The Impact of Climate, CO2 and Population on Regional Food and Water Resources in the 2050s

Abstract
:1. Introduction
2. Methodology

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | 2000 | 2050s |
|---|---|---|
| Global | 6.1 | 8.7 |
| Africa | 0.78 | 1.9 |
| Australia | 0.019 | 0.02 |
| Asia | 3.7 | 4.8 |
| Europe | 0.77 | 0.85 |
| North America | 0.36 | 0.49 |
| South America | 0.42 | 0.64 |

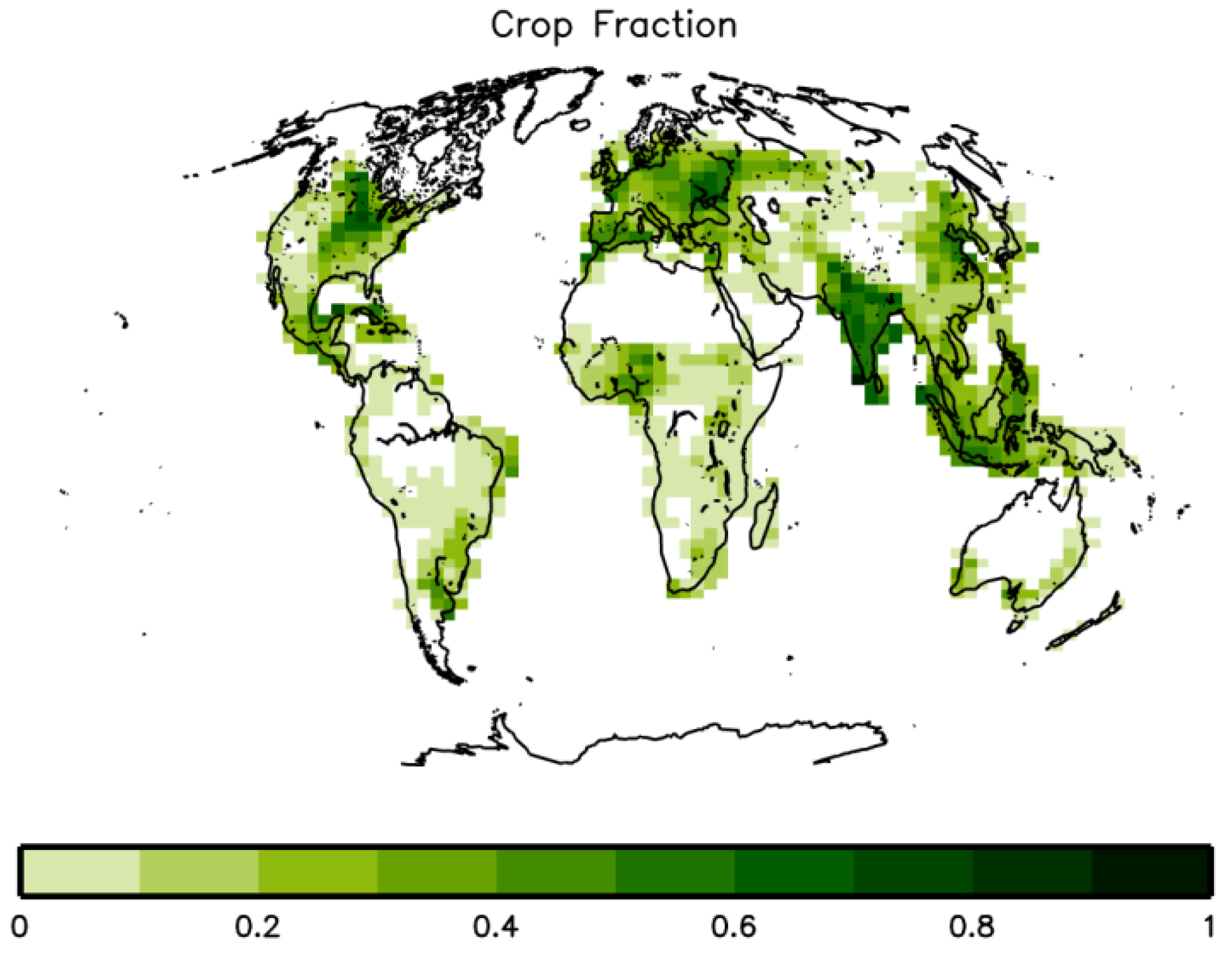
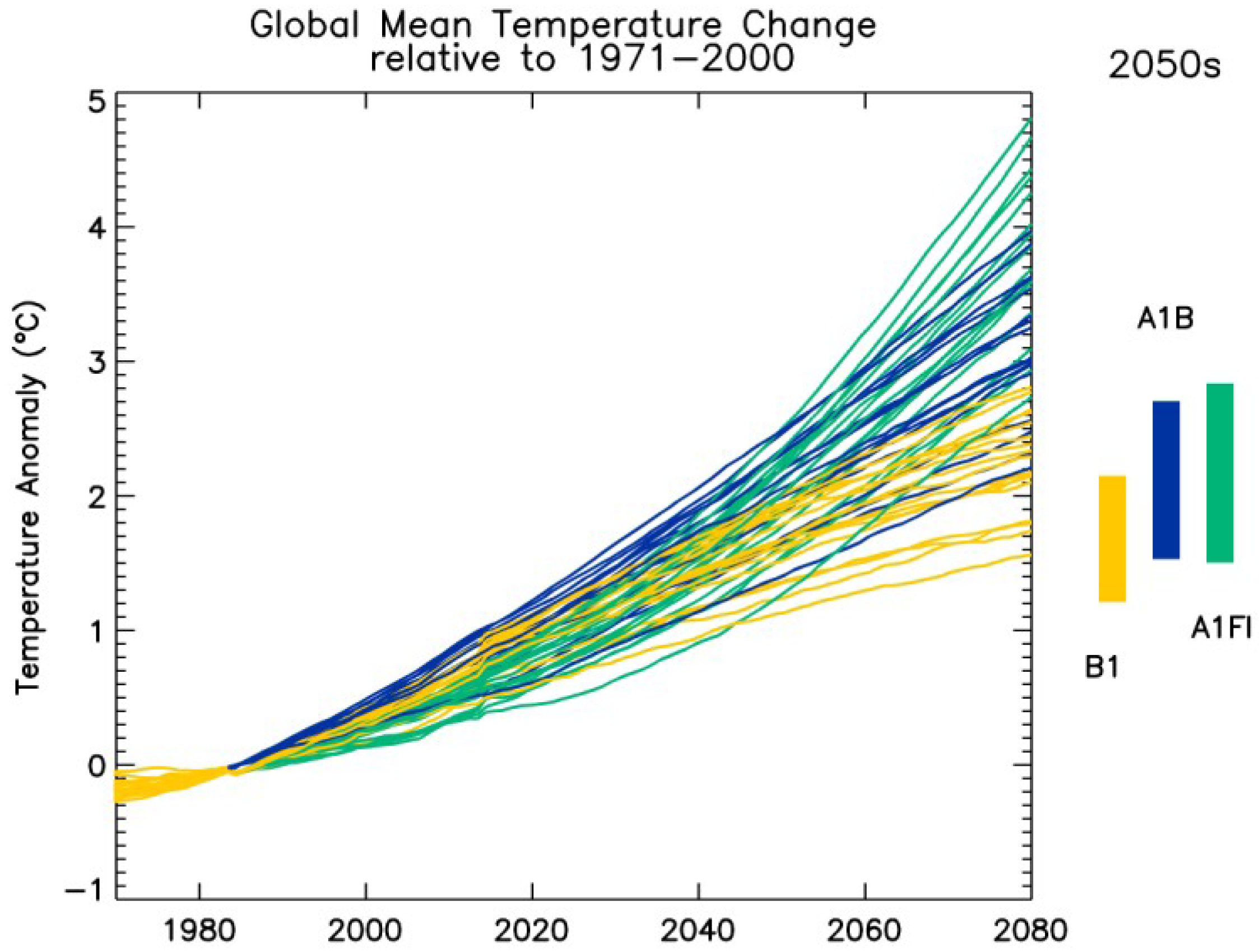
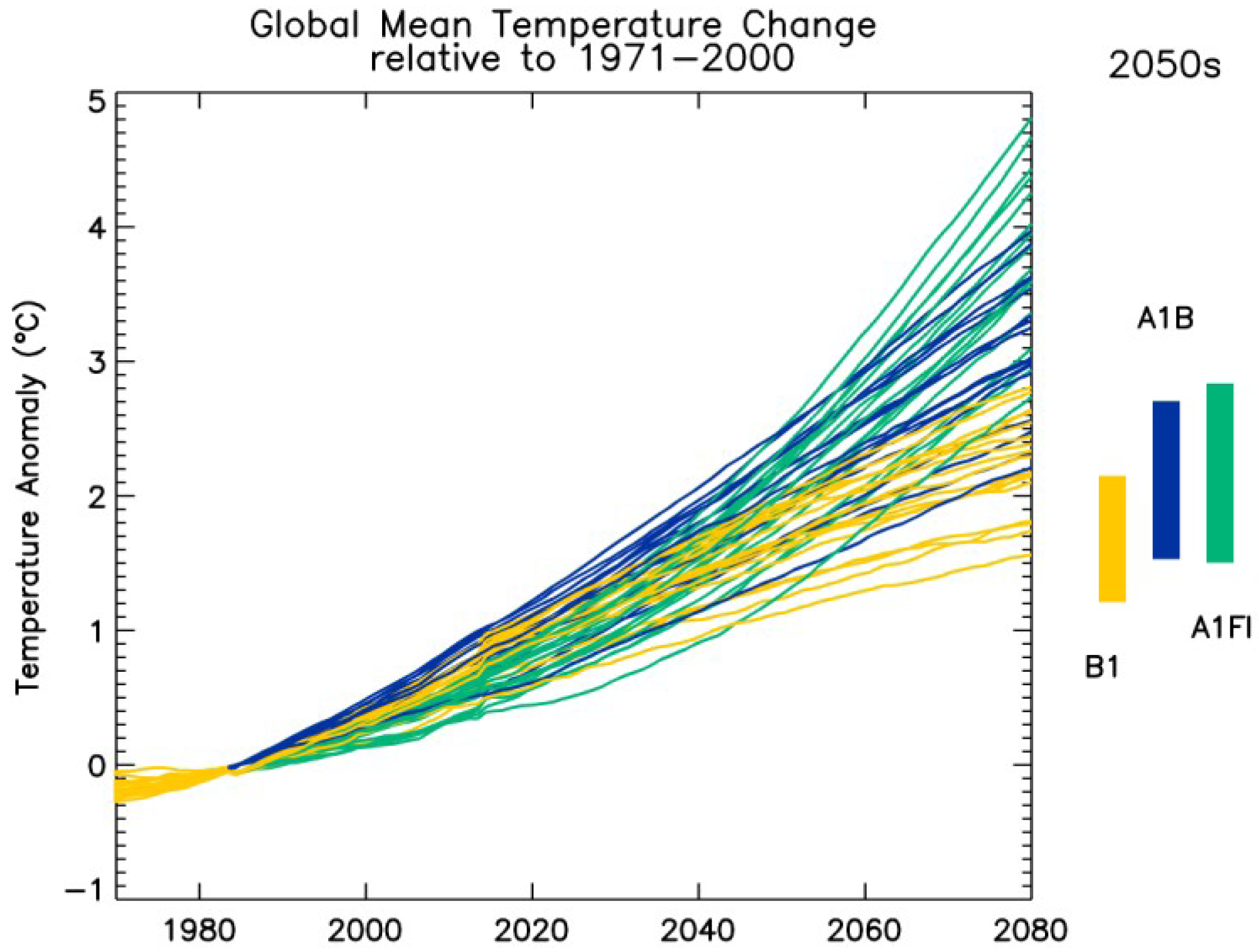
2.1. GCM Ensemble
2.2. CO2 Fertilisation and Physiological Forcing
2.3. Runoff and Water Resources
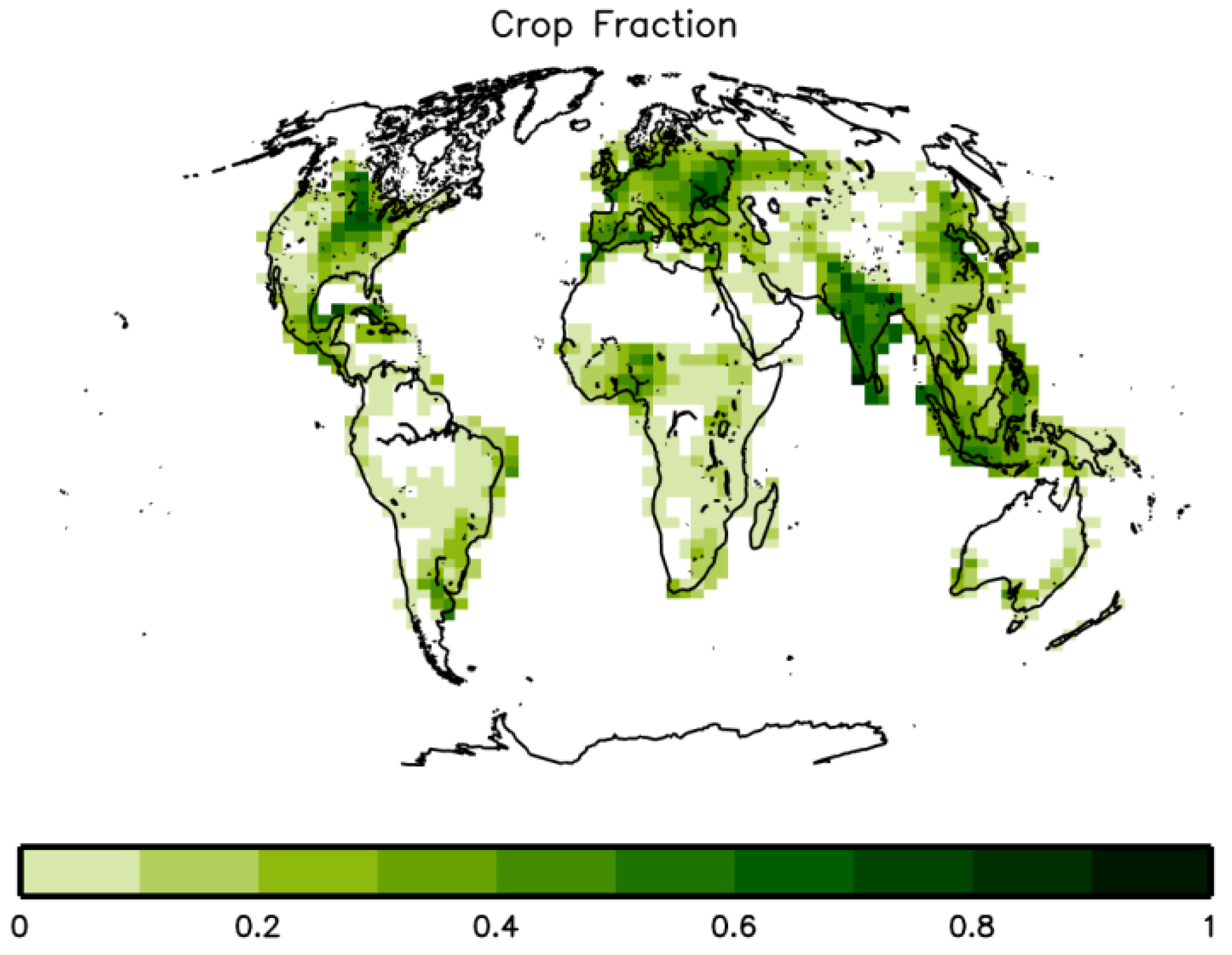
2.4. Crop Productivity and Food Availability
3. Results
| Region | Crop total NPP (Gt C) | Crop NPP (kg C per capita) |
|---|---|---|
| Global | 10.7 | 1,772 |
| Africa | 1.79 | 2,290 |
| Australia | 0.056 | 2,942 |
| Asia | 4.83 | 1,290 |
| Europe | 1.09 | 1,410 |
| North America | 1.18 | 3,323 |
| South America | 1.8 | 4,313 |
