The Soil Microbial Community and Grain Micronutrient Concentration of Historical and Modern Hard Red Spring Wheat Cultivars Grown Organically and Conventionally in the Black Soil Zone of the Canadian Prairies

Abstract
: Micronutrient deficiencies in the diet of many people are common and wheat is a staple food crop, providing a carbohydrate and micronutrient source to a large percentage of the world's population. We conducted a field study to compare five Canadian red spring wheat cultivars (released over the last century) grown under organic and conventional management systems for yield, grain micronutrient concentration, and soil phospholipid fatty acid (PLFA) profile. The organic system had higher grain Zn, Fe, Mg and K levels, but lower Se and Cu levels. There was no trend in the results to suggest that modern western Canadian hard red spring cultivars have lower grain micronutrient content than historical cultivars. Wheat cultivar choice is important for maximizing grain nutrient levels, which was influenced by management system. It is evident that the emphasis on elevated grain quality in the western Canadian hard red spring class has resulted in the retention of micronutrient quality characters. Three fungal PLFAs were indicators for the organic system, and all three of these indicators were positively correlated with grain Cu concentration. In the organic system, percent arbuscular mycorrhizal fungi were negatively correlated with grain Zn and Fe concentrations, and positively correlated with grain Mn, Cu, K concentrations and grain yield. The organic system had higher levels of fungi in the soil, including arbuscular mycorrhizal fungi. Organic management practices appear to result in elevated levels of grain micronutrient concentration. The hard red spring breeding effort in and for the black soil zone of the northern Great Plains also appears to have led to no diminishment of grain micronutrient concentration. It is evident that both the agronomic system and breeding strategies in this region can be exploited for future increases in grain micronutrient concentration.1. Introduction
Consumers purchase organic food products because they perceive these foods to have unique and/or superior quality attributes relative to conventional foods [1]. Organic food products are viewed as more healthy, safe and/or environmentally friendly than conventional food products [1]. Consumers list a number of benefits of organic products as important in their choice to buy organic from most important to least important: health, environment, taste, animal welfare, minimal processing, novelty and fashion [2]. Despite the perceived benefits of consuming organic foods, organic certification is based on the process used to produce the good, not on the product itself; meaning that organic certification is not a guarantee of the quality attributes of the product [3]. Many of the perceived attributes of organic products cannot be measured, and necessitate faith on the part of the consumer that the desired attributes are present [4].
Micronutrients play important roles in human body functioning and health. Deficiencies are most generally present in developing countries, often affecting women and children [5]. However, at least half the US population fails to meet the recommended dietary allowance for the micronutrients magnesium (Mg), calcium (Ca), and zinc (Zn) [6]. The essential trace elements Zn, copper (Cu) and Se are antioxidant micronutrients. They play a key role in the body's defenses against free radicals and reactive oxygen molecules as constituents of antioxidant exzymes. Increasing mineral nutrient content of crops, or biofortification, can help prevent nutrient deficiencies. Biofortification may be achieved through agronomic measures to increase the availability of mineral elements in the soil, as well as the development and use of crops with increased ability to absorb nutrients [7]. Globally, wheat is a major source of nourishment and the main source of carbohydrates in temperate areas [8]. There has been speculation that the development of high yielding cultivars in association with input intensification has resulted in lower levels of micronutrients in grains produced in conventional environments. In a meta-analysis study, Worthington [9] reported that organic crops contained significantly more Fe and Mg than conventional crops. There were nonsignificant trends showing a higher content of nutritionally significant minerals in organic crops. Murphy et al. [10] reported, in a Pacific Northwest USA study of 63 historical and modern wheat cultivars that, while grain yield has increased over time, the concentrations of all minerals except Ca have decreased. They further reported that the decrease in mineral concentration over the past 120 years occurred mainly in the soft white wheat and not the hard red market class in the western USA.
Soils, climate, crop type and cultivar, management practices and post-harvest factors can all affect the nutritional quality of crops [11]. Research into nutritional differences between organic and conventional crops has yielded conflicting results or small differences [12]. Mäder et al. [13] reported few differences in nutrient content of organic and conventional wheat grain in a long-term field study. Ryan et al. [14] found that the nutritional value of organic grain was slightly better than conventional grain, with higher Zn and Cu concentrations in organic wheat. However, the authors concluded that universal statements about differences between organic and conventional management were not useful; but that knowing the underlying mechanisms leading to nutritional quality differences could lead to improved nutritional quality. In another long-term field study, Turmel et al. [15] reported that differences in grain mineral content of organic and conventional wheat are dependent on the system's crop rotation (annual grain vs. perennial forage-grain rotation).
