sensing. Ecohydrol. 2008, 1, 316–329. [Google Scholar] [CrossRef]
- Nagler, P.L.; Glenn, E.P.; Kim, H.; Emmerich, W.; Scott, R.L.; Huxmam, T.E.; Huete, A.R. Relationship between evapotranspiration and precipitation pulses in a semiarid rangeland estimated by moisture flux towers and MODIS vegetation indices. J. Arid. Environ. 2007, 70, 443–462. [Google Scholar] [CrossRef]
- Hunsaker, D.J.; Fitzgerald, G.J.; French, A.N.; Clarke, T.R.; Ottman, M.J.; Pinter, P.J. Wheat irrigation management using multispectral crop coefficients. I. Crop evapotranspiration prediction. Trans. ASABE 2007, 50, 2017–2033. [Google Scholar] [CrossRef]
- Kim, J.Y.; Hogue, T.S. Evaluation of a MODIS-based potential evapotranspiration product at the point scale. J. Hydrometeor. 2008, 9, 444–460. [Google Scholar] [CrossRef]
- Gonzalez-Dugo, M.P.; Neale, C.M.U.; Mateos, L.; Kustas, W.P.; Anderson, M.C.; Li, F. A comparison of operational remote-sensing based models for estimating crop evapotranspiration. Agr. Forest Meteor. 2009, 49, 2082–2097. [Google Scholar] [CrossRef]
- Nagler, P.; Scott, R.; Westenburg, C.; Cleverly, J.; Glenn, E.; Huete, A. Evapotranspiration on western US rivers estimated using the Enhanced Vegetation Index from MODIS and data from eddy covariance and Bowen ratio flux towers. Remote Sens. Environ. 2005, 97, 337–351. [Google Scholar] [CrossRef]
- Nagler, P.L.; Morino, K.; Didan, K.; Osterberg, J.; Hultine, K.; Glenn, E. Wide-area estimates of saltcedar (Tamarix spp.) evapotranspiration on the lower Colorado River measured by heat balance and remote sensing methods. Ecohydrol. 2009, 2, 18–33. [Google Scholar] [CrossRef]
- Juarez, R.I.N.; Goulden, M.L.; Myneni, R.B.; Fu, R.; Bernardes, S.; Gao, H. An empirical approach to retrieving monthly evapotranspiration over Amazonia. Int. J. Remote Sens. 2008, 29, 7045–7063. [Google Scholar] [CrossRef]
- Leuning, R.; Zhang, Y.Q.; Rajaud, A.; Cleugh, H.; Tu, K. A simple surface conductance model to estimate regional evapotranspiration using MODIS leaf area index and the Penman-Monteith equation. Water Resour. Res. 2008, 44, Article No. W10419. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Chiew, F.H.S.; Zhang, L.; Leuning, R.; Cleugh, H.A. Estimating catchment evaporation and runoff using MODIS leaf area index and the Penman-Monteith equation. Water Resou. Res. 2008, 44, Article No. W10420. [Google Scholar] [CrossRef]
- Mu, Q.; Heinsch, F.A.; Zhao, M.; Running, S.W. Development of a global evapotranspiration algorithm based on MODIS and global meteorology data. Remote Sens. Environ. 2007, 111, 519–536. [Google Scholar] [CrossRef]
- Wang, K.C.; Liang, S.L. An improved method for estimating global evapotranspiration based on satellite determination of surface net radiation, vegetation index, temperature, and soil moisture. J. Hydrometeor. 2008, 9, 712–727. [Google Scholar] [CrossRef]
- Kalma, J.D.; McVicar, T.R.; McCabe, M.F. Estimating land surface evaporation: a review of methods using remotely sensed surface temperature data. Surv. Geophys. 2008, 29, 421–469. [Google Scholar] [CrossRef]
- Kustas, W.; Norman, J. Use of remote sensing for evapotranspiration monitoring over land surfaces. Hydrologic. Sci. J. 1996, 41, 495–516. [Google Scholar] [CrossRef]
- Glenn, E.; Huete, A.; Nagler, P.; Hirschboek, K.; Brown, P. Integrating remote sensing and ground methods to estimate evapotranspiration. Crit.Rev. Plant Sci. 2007, 26, 139–168. [Google Scholar] [CrossRef]
- Glenn, E.; Huete, A.; Nagler, P.L.; Nelson, S.G. Relationship between remotely-sensed vegetation indices, canopy attributes and plant physiological processes: what vegetation indices can and cannot tell us about the landscape. Sensors 2008, 8, 2136–2160. [Google Scholar] [CrossRef]