| Region | Water Stressed | High Water Stress | Extreme Water Stress |
|---|---|---|---|
| Global | 3.13 | 2.67 | 1.56 |
| Africa | 0.29 | 0.19 | 0.12 |
| Australia | 0.013 | 0.0066 | 0.0063 |
| Asia | 2.09 | 1.97 | 1.11 |
| Europe | 0.49 | 0.29 | 0.19 |
| North America | 0.067 | 0.06 | 0.035 |
| South America | 0.15 | 0.12 | 0.071 |
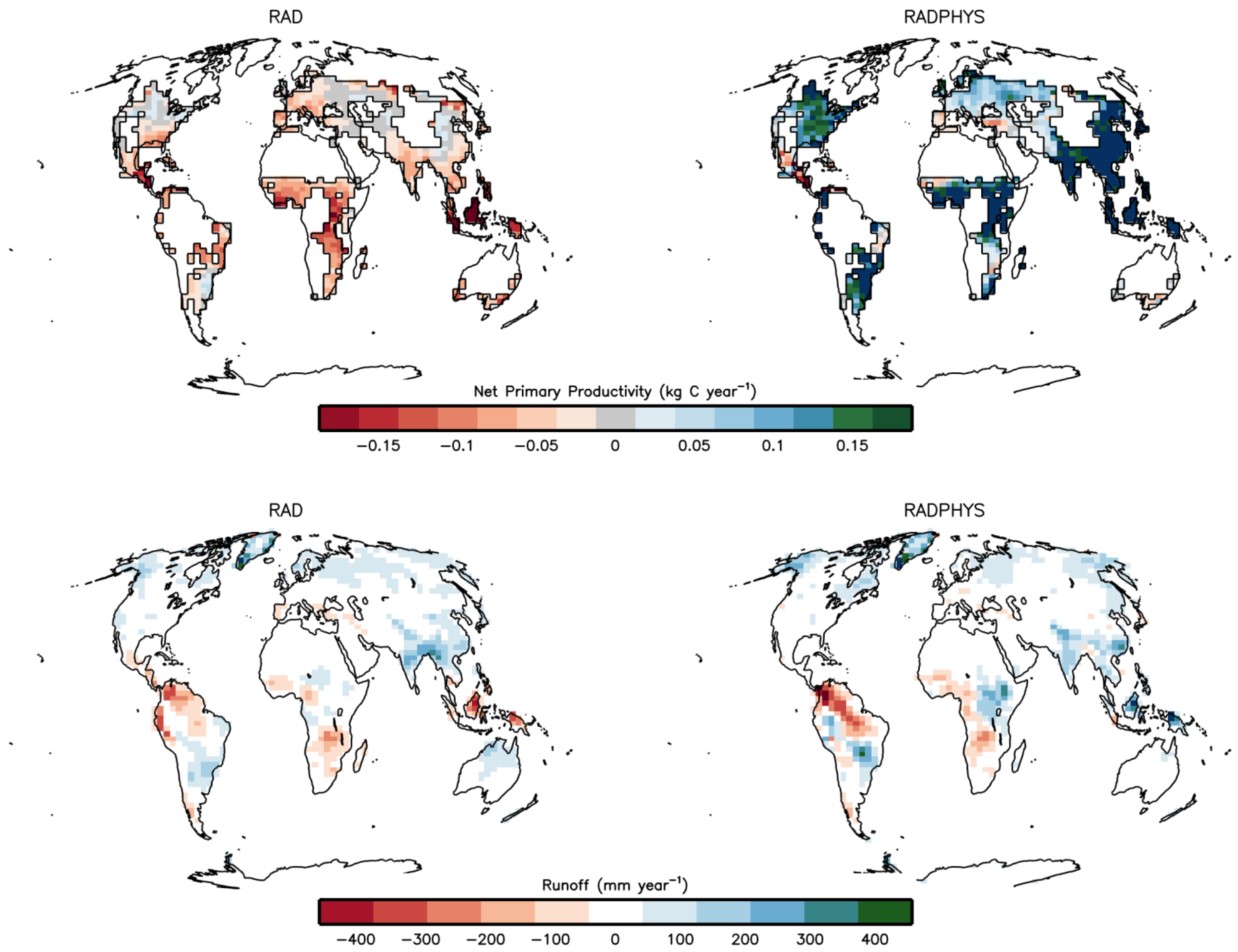
3.1. Climate Effect on Crop Productivity and Runoff
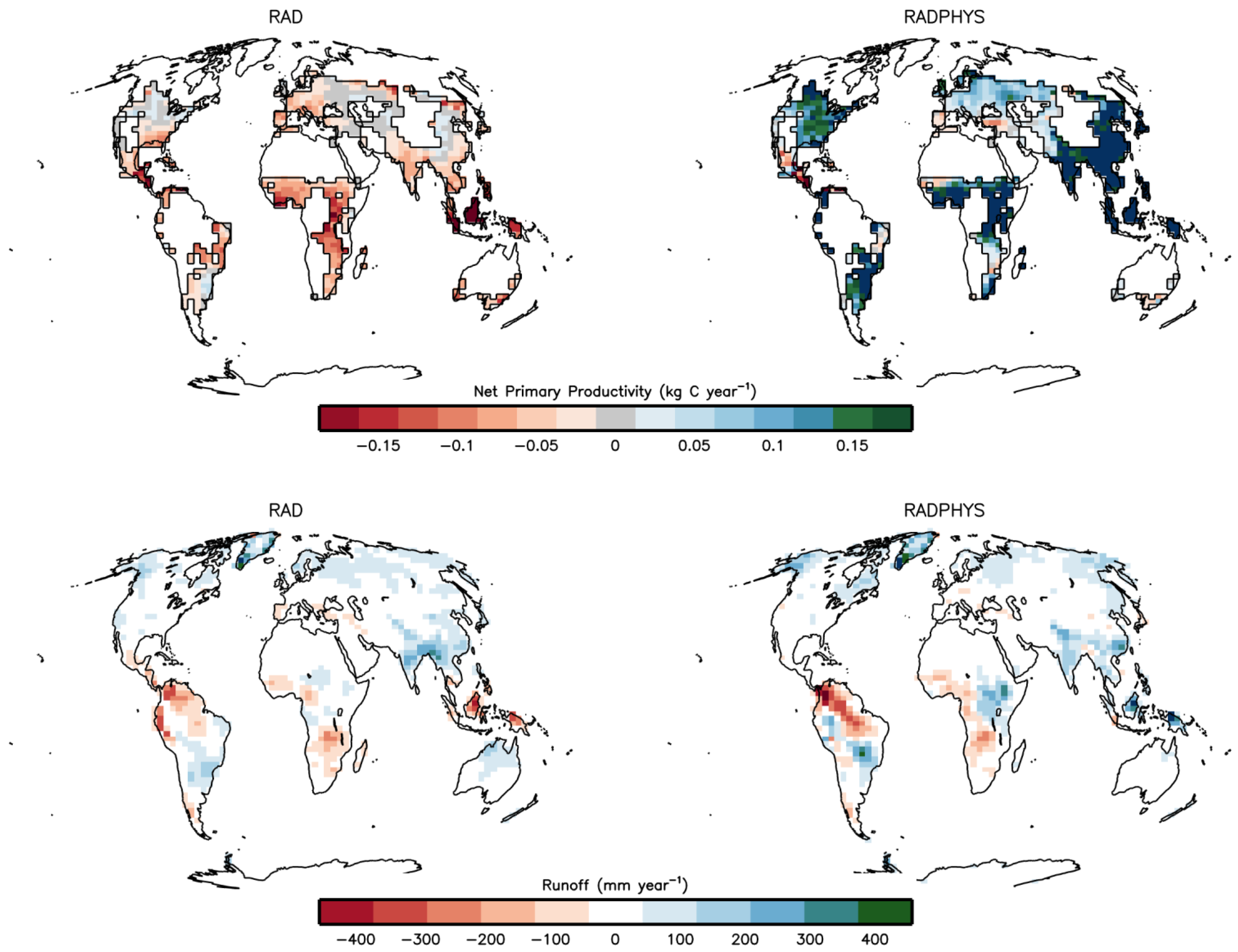
| Region | Cropland Production Change (Gt C per year) | Runoff Change (m3 s−1) | ||
|---|---|---|---|---|
| RAD | RADPHYS | RAD | RADPHYS | |
| Global | −0.67 | 2.14 | 68,097 | 108,225 |
| Africa | −0.14 | 0.39 | −9,776 | 5,900 |
| Australia | −0.016 | 0.003 | 2,670 | 619 |
| Asia | −0.25 | 1.09 | 40,309 | 55,839 |
| Europe | −0.053 | 0.14 | 1,618 | 2,950 |
| North America | −0.026 | 0.29 | 34,214 | 43,482 |
| South America | −0.1 | 0.16 | −13,781 | −413 |
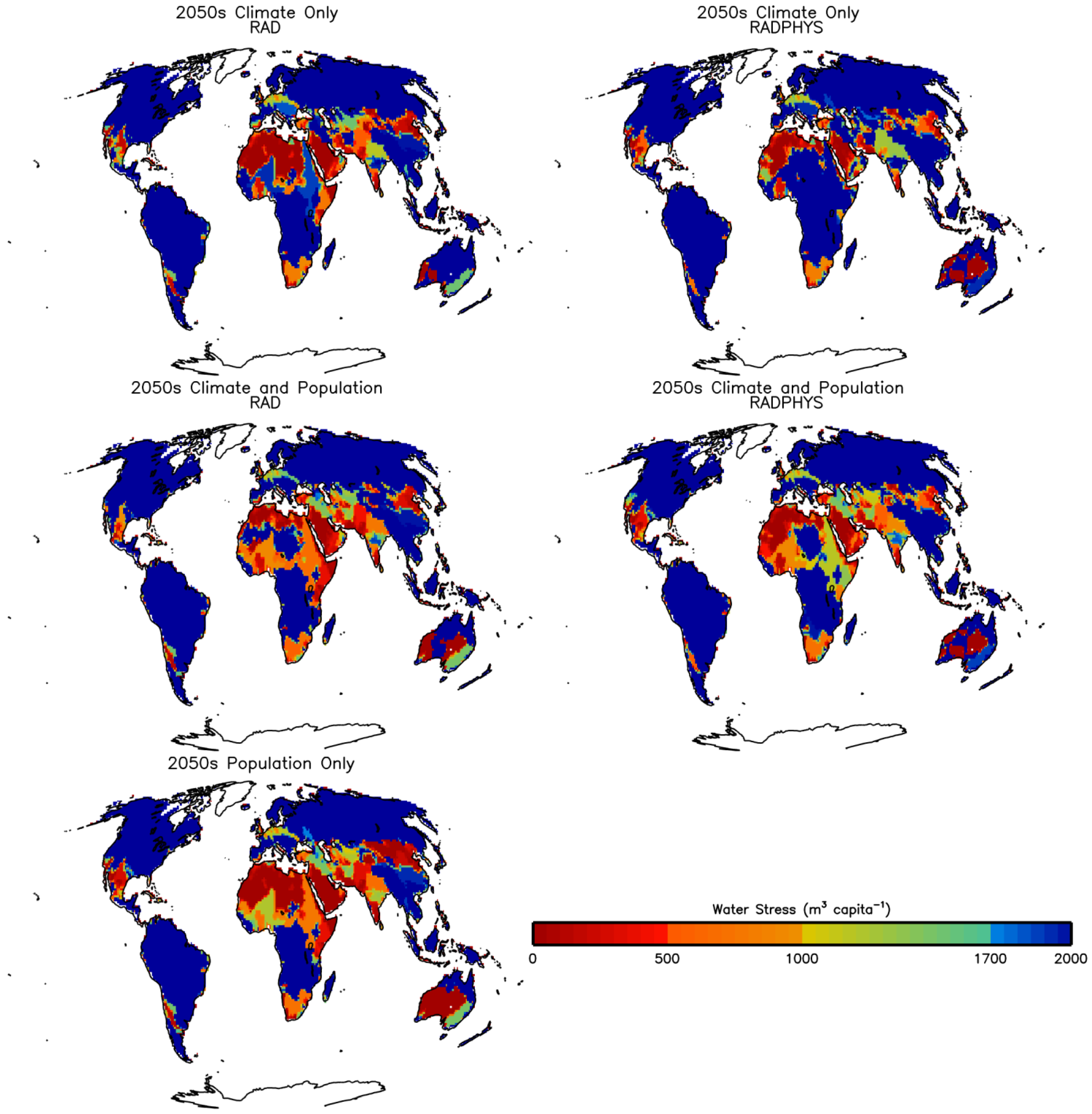
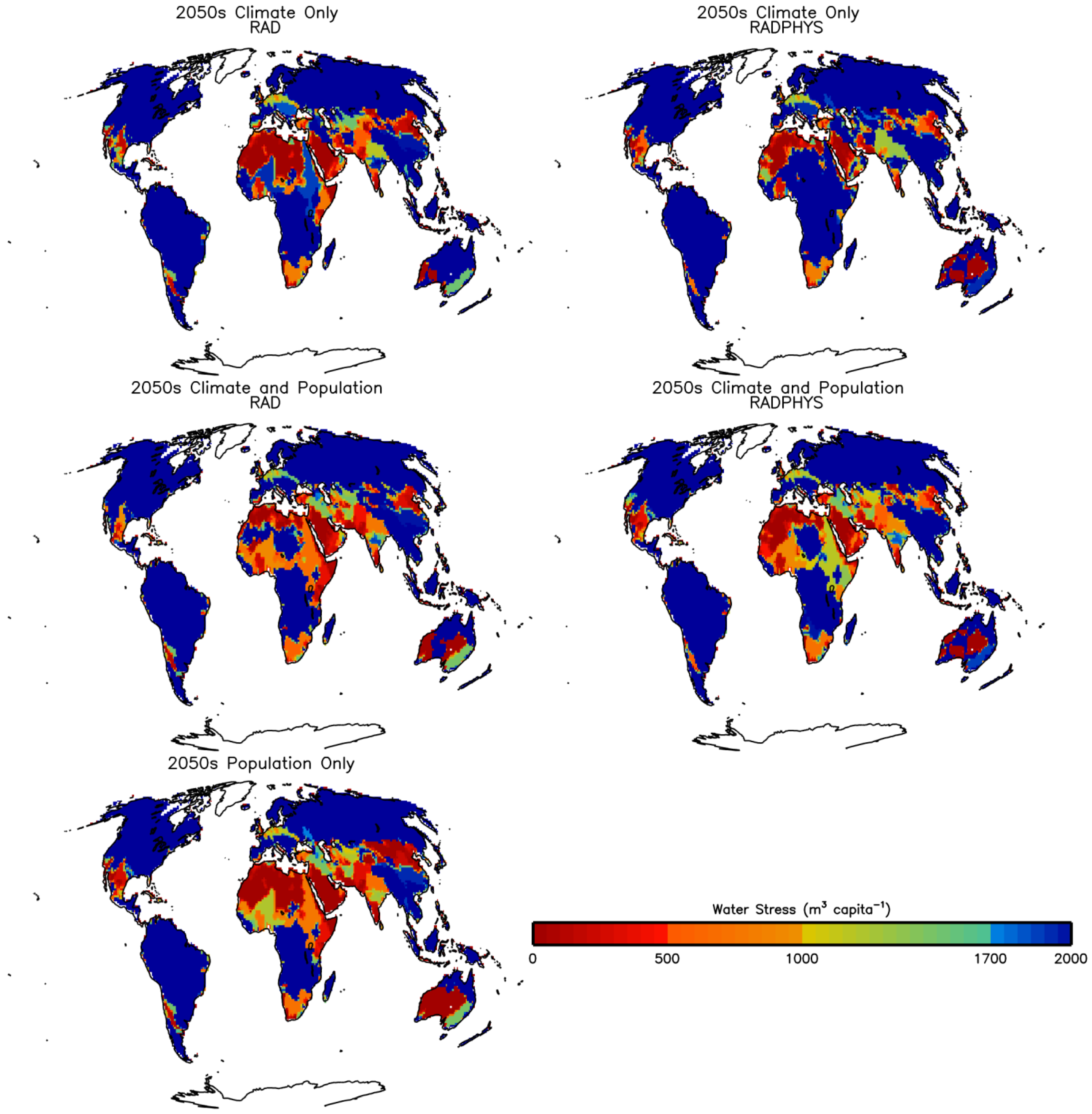