Soil microbes affect soil fertility in many ways, for instance through their influence on organic matter turnover, mineral immobilization and dissolution, and soil aggregation [16]. As well, soil microbes such as Rhizobia bacteria and arbuscular mycorrhizal fungi form plant symbioses, improving plant nutrient supply or uptake. While management practices can alter microbial communities directly, for the most part, management practices change microbial communities indirectly by altering soil properties affecting microbial diversity. Because most microbes are heterotrophic, soil organic matter content and the type and amount of organic materials added to the soil are two critical soil factors affecting microbial diversity [17]. Genetic differences within a crop species (i.e., cultivar differences) may play a role in the structure of the microbial community. Differences have been found in microbial communities associated with different wheat and canola (Brassica napus L.) cultivars [18].
Both organic and conventional cropping systems involve the combination of a number of different management practices (chemical use, tillage, crop rotation and crop choice). Despite the ranges of management practices used in organic and conventional cropping systems, there are a number of factors which commonly differ between the two systems. On the western Canadian prairies, according to research conducted over the last 10 years, organic systems tend to have higher organic matter, more weeds, 10 to 50% lower grain yields and lower phosphorus (P) levels than conventional systems [19,20]. The absence of inorganic fertilizers and pesticides in organic systems generally leads to greater weed populations, higher tillage intensities, and lower soil nutrient levels. Thus, at least in extensive grain producing systems of the northern Great Plains, organic systems generally tend to present greater biotic and abotic stresses to production systems than their conventional counterparts. This is, in part, because these stresses cannot be managed through chemical pesticides and fertilizer [20].
Organic systems often have more diverse crop rotations and higher plant diversity within fields than conventional systems. Organic systems employ a number of practices that serve to increase organic matter content, generally resulting in slightly higher levels of organic matter in organic systems versus conventional systems [21,22]. Some studies have reported shifts in microbial communities with organic versus conventional management [21,23], while others have reported no differences between microbial communities under the two management systems [24,25].
We hypothesized that the stress levels associated with organic management systems would result in higher levels of micronutrients in the grain, and specifically those with antioxidant properties. We further hypothesized that western Canadian historic hard red spring cultivars would have higher mineral content than modern cultivars, especially in organic systems; and that those organic systems would have a more diverse microbial community structure. Thus, the objectives of this study were to compare the concentrations of seven minerals, including Se, Cu, Mn, Zn, Fe, Mg and K in a subset of hard spring wheat cultivars grown over the last 100 years in western Canada in both conventional and organic environments. We were also interested in exploring the effect of the microbial community structure in the two management systems on micronutrient content.
2. Materials and Methods
We reported the effect of cultivar and management system (organic vs. conventional) on agronomic traits, soil biotic data and breadmaking quality traits using the present experimental design [26]. The present study is thus a continuation of a reported study where the experimental design is similar in many respects. To summarize, we grew five western Canadian spring wheat cultivars (AC Elsa, Glenlea, Marquis, Park and AC Superb) (see descriptions in McCallum and DePauw (2008)) in four-replicate, randomized complete block experiments: one organically managed site and one conventionally managed site in 2005, 2006 and 2007 in Edmonton, AB (55°34′ N, 113°31′ W). The cultivars were chosen to represent spring wheat cultivars commonly grown in western Canada over the past century (Table 1). The two sites were located within 500 m of one another with the same soil type (Orthic Black Chernozem) and topography. Plot dimensions were 4 m by 1.38 m, with 6 rows spaced 23 cm apart, seeded with a no-till double disk drill (Fabro Enterprises Ltd., Swift Current, SK, Canada) in May. The 5 ha organic site has been in a long term, three-year rotation of experimental spring-sown grain, followed by a fall-sown rye (S. cereal L.) taken for silage the following year, followed by a triticale (X Triticosecale Wittmack)-pea (P. sativum L.) mixture again harvested for silage. The entire organic site has been managed organically since 1999. The conventional site is in a long term four-year rotation in the following order: (1) experimental canola (B. napus L.) plots, (2) triticale-pea mixtures (or uniform pea) harvested as silage, (3) experimental spring wheat plots, and (4) barley (H. vulgare L.) silage.