- Choudhury, B.J.; Ahmed, N.U.; Idso, S.B.; Reginato, R.J.; Daughtry, C.S.T. Relations between evaporation coefficients and vegetation indexes studied by model simulations. Remote Sens. Environ. 1994, 50, 1–17. [Google Scholar] [CrossRef]
- Allen, R.; Pereira, L.; Rais, D.; Smith, M. Crop Evapotranspiration—Guidelines for Computing Crop Water Requirements—FAO Irrigation and Drainage Paper 56; Food and Agriculture Organization of the United Nations: Rome, Italy, 1998. [Google Scholar]
- Mata-Gonzalez, R.; McLendon, T.; Martin, D.W. The inappropriate use of crop transpiration coefficients (Kc) to estimate evapotranspiration in arid ecosystems. Arid Land Res. Manag. 2005, 19, 285–295. [Google Scholar] [CrossRef]
- Xu, D.; Shen, Y. External and internal factors responsible for midday depression of photosynthesis. In Handbook of Photosynthesis, 2nd ed.; Pessarakli, M., Ed.; Taylor & Francis: Boca Raton, FL, USA, 2005; pp. 297–298. [Google Scholar]
- Gaskin, J.; Schaal, B. Hybrid Tamarix widespread in US invasion and undetected in native Asian range. P. Nat. Acad. Sci.USA 2002, 99, 11256–11259. [Google Scholar] [CrossRef] [PubMed]
- Glenn, E.; Nagler, P. Comparative ecophysiology of Tamarix ramosissima and native trees in western US riparian zones. J. Arid Envir. 2005, 61, 419–446. [Google Scholar] [CrossRef]
- Stromberg, J.; Chew, M.; Nagler, P.L.; Glenn, E.P. Change perceptions of change: the role of scientists in Tamarisk and river management. Restor. Ecol. 2009, 17, 177–186. [Google Scholar] [CrossRef]
- Di Tomaso, J. Impact, biology, and ecology of saltcedar (Tamarix spp.) in the southwestern United States. Weed Technol. 1998, 12, 326–336. [Google Scholar]
- Zavaleta, E. The economic value of controlling an invasive shrub. Ambio 2000, 29, 462–467. [Google Scholar] [CrossRef]
- United States 109th Congress. HR 2720: Salt Cedar and Russian Olive Control Demonstration Act; United State Congress: Washington, DC, USA, 2009. [Google Scholar]
- Devitt, D.; Sala, A.; Smith, S.; Cleverly, J.; Shaulis, L.; Hammett, R. Bowen ratio estimates of evapotranspiration for Tamarix ramosissima stands on the Virgin River in southern Nevada. Water Resour. Res. 1998, 34, 2407–2414. [Google Scholar] [CrossRef]
- Cleverly, J.; Dahm, C.; Thibault, J.; McDonnell, D.; Coonrod, J. Riparian ecohydrology: regulation of water flux from the ground to the atmosphere in the Middle Rio Grande, New Mexico. Hydrol. Process. 2006, 20, 3207–3225. [Google Scholar] [CrossRef]
- Westenberg, C.; Harper, D.; DeMeo, G. Evapotranspiration by Phreatophytes Along the Lower Colorado River at Havasu National Wildlife Refuge, Arizona; US Geological Survey Scientific Investigations Report, 2006-5043; USGS: Henderson, NV, USA, 2006.
- Sala, A.; Smith, S.; Devitt, D. Water use by Tamarix ramosissima and associated phreatophytes in a Mojave Desert floodplain. Ecol. Appl. 1996, 6, 888–898. [Google Scholar] [CrossRef]
- Owens, M.; Moore, G. Saltcedar water use: Realistic and unrealistic expectations. Rangeland Ecol. Manag. 2007, 60, 553–557. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriquez, E.; Gao, X.; Ferreira, L. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Nagler, P.; Glenn, E.; Didan, K.; Osterberg, J.; Jordan, F.; Cunningham, J. Wide-area estimates of stand structure and water use of Tamarix spp. on the Lower Colorado River: implications for restoration and water management projects. Restor. Ecol. 2009, 16, 136–145. [Google Scholar] [CrossRef]
- AZMET. The Arizona Meteorological Network; University of Arizona: Tucson, AZ, USA, 2009. Available online: http://cals.arizona.edu/azmet/ (accessed on November 18, 2009).