3.2. Future Impacts on Food and Water Availability
3.2.1. Climate Change
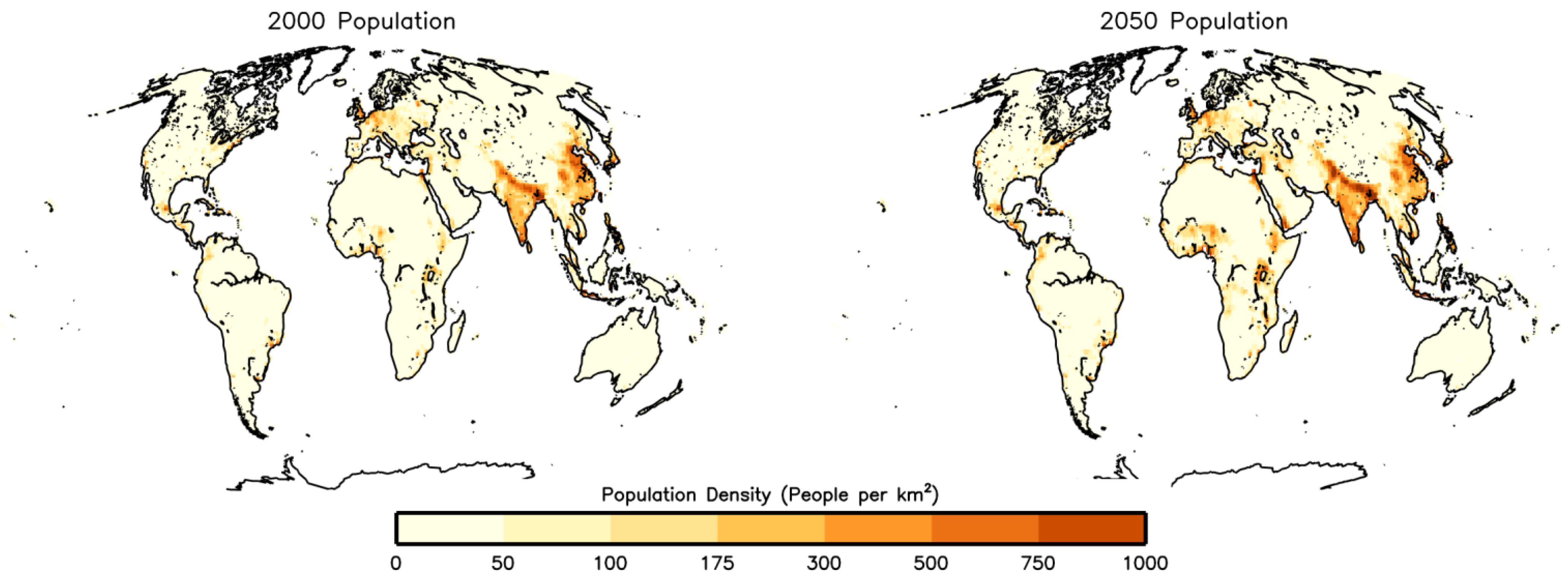
3.2.2. Population Change
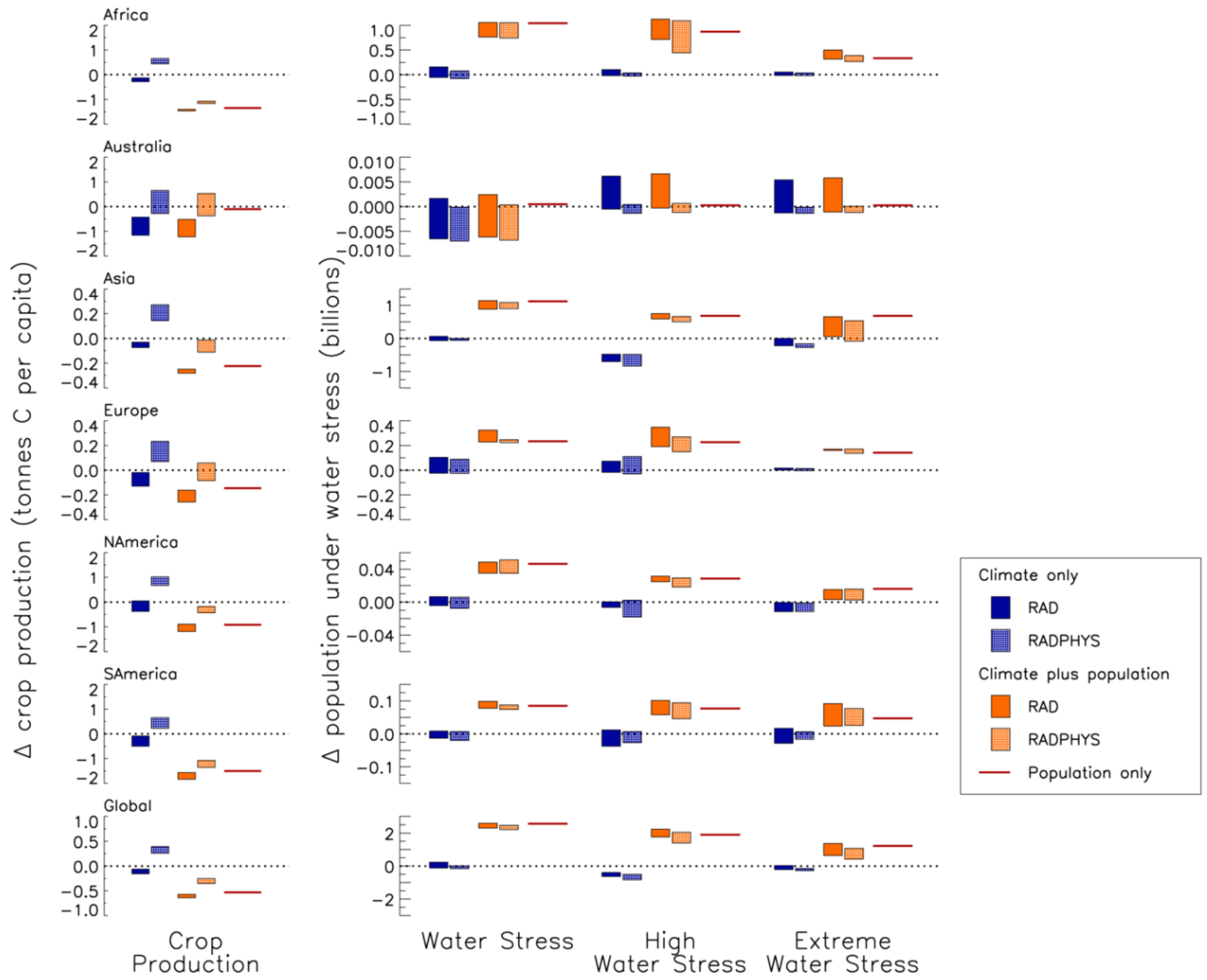
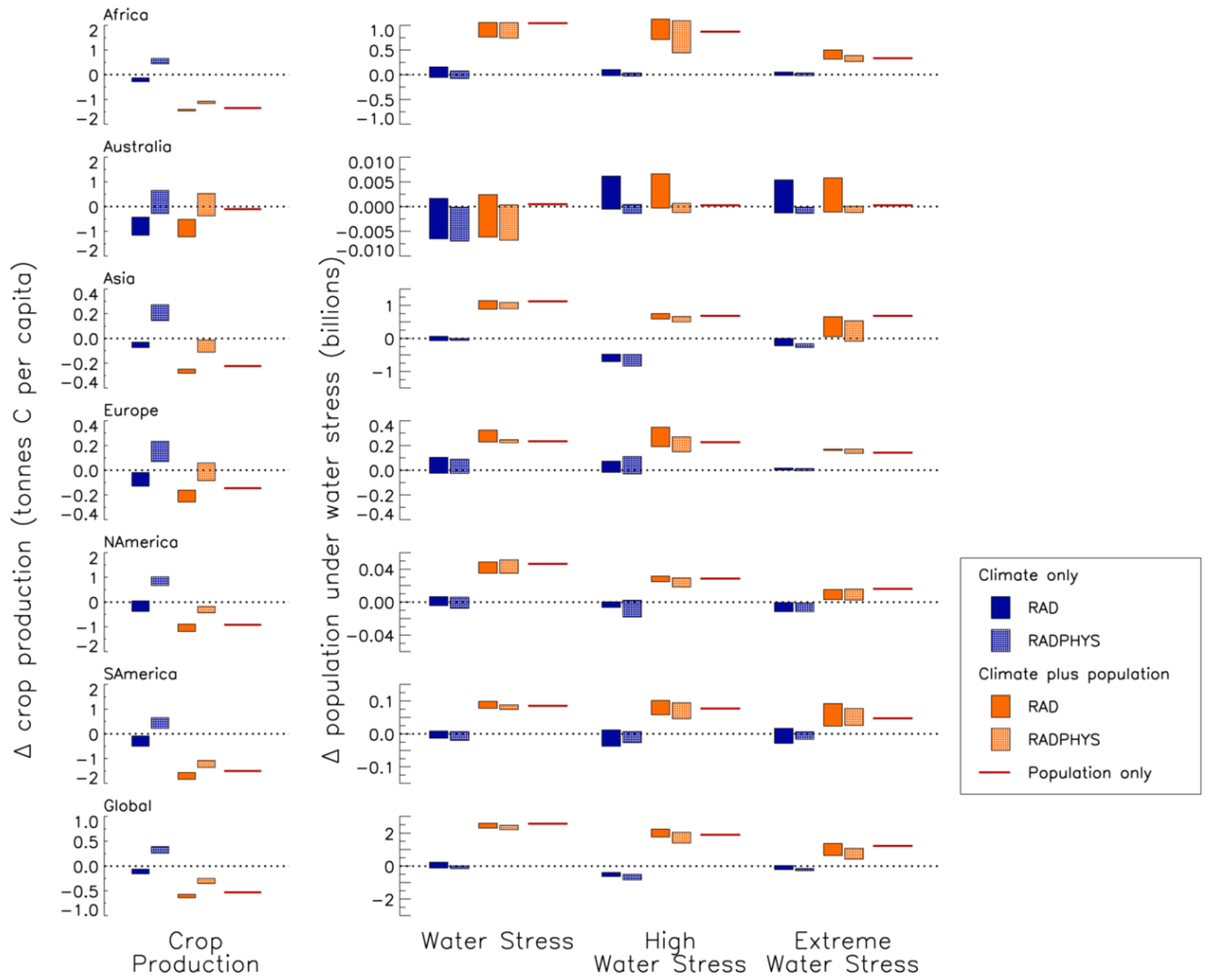
| Region | Climate only | Climate plus population | Population only | ||
|---|---|---|---|---|---|
| RAD | RADPHYS | RAD | RADPHYS | ||
| Global | −0.11 | 0.35 | −0.61 | −0.28 | −0.53 |
| Africa | −0.19 | 0.50 | −1.42 | −1.14 | −1.35 |
| Australia | −0.86 | 0.15 | −0.93 | 0.050 | −0.10 |
| Asia | −0.068 | 0.29 | −0.33 | −0.053 | −0.28 |
| Europe | −0.068 | 0.18 | −0.24 | −0.021 | −0.18 |
| North America | −0.074 | 0.81 | −0.98 | −0.33 | −0.92 |
| South America | −0.25 | 0.40 | −1.67 | −1.25 | −1.51 |
| Region | Water stress level | Climate only | Climate plus population | Population only | ||
|---|---|---|---|---|---|---|
| RAD | RADPHYS | RAD | RADPHYS | |||
| Global | Water Stressed | 0.12 | −0.073 | 2.41 | 2.32 | 2.54 |
| High Water Stress | −0.48 | −0.69 | 1.95 | 1.55 | 1.90 | |