Fertilizer application followed soil tests and local recommendations for the conventional site (Table 2). The soil nutrient analyses were conducted at Exova Labs (formerly Bodycote) in Edmonton, AB Canada. The conventional land had 112 kg N ha−1 banded in the form of urea (46-0-0) in the fall. Thirty-six kg ha−1 of 11-52-0 was banded with the seed for 4 kg ha−1 of N and 19 kg ha−1 of P2O5. The organic land had compost (comprised of dairy manure, sawdust, wood chips and straw) applied each year for the 5 years prior to 2005 at a rate of 50–62 t ha−1 (fresh weight). No compost was added during the trial years of 2005 to 2007. Dyvel ® (active ingredients Dicamba [present as dimethylamine salt] and MCPA [present as potassium salt]) (BASF Canada Inc., Mississauga, ON) was applied at 1.2 L of product per hectare (100.8 g ha−1 of Dicamba and 403.2 g ha−1 of MCPA) at the recommended crop and weed stage in the conventional site only. Weeds were controlled in the fall (cultivator only) and spring with tillage (cultivator and harrows), with an additional final pass with a cultivator and harrows directly prior to seeding on the organic site. A 5-m wide Morris field cultivator and 10-m wide diamond harrows were used for these operations.
Soil for mycorrhyzal samples was taken approximately 60 days after seeding. Soil cores (0–15 cm) were taken directly below five randomly chosen wheat plants from each plot and placed in plastic bags in storage at −20 °C until subsampling. Two grams of soil was subsampled from each soil core at the origin of root growth where roots were most dense (to collect rhizosphere soil) and freeze-dried. We harvested the wheat plots using a Wintersteiger plot combine Model 1991 Nurserymaster Elite (Ried, Austria) in late September and early October, depending on crop maturity. Harvested grain was dried at 40 °C for approximately 48 hours. Grain yield was determined and is presented after drying to 14% moisture. Weed seeds were removed using sieves and a grain blower. Sieve size used for cleaning grain was a 2-mm mesh sieve (Canadian Standard Sieve Series no.10). Small weed seeds were also removed using a fan.
Composition of the soil microbial community was determined using the phospholipid fatty acid analysis (PLFA) method as described by Dalpé and Hamel [27]. In brief, fatty acids were extracted using a dichloromethane : methanol : citrate buffer. Phospholipid fatty acids were separated from other lipids on solid phase extraction columns made from flamed glass pipettes and silica gel. Samples were methylated using a mild acid methanolysis. Samples were analyzed using a Hewlett Packard 5890 II Plus Gas Chromatograph, with MIDI Inc. MIS (Microbial Identification System) Version 4 used for peak identification. Weight of individual PLFAs in the sample were determined by comparing the area under a specific peak with the 19:0 peak value, which was determined using a standard curve of 19:0 Fatty acid methyl ester (FAME) standard dissolved in hexane. PLFA markers from DeGrood et al. [28] were used to calculate the percent of total biomass as gram negative bacteria, gram positive bacteria, and actinomycetes. The PLFA marker 16:1 w5 was used to indicate mycorrhizal fungi [27]. Richness was determined by counting the number of unique PLFAs in a sample. Shannon's diversity index was calculated for each soil sample using the formula:
Micronutrient traits were determined on the whole grain at the Crop Development Centre, Saskatoon, SK. A subsample of 100 g of dry seed was taken from each plot, ground and prepared following a standard HNO3 H2O2 digestion methods [29,30]. CDC Redberry lentil seeds and organic wheat were used as laboratory reference materials and measured periodically to ensure consistency in the method. Total micronutrient (Mg, K, Mn and Cu) concentrations in lentil seeds were measured using flame-AAS (AJ ANOVA 300, Lab Synergy, Goshen, NY, USA). Total Se concentrations in lentil seeds before and after cooking were measured using hydride generation (HG)-AAS (AJ ANOVA 300, Lab Synergy, Goshen, NY, USA).
The organic and conventional sites were less than 500 m apart, with similar growing temperatures and precipitation levels within years. Temperatures were within 3 °C of the 30-year average for the three field seasons. Precipitation averaged 206 mm from May-September over the three years, with the 30-year average being 341 mm. Differences between the two systems are likely due to the different management strategies and the resulting differences in nutrient levels (Table 2) and weed pressure. Analyses of data were performed using the PROC MIXED procedure of SAS (version 9.1, SAS Institute). The experiment was analysed as a split plot with management system as the main plot and cultivar as the sub plot. Using such an analysis implies that one complete block was grown per year. Thus, years were treated as blocks and were considered as a random effect in both combined analysis and in the analyses by management system. Blocks within site were considered as random sampling units. Cultivar and management system (where applicable) were considered as fixed effects. All interactions with random effects were also considered random. We also analyzed data by management system, combined over years. For both analyses, years and blocks were considered random and management system and cultivar were considered fixed effects. Pearson correlations were conducted on plot means for the organic and conventional sites. The split-plot design has a decreased precision for main plot (management) comparisons [31], which was of interest to us. Preliminary analysis revealed no significant differences in microbial community among the wheat cultivars in the organic system. We discuss and report differences between treatments only when P ≤ 0.05, except for the mixed model analysis of the experiment, where P ≤ 0.10 was used as a significance level appropriate for the small experimental size [31].