- Grime, V.; Sinclair, F. Sources of error in stem heat balance sap flow measurements. Agr. Forest Meteor. 1999, 94, 103–121. [Google Scholar] [CrossRef]
- Kjelgaard, J.; Stockle, C.; Black, R.; Campbell, G. Measuring sap flow with the heat balance approach using constant and variable heat inputs. Agr. Forest Meteor. 1997, 85, 239–250. [Google Scholar] [CrossRef]
- Nagler, P.; Glenn, E.; Thompson, T. Comparison of transpiration rates among saltcedar, cottonwood and willow trees by sap flow and canopy temperature methods. Agr. Forest Meteor. 2003, 116, 73–89. [Google Scholar] [CrossRef]
- Nagler, P.; Jetton, A.; Fleming, J.; Didan, K.; Glenn, E.; Erker, J.; Morino, K.; Milliken, J.; Gloss, S. Evapotranspiration in a cottonwood (Populus fremontii) restoration plantation estimated by sap flow and remote sensing methods. Agr, Forest Meteor. 2007, 144, 95–110. [Google Scholar] [CrossRef]
- Scott, R.; Huxman, T.; Cable, W.; Emmerich, W. Partitioning of evapotranspiration and its relation to carbon dioxide exchange in a Chihuahuan Desert shrubland. Hydrol. Process. 2006, 20, 3227–3243. [Google Scholar] [CrossRef]
- Phillips, N.; Oren, R. A comparison of daily representations of canopy conductance based on two conditional time-averaging methods and the dependence of daily conductance on environmental factors. Ann. Sci. Forest. 1998, 55, 217–235. [Google Scholar] [CrossRef]
- Ewers, B.; Oren, R. Analyses of assumptions and errors in the calculation of stomatal conductance from sap flux measurements. Tree Physiol. 2000, 20, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Moore, G.; Cleverly, J.; Owens, M. Nocturnal transpiration in riparian Tamarix thickets authenticated by sap flux, eddy covariance and leaf gas exchange measurements. Tree Physiol. 2008, 28, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Nagler, P.; Glenn, E.; Thompson, T.; Huete, A. Leaf area index and Normalized Difference Vegetation Index as predictors of canopy characteristics and light interception by riparian species on the Lower Colorado River. Agr. Forest Meteor. 2004, 116, 103–112. [Google Scholar] [CrossRef]
- Bell, J.P. Neutron Probe Practice. Institute of Hydrology Report 19; Institute of Hydrology: Oxfordshire, UK, 1987. [Google Scholar]
- Glenn, E.P.; McKeon, C.; Gerhart, V.; Nagler, P.L.; Jordan, F.; Artiola, J. Deficit irrigation of a landscape halophyte for reuse of saline waste water in a desert city. Landscape Urban Plan. 2009, 89, 57–64. [Google Scholar] [CrossRef]
- Oak Ridge National Laboratory Distributed Active Archive Center (ORNL DAAC). MODIS Subsetted Land Products, Collection 5; ORNL DAAC: Oak Ridge, TN, USA. Available online: www.daac.ornl/MODIS/modis.html (accessed on November 18, 2009).
- Monteith, J.; Unsworth, M. Principles of Environmental Physics, 2nd ed.; Edward Arnold: London, UK, 1990. [Google Scholar]
- Brouwer, C.; Heibloem, M. Irrigation Water Management Training Manual No. 3; FAO: Rome, Italy, 1986. [Google Scholar]
- Nagler, P.L.; Cleverly, J.; Glenn, E.; Pampkin, D.; Huete, A.; Wan, Z.M. Predicting riparian evapotranspiration from MODIS vegetation indices and meteorological data. Remote Sens. Environ. 2005, 94, 17–30. [Google Scholar] [CrossRef]
- Crago, R.D. Conservation and validity of the evaporative fraction during the daytime. J. Hydrol. 1996, 180, 173–180. [Google Scholar] [CrossRef]
- Chavez, J.L.; Neale, C.M.; Prueger, J.H.; Kustas, W.P. Daily evapotranspiration estimates from extrapolating instantaneous airborne remote sensing ET values. Irrig. Sci. 2008, 27, 67–81. [Google Scholar] [CrossRef]
- Wilson, K.; Balodocchi, D.; Falge, E.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Dolman, H.; Field, C.; Goldstein, A.; Granier, A.; Hollinger, D.; Katul, G.; Law, B.; Meyers, T.; Moncreiff, J.; Monson, R.; Tenhunen, J.; Valentini, R.; Verma, S.; Wofsy, S. Diurnal centroid of ecosystem energy and carbon fluxes at FLUXNET sites. J. Geophys. Res.-Atmos. 2003, 108, Art. No. 4664. [Google Scholar] [CrossRef]
- Mackay, D.S.; Samanta, S.; Nemani, R.R.; Band, L.E. Multi-objective parameter estimation for simulating canopy transpiration in forested watersheds. J. Hydrol. 2003, 277, 230–247. [Google Scholar] [CrossRef]
- Bevington, P. Data Reduction and Error Analysis for the Physical Sciences; McGraw-Hill Inc.: New York, NY, USA, 1969. [Google Scholar]
- GraphPad Inc. GraphPad QuickCalcs. Available online: www.graphpad.com/quickcalcs/ErrorPropplusminus/cfm (accessed on November 18, 2009).