| Extreme Water Stress | −0.11 | −0.24 | 1.16 | 1.03 | 1.23 | |
| Africa | Water Stressed | 0.053 | −0.043 | 1.04 | 1.00 | 1.04 |
| High Water Stress | 0.011 | −0.007 | 1.02 | 0.63 | 0.87 | |
| Extreme Water Stress | 0.0079 | −0.0017 | 0.37 | 0.29 | 0.34 | |
| Australia | Water Stressed | −0.0003 | −0.0062 | 0.0002 | −0.0057 | 0.0005 |
| High Water Stress | 0.0005 | −0.0004 | 0.0008 | −0.0002 | 0.0002 | |
| Extreme Water Stress | −0.0003 | −0.0005 | 0.0000 | −0.0003 | 0.0002 | |
| Asia | Water Stressed | −0.014 | −0.039 | 1.08 | 1.02 | 1.13 |
| High Water Stress | −0.49 | −0.70 | 0.66 | 0.64 | 0.68 | |
| Extreme Water Stress | −0.09 | −0.22 | 0.55 | 0.50 | 0.68 | |
| Europe | Water Stressed | 0.004 | −0.018 | 0.24 | 0.23 | 0.23 |
| High Water Stress | 0.018 | −0.0042 | 0.24 | 0.20 | 0.23 | |
| Extreme Water Stress | 0.014 | 0.0062 | 0.17 | 0.15 | 0.14 | |
| North America | Water Stressed | −0.0010 | −0.0008 | 0.040 | 0.036 | 0.046 |
| High Water Stress | −0.0005 | −0.0034 | 0.028 | 0.027 | 0.029 | |