The structure of the microbial community (PLFA) for the entire experiment, and for the two management systems separately was characterized with non-metric multidimensional scaling (NMS) ordination using PC-ORD (version 5, MjM Software Design). Statistical analyses were conducted on the mean laboratory results of the five sub-samples per plot. Fatty acids present in less than 10% of the samples were omitted from the analysis. A total of 43, 47 and 40 unique PLFAs were used in the combined management system, organic and conventional ordinations, respectively. The PLFA data were arcsine square-root transformed prior to ordination. A Sorensen (Bray-Curtis) distance measurement was used in all ordinations. Biplots were used to show the relationship between grain micronutrient concentration, yield and PLFA ordination scores. Significant differences between management system, year and wheat cultivars were evaluated on the NMS analyses using a multi-response permutation procedure (MRPP). Indicator species analyses were used to evaluate the relationship between individual PLFAs and the two management systems.
The experiments were conducted on a single organic/conventional site comparison and differences between the management systems are therefore specific to the site soil and growing conditions. These results should be interpreted in conjunction with further data compiled in this region (e.g., [20]).
3. Results
Yields were significantly lower in the organic system than the conventional system (2.74 vs. 5.02 t ha−1, respectively) (Table 3). AC Superb (the most modern and semi-dwarf cultivar) yielded the most grain, and Marquis (the oldest cultivar) the least in both systems. Flour yield was significantly higher in the conventional system than the organic system. AC Elsa (a modern tall cultivar) had the highest flour yields in both the organic and conventional systems. Full agronomic and baking quality data and analyses for this study have been reported elsewhere [26].
3.1. Grain Micronutrient Concentration
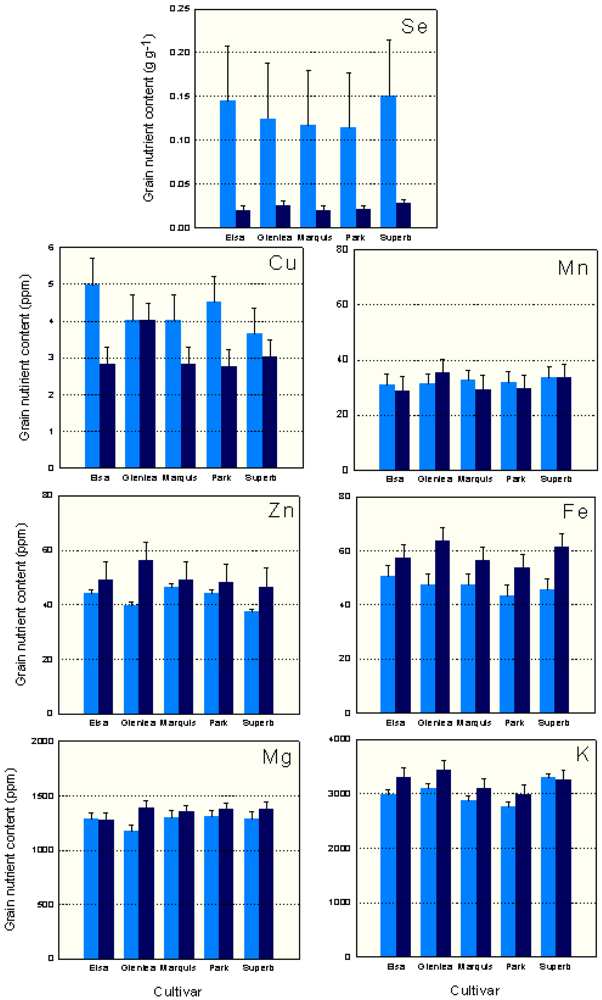
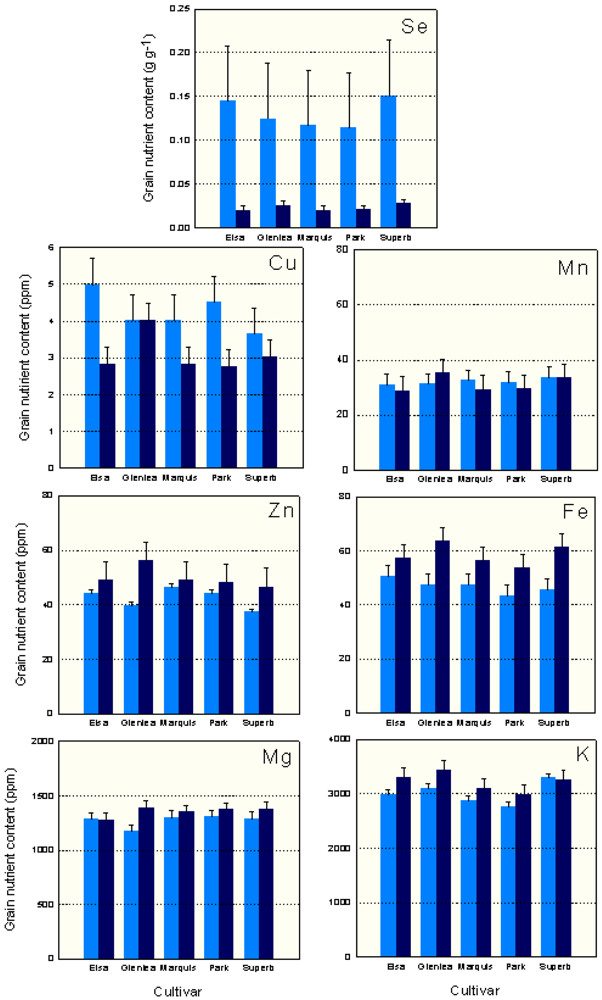
In the organic system, Glenlea had the highest levels of Zn, Mn, Mg, K and Fe, followed by the most recent cultivars AC Elsa or AC Superb (Figure 1). In the conventional system, the historical cultivar Marquis had the highest levels of Zn, and Park (1963 release) had the highest Mg levels; the modern cultivars had the highest K (AC Superb) and Fe (AC Elsa) grain concentrations. Under conventional management, Glenlea had intermediate to low levels of grain micronutrients (Figure 1).
There were significant management × cultivar interactions for Cu, Mn, Zn, Fe, Mg and K (Table 3). AC Elsa grown conventionally had higher levels of grain Cu than AC Elsa, Marquis and Park grown organically. Organically grown Glenlea had higher grain Mn, Zn, Fe, Mg and K concentrations than other organically and conventionally grown cultivars. In the organic system, Glenlea grain had higher Mn, Fe and K levels than Park, higher Mn and Mg levels than AC Elsa and higher Zn concentration than AC Superb. Organically grown Glenlea grain also had higher Fe than conventional Park and AC Superb, and higher K concentration than conventional Park and Marquis grain.