- Bosabalidis, A.M.; Thomson, W.W. Ultrastructural development and secretion in the salt-glands of Tamarix aphylla L. J. Ultra. Res. 1985, 92, 55–62. [Google Scholar] [CrossRef]
- Horton, J.; Hart, S.; Kolb, T. Physiological condition and water source use of Sonoran Desert riparian trees at the Bill Williams River, Arizona, USA. Isot. Environ. Health S. 2003, 39, 69–82. [Google Scholar]
- Horton, J.; Kolb, T.; Hart, S. Physiological response to groundwater depth varies among species and with river flow regulation. Ecol. App. 2001, 11, 1046–1059. [Google Scholar] [CrossRef]
- Horton, J.; Kolb, T.; Hart, S. Responses of riparian trees to interannual variation in ground water depth in a semi-arid river basin. Plant Cell Environ. 2001, 24, 293–304. [Google Scholar] [CrossRef]
- Hultine, K.; Koepke, D.; Pockman, W.; Fravolini, A.; Sperry, J.; Williams, D. Influence of soil texture on hydraulic properties and water relations of a dominant warm-desert phreatophyte. Tree Physiol. 2006, 26, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Sperry, J.; Adler, F.; Campbell, G.; Comstock, J. Limitation of plant water use by rhizosphere and xylem conductance: results from a model. Plant Cell Environ. 1998, 21, 347–359. [Google Scholar] [CrossRef]
- Snyder, K.; Richards, J.; Donovan, L. Night-time conductance in C3 and C4 species: do plants lose water at night? J. Exp. Bot. 2003, 54, 861–865. [Google Scholar] [CrossRef] [PubMed]
- Su, Z. The Surface Energy Balance System (SEBS) for estimation of turbulent heat fluxes. Hydrol. Earth Syst. S. 2002, 6, 85–99. [Google Scholar] [CrossRef]
- Allen, R.; Tasumi, M.; Trezza, R. Satellite-based energy balance for mapping evapotranspiration with internalized calibration (METRIC)—Model. J. Irrig. Drain. E-ASCE 2007, 133, 380–394. [Google Scholar] [CrossRef]
- Bastiaanssen, W.G.M.; Noordman, E.J.M.; Pelgrum, H.; Davids, G.; Thoreson, B.P.; Allen, R.G. SEBAL model with remotely sensed data to improve water-resources management under actual field conditions. J. Irrig. Drain. E-ASCE 2005, 131, 85–93. [Google Scholar] [CrossRef]
- Tamarisk Coalition. Independent Peer Review of Tamarisk and Russian Olive Evapotranspiration Colorado River Basin; Tamarisk Coalition: Grand Junction, CO, USA, 2009. Available online: http://www.tamariskcoalition.org/tamariskcoalition/PDF/ET%20Report%20FINAL%204-16-09%20(2).pdf (accessed on December 1, 2009).
- Murray, R.S.; Nagler, P.L.; Morino, K.; Glenn, E.P. An empirical algorithm for estimating agricultural and riparian evapotranspiration using MODIS Enhanced Vegetation Index and ground measurements of ET. II. Application to the Lower Colorado River, US. Remote Sens. 2009, 1, 1125–1138. [Google Scholar] [CrossRef]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nagler, P.L.; Morino, K.; Murray, R.S.; Osterberg, J.; Glenn, E.P. An Empirical Algorithm for Estimating Agricultural and Riparian Evapotranspiration Using MODIS Enhanced Vegetation Index and Ground Measurements of ET. I. Description of Method. Remote Sens. 2009, 1, 1273-1297. https://doi.org/10.3390/rs1041273
Nagler PL, Morino K, Murray RS, Osterberg J, Glenn EP. An Empirical Algorithm for Estimating Agricultural and Riparian Evapotranspiration Using MODIS Enhanced Vegetation Index and Ground Measurements of ET. I. Description of Method. Remote Sensing. 2009; 1(4):1273-1297. https://doi.org/10.3390/rs1041273
Chicago/Turabian StyleNagler, Pamela L., Kiyomi Morino, R. Scott Murray, John Osterberg, and Edward P. Glenn. 2009. "An Empirical Algorithm for Estimating Agricultural and Riparian Evapotranspiration Using MODIS Enhanced Vegetation Index and Ground Measurements of ET. I. Description of Method" Remote Sensing 1, no. 4: 1273-1297. https://doi.org/10.3390/rs1041273