| Extreme Water Stress | −0.0036 | −0.0090 | 0.013 | 0.0081 | 0.016 | |
| South America | Water Stressed | 0.0054 | −0.0010 | 0.087 | 0.084 | 0.085 |
| High Water Stress | −0.0002 | 0.0010 | 0.088 | 0.078 | 0.077 | |
| Extreme Water Stress | 0.0026 | −0.0042 | 0.065 | 0.058 | 0.047 | |
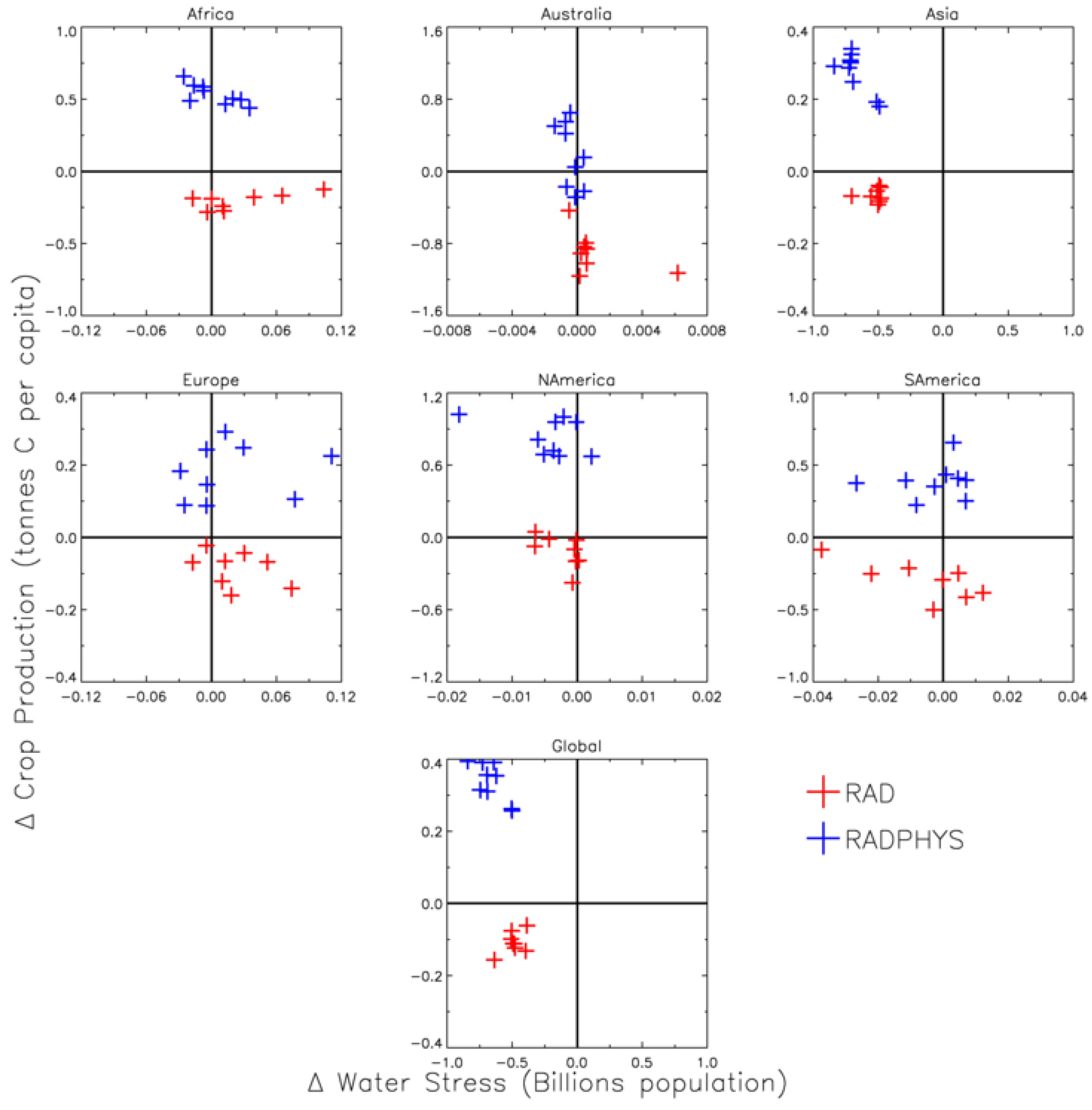
3.2.3. Climate, CO2 and Population Change
3.3. CO2 Implications for Food and Water Resources
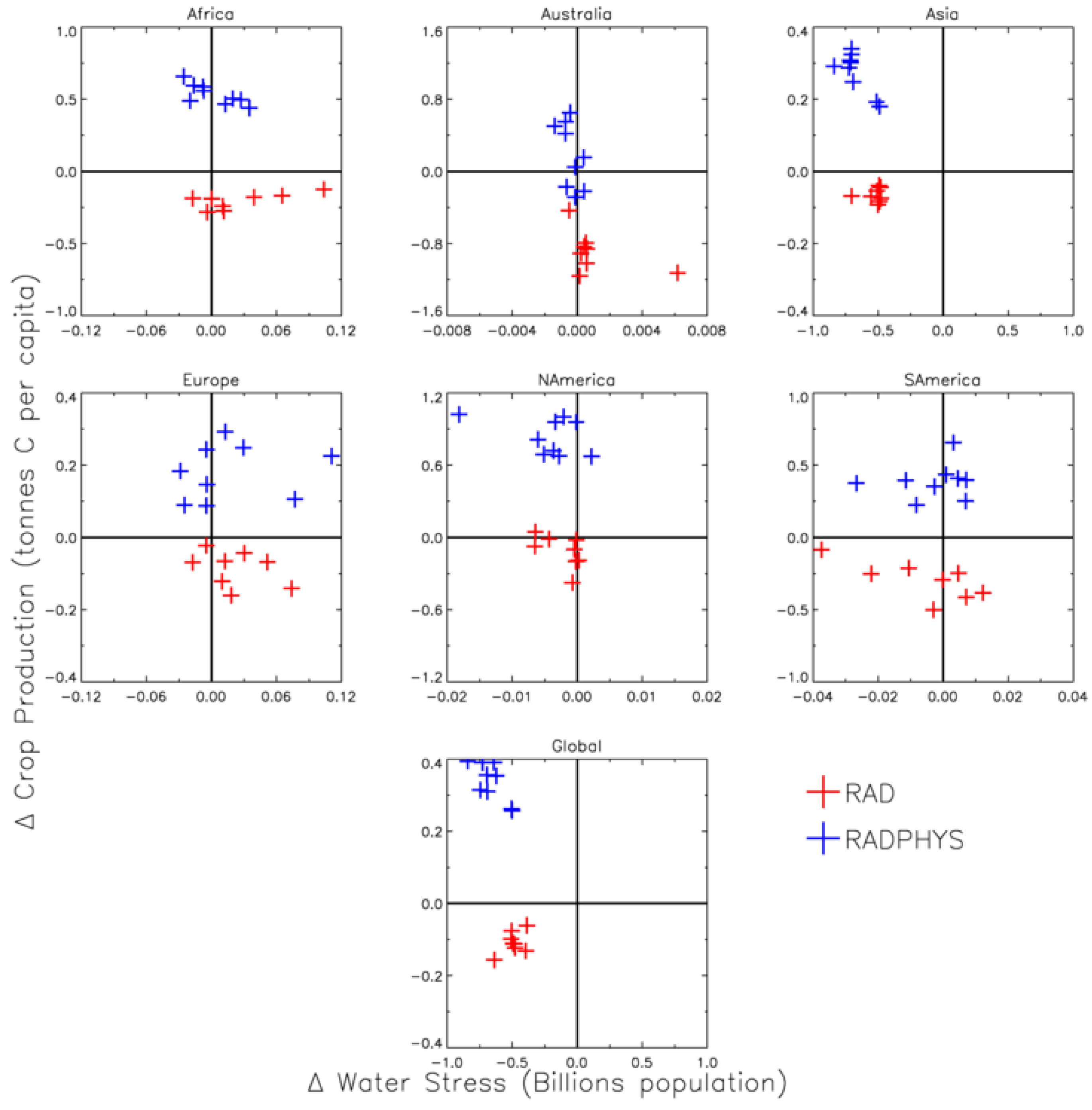
4. Discussion
Carbon Dioxide
5. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Meadows, D.H.; Meadows, D.L.; Randers, J.; Behrens, W.W., III. The Limits to Growth: A Report for the Club of Rome’s Project on the Predicament of Mankind; Universe Books: New York, NY, USA, 1972. [Google Scholar]
- Beddington, J.R.; Asaduzzaman, M.; Clark, M.E.; Bremauntz, A.F.; Guillou, M.D.; Howlett, D.J.B.; Jahn, M.M.; Lin, E.; Mamo, T.; Negra, C.; et al. What next for agriculture after durban? Science 2012, 335, 289–290. [Google Scholar]
- Molden, D.; Oweis, T.Y.; Steduto, P.; Kijne, J.W.; Hanjra, M.A.; Bindraban, P.S.; Bouman, B.A.M.; Cook, S.; Erenstein, O.; Farahani, H.; et al. Pathways for increasing agricultural water productivity. In Agriculture, Water for Food, Water for Life: A Comprehensive Assessment of Water Management in Agriculture; Molden, D., Ed.; International Water Management Institute: London, UK, 2007. [Google Scholar]
- Hanjra, M.A.; Qureshi, M.E. Global water crisis and future food security in an era of climate change. Food Policy 2010, 35, 365–377. [Google Scholar] [CrossRef]
- Molden, D.; Oweis, T.; Steduto, P.; Bindraban, P.; Hanjra, M.A.; Kijne, J. Improving agricultural water productivity: Between optimism and caution. Agric. Water Manage 2010, 97, 528–535. [Google Scholar] [CrossRef]
- Rosenzweig, C.; Parry, M.L. Potential impact of climate change on world food supply. Nature 1994, 367, 133–138. [Google Scholar] [CrossRef]
- World Water Assessment Programme, The United Nations World Water Development Report 3: Water in a Changing World; UNESCO: London, UK, 2009.