On average, the conventional system had higher (P < 0.05) grain Se and Cu concentration, while the organic system had higher grain Zn, Fe, Mg and K levels. In the organic system, grain yield and flour yield were negatively correlated with grain Zn, Fe and Mg concentrations (Table 4). In the conventional system, only grain Mg concentration was negatively correlated with both grain and flour yield. Grain protein content was positively correlated with Zn in both systems, and Fe in the organic system. In both systems, grain Fe and Zn levels were positively correlated. In the organic system, grain Mg concentration was correlated with Zn and Fe. As well, grain K concentration was correlated with Mn and Cu concentrations. In the conventional system, grain Mn was positively correlated with grain Fe and K concentrations. Negative correlations were detected between conventional grain Cu and Mn, and Mg and Fe concentrations (Table 4).
3.2. Soil Microbial Community
The non-metric multidimensional scaling (MRPP) analysis revealed that the soil microbial community grouped differently under organic versus conventional management (P < 0.001) and between years; with the management effect having a stronger effect than years (data not shown). Cultivars exhibited no significant grouping patterns. There were significant grouping patterns in the management × cultivar interaction. Nevertheless, the individual comparisons suggest that all cultivars grown under the organic management system group differently than all cultivars grown under conventional management.
Three fungal PLFAs (16:1 w5c, 18:1 w9c and 20:1 w9c) were indicators for the organic system, with the fungal PLFA 20:1 w9c strongly associated with the organic system. Other strong indicator PLFAs for the organic system were the Gram+ bacterial PLFAs i14:0, i18:0 and i19:0 (Table 5).
In the organic system % AM fungi was negatively correlated with grain Zn and Fe concentrations and positively correlated with grain Mn, Cu, K concentrations, as well as grain yield (Table 6). PLFA diversity was positively correlated with grain Zn and Fe concentrations and seed yield, but negatively correlated with grain concentrations of Se, Mn, and Cu, and grain yield. Some indicator species for the organic system had weak positive correlations with Mn, Cu and K grain concentrations. The three fungal PLFAs were moderately correlated with grain Cu concentration.
In the conventional system % AM fungi was negatively correlated with grain concentrations of Zn and Fe, as well as grain protein content. Percent gram positive bacteria and actinomycetes had moderate negative correlations with grain Se and Cu concentrations, as well as grain and flour yield (Table 7). These biomarkers had moderate positive correlations with grain Mn, Mg and K concentrations. PLFA diversity was positively correlated with grain Mn, Mg and K concentrations, but had moderate negative correlations with grain Se and Cu concentrations, along with grain and flour yields.
4. Discussion
We originally hypothesized that the stress levels associated with organic management systems would result in higher levels of micronutrients in the grain, and specifically those with antioxidant properties. The organic system in the present study had higher grain Zn, Fe, Mg and K, but lower grain Se and Cu. Thus, only one (Zn) of the three antioxidants studied (Zn, Se and Cu) was indeed present in the grain in greater concentrations in the organic system studied here. Selenium was present in very low concentrations in the wheat grain in this study when compared with other studies of similar ilk [13,32]. The Se concentration of wheat cultivars has been reported to be more influenced by soil supply than genotype [32]. Some studies have reported some differences in wheat nutrient content grown organically versus conventionally [13-15]. However, the choice of management practices (e.g., tillage practices, crop rotation, fertility amendments) used within both of the systems under the specific local conditions will have an effect on soil fertility, and the subsequent grain nutrient concentration. Farmyard manure has been reported to provide a sustained supply of Cu and Zn to cropping systems, increasing yields and improving wheat Cu and Zn uptake over fallow for at least three years following manure application [33]. Ryan et al. [14] reported that organic systems increased wheat grain Cu, N and Zn and decreased grain Fe and Mn, but concluded that differences in mineral contents were due to specific management practices chosen by producers, not from the overall management system. Turmel et al. [15] concluded that differences in mineral content of wheat grown organically and conventionally was dependent on crop rotation. Decreased grain yield, and possibly smaller grain size, may, in part, account for increased grain nutrient concentration in the present organic system. We further hypothesized that historic western Canadian hard red spring cultivars would have higher mineral content than modern cultivars, especially in organic systems; and that those organic systems would have a more diverse microbial community structure. Significant interaction effects of management × cultivar indicate that the choice of wheat cultivar to maximize grain micronutrient level is dependent on management system. Glenlea grown organically had the highest grain nutrient levels compared to cultivars grown either organically and conventionally. Glenlea was the only Canadian extra strong wheat grown in the trial. Glenlea is generally characterized as having medium protein content and stronger gluten than the other cultivars studied [34]. Recently, it was reported that the introduction of semi-dwarf high-yielding cultivars has coincided with a decrease in wheat grain Cu, Zn, Fe and Mg contents [35]. However, our results suggest no trend to lower grain mineral concentrations in the modern, high-yielding red spring wheat cultivars in Western Canada. This result is similar to that of Murphy et al. [10] who reported a neutral trend between year of release and grain mineral content of hard red spring wheat cultivars of the Pacific Northwest. Murphy et al. [10] did report a decline in all minerals tested (except Ca) in soft white wheat, where yield has been successfully increased through breeding (apparently at the expense of some quality characters). Canadian hard spring bread wheat breeding programs have emphasized grain quality and disease resistance over the last century. It is evident that this emphasis on elevated grain quality in the western Canadian hard red spring class has resulted in retention of micronutrient quality characters and not an erosion of this quality characteristic.
Negative relationships between grain yield and grain Se, Zn, Fe, Mg and P have been reported in bread wheat [7]. In the organic system, grain Zn, Fe, Mn and Mg concentrations were negatively correlated with grain yield. However, in the conventional system only grain Zn and Mg concentrations had weak and moderate (respectively) negative associations with yield. In both the organic and conventional systems there was a positive relationship between grain Zn and Fe concentrations. Rawat et al. [36] found wheat accessions with high Fe content also had high Zn content, leading the authors to hypothesize that the nutrients shared similar uptake, translocation and deposition mechanisms.
In the organic system of the present study, the presence of AM fungi appears to have improved the uptake of Mn and Cu, but had a detrimental effect on Zn uptake. The indicator species for the organic system were positively correlated with grain Mn and Cu concentrations. Specifically, the PLFA biomarkers for fungi were all moderately correlated to grain Cu concentration, indicating that soil fungi played a role in plant Cu fertility.
In both the organic and conventional systems, % AM fungi were negatively correlated with grain Zn and Fe concentrations. In the organic system, % AM fungi and weight of AM fungi were positively correlated with Mn and Cu concentrations of wheat grain, as well as grain yield. Mycorrhizae can increase plant uptake of a number of nutrients, including: N, P, K, Ca, Mg, S, Fe, Mn, Cu and Zn [14,37-40], however, where mycorrhizae are most beneficial is in the uptake of relatively immobile nutrients such as phosphorus, copper and zinc [41].
Phospholipid fatty acid diversity was positively correlated with grain nutrient concentration of some nutrients in both the organic and conventional systems, but differing nutrients in the two systems. Diversity was also negatively correlated with some grain mineral concentrations. In both systems, diversity was negatively correlated with grain Se and Cu concentrations, as well as grain yield.