- Barriopedro, D.; Fischer, E.M.; Luterbacher, J.; Trigo, R.; Garcia-Herrera, R. The Hot summer of 2010: Redrawing the Temperature record map of europe. Science 2011, 332, 220–224. [Google Scholar] [CrossRef]
- Schmidhuber, J.; Tubiello, F.N. Global food security under climate change. Proc. Natl. Acad. Sci. USA 2007, 104, 19703–19708. [Google Scholar] [CrossRef]
- Pope, V.D.; Gallani, M.L.; Rowntree, P.R.; Stratton, R.A. The impact of new physical parametrizations in the Hadley Centre climate model: HadAM3. Clim. Dynam. 2000, 16, 123–146. [Google Scholar] [CrossRef]
- Gordon, C.; Cooper, C.; Senior, C.A.; Banks, H.; Gregory, J.M.; Johns, T.C.; Mitchell, J.F.B.; Wood, R.A. The simulation of SST, sea ice extents and ocean heat transports in a version of the Hadley Centre coupled model without flux adjustments. Clim. Dynam. 2000, 16, 147–168. [Google Scholar] [CrossRef]
- Gornall, J.; Betts, R.; Burke, E.; Clark, R.; Camp, J.; Willett, K.; Wiltshire, A. Implications of climate change for agricultural productivity in the early twenty-first century. Philos Trans. R. Soc. B 2010, 365, 2973–2989. [Google Scholar] [CrossRef]
- Nakicenovic, N.; Alcamo, J.; Davis, G.; de Vries, B.; Fenhann, J.; Gaffin, S.; Gregory, K.; Grübler, A.; Jung, T.Y.; Kram, T.; et al. IPCC Special Report on Emissions Scenarios; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Collins, M.; Booth, B.B.B.; Harris, G.R.; Murphy, J.M.; Sexton, D.M.H.; Webb, M.J. Towards quantifying uncertainty in transient climate change. Clim. Dynam. 2006, 27, 127–147. [Google Scholar] [CrossRef]
- Lutz, W.; Sanderson, W.; Scherbov, S. The coming acceleration of global population ageing. Nature 2008, 451, 716–719. [Google Scholar] [CrossRef]
- Lutz, W.; Sanderson, W.; Scherbov, S. IIASA’s 2007 Probabilistic World Population Projections, IIASA World Population Program Online Data Base of Results. Available online: http://www.iiasa.ac.at/Research/POP/proj07/index.html?sb=5/ (accessed on 1 February 2012).
- Van Vuuren, D.P.; Lucas, P.L.; Hilderink, H. Downscaling drivers of global environmental change: Enabling use of global SRES scenarios at the national and grid levels. Global Environ. Change 2007, 17, 114–130. [Google Scholar] [CrossRef]
- Ramankutty, N.; Evan, A.T.; Monfreda, C.; Foley, J.A. Farming the planet: 1. Geographic distribution of global agricultural lands in the year 2000. Glob. Biogeochem. Cycles 2008. [Google Scholar] [CrossRef]
- Murphy, J.M.; Sexton, D.M.H.; Barnett, D.N.; Jones, G.S.; Webb, M.J.; Collins, M. Quantification of modelling uncertainties in a large ensemble of climate change simulations. Nature 2004, 430, 768–772. [Google Scholar]
- Collins, M.C.M.; Booth, B.B.; Bhaskaran, B.; Harris, G.R.; Murphy, J.M.; Sexton, D.M.H.; Webb, M.J. Climate model errors, feedbacks and forcings: a comparison of perturbed physics and multi-model ensembles. Clim. Dynam. 2011, 36, 1737–1766. [Google Scholar] [CrossRef]
- Wiltshire, A.J.; Gornall, J.; Booth, B.B.B.; Dennis, E.; Falloon, P.; Kay, G.; McNeall, D.; Betts, R.A. The importance of population, climate change and CO2 plant physiological forcing in determining future global water stress. Global Environ. Change 2013. submitted. [Google Scholar]
- Field, C.B.; Jackson, R.B.; Mooney, H.A. Stomatal responses to increased CO2—Implications from the plant to the global-scale. Plant Cell Environ. 1995, 18, 1214–1225. [Google Scholar] [CrossRef]
- Jarvis, P.G.; McNaughton, K.G. Stomatal control of transpiration—Scaling up from leaf to region. Adv. Ecol. Res. 1986, 15, 1–49. [Google Scholar] [CrossRef]
- Bernacchi, C.J.; Kimball, B.A.; Quarles, D.R.; Long, S.P.; Ort, D.R. Decreases in stomatal conductance of soybean under open-air elevation of CO2 are closely coupled with decreases in ecosystem evapotranspiration. Plant Physiol. 2007, 143, 134–144. [Google Scholar]
- Hungate, B.A.; Reichstein, M.; Dijkstra, P.; Johnson, D.; Hymus, G.; Tenhunen, J.D.; Hinkle, C.R.; Drake, B.G. Evapotranspiration and soil water content in a scrub-oak woodland under carbon dioxide enrichment. Glob. Change Biol. 2002, 8, 289–298. [Google Scholar] [CrossRef]
- Li, J.H.; Erickson, J.E.; Peresta, G.; Drake, B.G. Evapotranspiration and water use efficiency in a Chesapeake Bay wetland under carbon dioxide enrichment. Glob. Change Biol. 2010, 16, 234–245. [Google Scholar] [CrossRef]
- Tubiello, F.N.; Amthor, J.S.; Boote, K.J.; Donatelli, M.; Easterling, W.; Fischer, G.; Gifford, R.M.; Howden, M.; Reilly, J.; Rosenzweig, C. Crop response to elevated CO2 and world food supply—A comment on “Food for Thought...” by Long et al., Science 312: 1918–1921, 2006. Eur. J. Agron. 2007, 26, 215–223. [Google Scholar]
- Ainsworth, E.A.; Long, S.P. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy. New Phytol. 2005, 165, 351–371. [Google Scholar] [CrossRef]
- Cox, P.M.; Huntingford, C.; Harding, R.J. A canopy conductance and photosynthesis model for use in a GCM land surface scheme. J. Hydrol. 1998, 213, 79–94. [Google Scholar] [CrossRef]
- Oki, T.; Sud, U.C. Design of Total Runoff Integrating Pathways (TRIP)—A global river channel network. Earth Interact. 1998, 2, 1–36. [Google Scholar]
- Fekete, B.M.; Vorosmarty, C.J.; Grabs, W. High-resolution fields of global runoff combining observed river discharge and simulated water balances. Glob. Biogeochem. Cycles 2002. [Google Scholar] [CrossRef]
- Falkenmark, M. Meeting water requirements of an expanding world population. Philos. Trans. R. Soc. B Biol. Sci. 1997, 353, 929–936. [Google Scholar] [CrossRef]
- Falkenmark, M.; Lundqvist, J.; Widstrand, C. Macro-scale water scarcity requires micro-scale approaches. Nat. Resour. Forum 1989, 13, 258–267. [Google Scholar] [CrossRef]
- Koster, R.D.; Dirmeyer, P.A.; Guo, Z.C.; Bonan, G.; Chan, E.; Cox, P.; Gordon, C.T.; Kanae, S.; Kowalczyk, E.; Lawrence, D.; et al. Regions of strong coupling between soil moisture and precipitation. Science 2004, 305, 1138–1140. [Google Scholar] [CrossRef]
- Harding, R.J.; Blyth, E.M.; Tuinenberg, O.; Wiltshire, A.J. Land atmosphere feedbacks and their role in the water resources of the Ganges basin. Sci. Total Environ. 2013, in press. [Google Scholar]
- Pijanowski, B.; Moore, N.; Mauree, D.; Niyogi, D. Evaluating error propagation in coupled land-atmosphere models. Earth Interact. 2011, 15, 1–25. [Google Scholar]
- Clark, D.B.; Mercado, L.M.; Sitch, S.; Jones, C.D.; Gedney, N.; Best, M.J.; Pryor, M.; Rooney, G.G.; Essery, R.L.H.; Blyth, E.; et al. The Joint UK Land Environment Simulator (JULES), model description—Part 2: Carbon fluxes and vegetation dynamics. Geosci. Model Dev. 2011, 4, 701–722. [Google Scholar] [CrossRef]
- Best, M.J.; Pryor, M.; Clark, D.B.; Rooney, G.G.; Essery, R.L.H.; Menard, C.B.; Edwards, J.M.; Hendry, M.A.; Porson, A.; Gedney, N.; et al. The Joint UK Land Environment Simulator (JULES), model description—Part 1: Energy and water fluxes. Geosci. Model Dev. 2011, 4, 677–699. [Google Scholar] [CrossRef]
- Weedon, G.P.; Gomes, S.; Viterbo, P.; Shuttleworth, W.J.; Blyth, E.; Österle, H.; Adam, J.C.; Bellouin, N.; Boucher, O.; Best, M. Creation of the WATCH Forcing Data and its use to assess global and regional reference crop evaporation over land during the twentieth century. J. Hydrometeorol. 2011, 12, 823–848. [Google Scholar] [CrossRef]
- Haberl, H.; Erb, K.H.; Krausmann, F.; Gaube, V.; Bondeau, A.; Plutzar, C.; Gingrich, S.; Lucht, W.; Fischer-Kowalski, M. Quantifying and mapping the human appropriation of net primary production in earth’s terrestrial ecosystems. Proc. Natl. Acad. Sci. USA 2007, 104, 12942–12945. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Ehrlich, P.R.; Ehrlich, A.H.; Matson, P.A. Human appropriation of the products of photosynthesis. Bioscience 1986, 36, 368–373. [Google Scholar] [CrossRef]
- Imhoff, M.L.; Bounoua, L.; Ricketts, T.; Loucks, C.; Harriss, R.; Lawrence, W.T. Global patterns in human consumption of net primary production. Nature 2004, 429, 870–873. [Google Scholar] [CrossRef]
- FAO. Food Balance Sheets—FAOSTAT. Available online: http://faostat.fao.org/default.aspx/ (accessed on 1 December 2012).