The soil microbial community structure was different under the two management systems. The management practices that differed between the organic and conventional systems (crop rotation, along with fertility, pest management and tillage regimes) can all affect soil microbes, so we expected to see differences between the community structures of the two systems. Differences between organic and conventional management systems have been previously reported [21,23]. Differences are generally expected between microbial communities of organic and conventional cropping systems, and instances of no significant differences have been attributed to overriding effects of soil type and/or time of sampling [21,24,25].
All but one of the indicator species of PLFAs were indicators for the organic system. The three fungal PLFA biomarkers found in the soil were biomarkers for the organic cropping system, including biomarker 16:1 w5c, which is used as a biomarker for AM fungi. Mycorrhizal potential, and actual colonization has been reported to be greater in grasslands, organic and low-input systems versus conventional systems [42-44]. This is largely attributed to the fact that the application of phosphorus fertilizers, even at low rates, decreases root colonization of mycorrhizal fungi [14,42]
5. Conclusions
The organic system in the present study had higher grain Zn, Fe, Mg and K, but lower grain Se and Cu. Wheat cultivar choice appears to be important for maximizing grain nutrient levels, and varies depending on the management system. It is evident that the emphasis on elevated grain quality in the western Canadian hard red spring class has resulted in the retention of micronutrient quality characters. The organic system had higher levels of fungi in the soil, including AM fungi, and these fungi biomarkers appeared to play a role in grain Cu concentration.
With elevated levels of micronutrients seemingly a component of both organic management practices and the general hard red spring breeding effort in western Canada, it is evident that both the agronomic system and breeding efforts in this region may lead to future increases.
Organic and conventional management systems have different soil microbial communities, and these communities can play a role in soil fertility and final grain nutritional content. This study focused on the effect of Canadian hard spring wheat cultivar choice on soil microbial communities and crop nutritional quality within organic and conventional systems in the black soil zone of the northern Great Plains. Further studies are required to determine the impact of other agronomic cropping practices such as tillage, fertility management and a larger subset of wheat cultivars to identify best management practices within organic and conventional systems for soil fertility and final crop quality.
It has been mentioned throughout the literature that increasing micronutrient levels in grain is but a small component of improving health outcomes for the many people in both the developed and developing world suffering nutrient deficiencies. Nevertheless, the results of the present study suggest that nutritional advances in the micronutrient content of wheat can be made through the choice of management system and through plant breeding. Such advances do not necessarily have to come at the expense of elevated or stable grain yield.

{kind=link}
| Year of registration | Wheat classz | Maturityy | Height (cm)x | Parentage | |
|---|---|---|---|---|---|
| Marquis | 1910 | CWRS | 2 | 113 | Red Fife × Hard Red Calcutta |
| Park | 1963 | CWRS | −2 | 91 | (Mida × Cadet) × Thatcher |
| Glenlea | 1972 | CWES | 0 | - | (Pembina × Bage) × CB100 |
| AC Elsa | 1996 | CWRS | 0 | 89 | BW90 × Laura |
| AC Superb | 2000 | CWRS | 1 | 85 | Grandin*2 × AC Domain |
zCWRS: Canadian Western Hard Red Spring Wheat, CWES: Canadian Western Extra Strong Wheat;yCompared to AC Barrie: 106 days to maturity. Source: [26];xHeight information taken from cultivar release information [26].