- Rakotoarisoa, M.A.; Lafrate, M.; Paschali, M. Why Has Africa Become a Net Food Importer? Explaining Africa Agricultural and Food Trade Deficits; Trade and Markets Division, Food and Agriculture Organization of the United Nations: Rome, Italy, 2011. [Google Scholar]
- Islam, M.S.; Oki, T.; Kanae, S.; Hanasaki, N.; Agata, Y.; Yoshimura, K. A grid-based assessment of global water scarcity including virtual water trading. Water Resour. Manage 2007, 21, 19–33. [Google Scholar]
- Arnell, N.W. Climate change and global water resources: SRES emissions and socio-economic scenarios. Global Environ. Change 2004, 14, 31–52. [Google Scholar] [CrossRef]
- Revenga, C.; Brunner, J.; Henniger, N.; Kassem, K.; Payne, R. Pilot Analysis of Global Ecosystems: Freshwater Systems; World Resources Institute: Washington, DC, USA, 2000. [Google Scholar]
- Kummu, M.; Ward, P.J.; de Moel, H.; Varis, O. Is physical water scarcity a new phenomenon? Global assessment of water shortage over the last two millennia. Environ. Res. Lett. 2010. [Google Scholar] [CrossRef]
- Hoff, H.; Falkenmark, M.; Gerten, D.; Gordon, L.; Karlberg, L.; Rockstrom, J. Greening the global water system. J. Hydrol. 2010, 384, 177–186. [Google Scholar] [CrossRef]
- Gornall, J.L.; Betts, R.A.; Wiltshire, A.J. Anthropogenic Drivers of Environmental Change. In Handbook of Environmental Change; Matthews, J., Ed.; SAGE: London, UK, 2012. [Google Scholar]
- Beddington, J. Food security: Contributions from science to a new and greener revolution. Philos. Trans. R. Soc. B 2010, 365, 61–71. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Pretty, J.; Thomas, S.M.; Warham, E.J.; Beddington, J.R. Linking policy on climate and food. Science 2011, 331, 1013–1014. [Google Scholar]
- Tilman, D.; Reich, P.B.; Knops, J.; Wedin, D.; Mielke, T.; Lehman, C. Diversity and productivity in a long-term grassland experiment. Science 2001, 294, 843–845. [Google Scholar] [CrossRef]
- Burke, M.B.; Lobell, D.B.; Guarino, L. Shifts in African crop climates by 2050, and the implications for crop improvement and genetic resources conservation. Global Environ. Change 2009, 19, 317–325. [Google Scholar] [CrossRef]
- Long, S.P.; Ainsworth, E.A.; Rogers, A.; Ort, D.R. Rising atmospheric carbon dioxide: Plants face the future. Annu. Rev. Plant Biol. 2004, 55, 591–628. [Google Scholar] [CrossRef]
- Gifford, R.M. The CO2 fertilising effect—Does it occur in the real world? The International Free Air CO2 Enrichment (FACE) Workshop: Short- and long-term effects of elevated atmospheric CO2 on managed ecosystems, Ascona, Switzerland, March 2004. New Phytol. 2004, 163, 221–225. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Pinter, P.J.; Kimball, B.A.; Adamsen, F.J.; LaMorte, R.L.; Wall, G.W.; Hunsaker, D.J.; Adam, N.; Brooks, T.J.; Garcia, R.L.; et al. Leaf nitrogen concentration of wheat subjected to elevated [CO2] and either water or N deficits. Agric. Ecosyst. Environ. 2000, 79, 53–60. [Google Scholar] [CrossRef]
- Parry, M.L.; Rosenzweig, C.; Iglesias, A.; Livermore, M.; Fischer, G. Effects of climate change on global food production under SRES emissions and socio-economic scenarios. Global Environ. Change 2004, 14, 53–67. [Google Scholar] [CrossRef]
- Long, S.P.; Ainsworth, E.A.; Leakey, A.D.B.; Nosberger, J.; Ort, D.R. Food for thought: Lower-than-expected crop yield stimulation with rising CO2 concentrations. Science 2006, 312, 1918–1921. [Google Scholar] [CrossRef]
- Fischer, G.; Shah, M.; Tubiello, F.N.; van Velhuizen, H. Socio-economic and climate change impacts on agriculture: An integrated assessment, 1990–2080. Philos. Tran. R. Soc. B 2005, 360, 2067–2083. [Google Scholar] [CrossRef]
- Vorosmarty, C.J.; Green, P.; Salisbury, J.; Lammers, R.B. Global water resources: Vulnerability from climate change and population growth. Science 2000, 289, 284–288. [Google Scholar] [CrossRef]
- Brown, M.E.; Funk, C.C. Climate—Food security under climate change. Science 2008, 319, 580–581. [Google Scholar] [CrossRef]
- Fischer, G.; Tubiello, F.N.; van Velthuizen, H.; Wiberg, D.A. Climate change impacts on irrigation water requirements: Effects of mitigation, 1990–2080. Technol. Forecast. Soc. Change 2007, 74, 1083–1107. [Google Scholar] [CrossRef]
- Betts, R.A.; Falloon, P.D.; Gornall, J.; Kaye, N.; Wiltshire, A.; Wheeler, T. Climate Change and Food Security. In Climate Sense; Asrar, G.R., Ed.; Tudor Rose: Leicester, UK, 2009. [Google Scholar]
- IPCC, Climate Change 2007: Synthesis Report; Cambridge University Press: Cambridge, UK, 2007.
- Betts, R.A.; Arnell, N.W.; Boorman, P.M.; Cornell, S.E.; House, J.I.; Kaye, N.R.; McCarthy, M.P.; McNeall, D.J.; Sanderson, M.G.; Wiltshire, A.J. Climate change impacts and adaptation: An earth system view. In Understanding the Earth System; Cornell, S.E., Prentice, I.C., House, J.I., Downy, C.J., Eds.; Cambridge University Press: Cambridge, UK, 2012; p. 296. [Google Scholar]
- Schmittner, A.; Urban, N.M.; Shakun, J.D.; Mahowald, N.M.; Clark, P.U.; Bartlein, P.J.; Mix, A.C.; Rosell-Mele, A. Climate sensitivity estimated from temperature reconstructions of the last glacial maximum. Science 2011, 334, 1385–1388. [Google Scholar] [CrossRef] [Green Version]
- Sitch, S.; Cox, P.M.; Collins, W.J.; Huntingford, C. Indirect radiative forcing of climate change through ozone effects on the land-carbon sink. Nature 2007, 448, 791–794. [Google Scholar]
- Bond, T.C.; Doherty, S.J.; Fahey, D.W.; Forster, P.M.; Berntsen, T.; DeAngelo, B.J.; Flanner, M.G.; Ghan, S.; Kärcher, B.; Koch, D.; et al. Bounding the role of black carbon in the climate system: A scientific assessment. J. Geophys. Res. Atmos. 2013. [Google Scholar] [CrossRef]
- Shindell, D.; Kuylenstierna, J.C.I.; Vignati, E.; van Dingenen, R.; Amann, M.; Klimont, Z.; Anenberg, S.C.; Muller, N.; Janssens-Maenhout, G.; Raes, F.; et al. Simultaneously mitigating near-term climate change and improving human health and food security. Science 2012, 335, 183–189. [Google Scholar]
- Kopp, R.E.; Mauzerall, D.L. Assessing the climatic benefits of black carbon mitigation. Proc. Natl. Acad. Sci. USA 2010, 107, 11703–11708. [Google Scholar] [CrossRef]
- UNFCCC. Decision 2/CP.15 Copenhagen Accord, 2009. Available online: http://unfccc.int/resource/docs/2009/cop15/eng/11a01.pdf (accessed on 1 February 2012).
- Robock, A.; Marquardt, A.; Kravitz, B.; Stenchikov, G. Benefits, risks, and costs of stratospheric geoengineering. Geophys. Res. Lett. 2009. [Google Scholar] [CrossRef]
- Pongratz, J.; Lobell, D.B.; Cao, L.; Caldeira, K. Crop yields in a geoengineered climate. Nat. Clim. Change 2012, 2, 101–105. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wiltshire, A.J.; Kay, G.; Gornall, J.L.; Betts, R.A. The Impact of Climate, CO2 and Population on Regional Food and Water Resources in the 2050s. Sustainability 2013, 5, 2129-2151. https://doi.org/10.3390/su5052129
Wiltshire AJ, Kay G, Gornall JL, Betts RA. The Impact of Climate, CO2 and Population on Regional Food and Water Resources in the 2050s. Sustainability. 2013; 5(5):2129-2151. https://doi.org/10.3390/su5052129
Chicago/Turabian StyleWiltshire, Andrew J., Gillian Kay, Jemma L. Gornall, and Richard A. Betts. 2013. "The Impact of Climate, CO2 and Population on Regional Food and Water Resources in the 2050s" Sustainability 5, no. 5: 2129-2151. https://doi.org/10.3390/su5052129
APA StyleWiltshire, A. J., Kay, G., Gornall, J. L., & Betts, R. A. (2013). The Impact of Climate, CO2 and Population on Regional Food and Water Resources in the 2050s. Sustainability, 5(5), 2129-2151. https://doi.org/10.3390/su5052129