| Year | Organic | Conventional | ||||
|---|---|---|---|---|---|---|
| 2005 | 2006 | 2007 | 2005 | 2006 | 2007 | |
| NO3 (kg ha−1) | 199 | 227 | 56 | 272 | 252 | 41 |
| P (kg ha−1) | 260 | 262 | 83 | 192 | 213 | >134 |
| K (kg ha−1) | 1,582 | 1,581 | 533 | 1,462 | 1,352 | 965 |
| SO4 (kg ha−1) | >90 | >85 | 16 | >90 | 60 | 21 |
| Cu (ppm) | - | 0.83 | 1.23 | - | 1.7 | 0.69 |
| Mn (ppm) | - | 13.8 | 14 | - | 12.9 | 10.9 |
| Zn (ppm) | - | 8.42 | 6.61 | - | 8.17 | 7.23 |
| Fe (ppm) | - | 125 | 141 | - | 115 | 107 |
| Mg (ppm) | - | 574 | 2,060 | - | 625 | 633 |
| pH | 6.1 | 6.3 | 6.4 | 7.3 | 5.9 | 6.6 |
| Electrical conductivity (dS m−1) | 0.91 | 0.83 | 0.5 | 0.99 | 0.69 | 0.44 |
| Organic matter % | 10.3 | 12.9 | 14.6 | 7.2 | 12.8 | 12.5 |
| Cultivar | Yield | Flour yield | Se | Cu | Mn | Zn | Fe | Mg | K |
|---|---|---|---|---|---|---|---|---|---|
| (YOR††) | (t ha−1) | (%) | (μg g−1) | (ppm) | (ppm) | (ppm) | (ppm) | (ppm) | (ppm) |
| Organic | |||||||||
| Marquis (1910) | 2.38 | 69 | 0.020 | 2.83 | 29.4 | 49.2 | 56.5 | 1352 | 3110 |
| Park (1963) | 2.65 | 71 | 0.021 | 2.75 | 29.4 | 48.0 | 53.8 | 1375 | 2989 |
| Glenlea (1972) | 2.78 | 69 | 0.025 | 4.02 | 35.4 | 56.0 | 63.5 | 1390 | 3443 |
| AC Elsa (1996) | 2.80 | 72 | 0.020 | 2.83 | 29.0 | 49.1 | 57.3 | 1278 | 3296 |
| AC Superb (2000) | 3.11 | 70 | 0.028 | 3.02 | 33.7 | 46.6 | 61.3 | 1376 | 3259 |
| F testcultivar | *** | *** | * | *** | *** | * | *** | ** | *** |
| SEcultivar | 0.796 | 1.0 | 0.0050 | 0.469 | 4.98 | 6.73 | 5.02 | 62.2 | 173.6 |
| Conventional | |||||||||
| Marquis (1910) | 4.04 | 71 | 0.117 | 4.00 | 32.6 | 46.3 | 47.48 | 1299 | 2886 |
| Park (1963) | 4.66 | 72 | 0.115 | 4.50 | 31.9 | 44.0 | 43.39 | 1306 | 2770 |
| Glenlea (1972) | 5.57 | 73 | 0.125 | 4.02 | 31.3 | 39.9 | 47.42 | 1171 | 3101 |
| AC Elsa (1996) | 5.09 | 74 | 0.145 | 4.99 | 31.1 | 44.2 | 50.72 | 1285 | 2995 |
| AC Superb (2000) | 5.73 | 74 | 0.151 | 3.64 | 33.6 | 37.2 | 45.73 | 1289 | 3296 |
| F testcultivar | *** | *** | ns | ns | ns | *** | ** | *** | *** |
| SEcultivar | 0.421 | 0.6 | 0.0627 | 0.709 | 3.81 | 1.26 | 4.00 | 61.5 | 78.8 |
| Organic | 2.74 | 70 | 0.022 | 3.07 | 31.3 | 49.6 | 58.3 | 1353 | 3218 |
| Conventional | 5.00 | 73 | 0.131 | 4.22 | 32.1 | 42.3 | 46.9 | 1270 | 3009 |
| F testmanagement | *** | *** | *** | *** | ns | *** | *** | *** | *** |
| SEmanagement | 0.401 | 0.6 | 0.0336 | 0.492 | 3.89 | 3.43 | 3.51 | 35.1 | 90.9 |
| F testcultivar | *** | * | ns | ns | *** | *** | *** | ** | *** |
| SEcultivar | 0.467 | 0.8 | 0.0359 | 0.523 | 3.93 | 3.63 | 3.63 | 44.2 | 97.1 |
| F testm*c | ** | ns | ns | ** | ** | *** | *** | *** | *** |
†ns = not significant (P ≥ 0.10);*significant at P < 0.10;**significant at P < 0.05;***significant at P < 0.01; SE = standard error;††Year of release.
| Se | Zn | Fe | Mn | Cu | Mg | K | Yield | Grain protein | Flour yield | |
|---|---|---|---|---|---|---|---|---|---|---|
| Organic Management | ||||||||||
| Se | - | - | 0.40 | - | - | - | - | - | - | |
| Zn | −0.30 | 0.72 | 0.28 | - | 0.74 | - | −0.76 | 0.65 | −0.63 | |
| Fe | - | 0.48 | 0.28 | - | 0.59 | - | −0.63 | 0.50 | −0.45 | |
| Mn | - | 0.31 | 0.50 | 0.27 | 0.41 | 0.63 | −0.41 | - | - | |
| Cu | - | - | - | −0.48 | - | 0.49 | 0.33 | - | - | |
| Mg | −0.29 | - | −0.45 | - | - | - | −0.68 | 0.52 | −0.66 | |
| K | −0.27 | - | - | 0.48 | −0.32 | - | - | - | - | |
| Yield | 0.37 | −0.27 | 0.39 | - | - | −0.51 | 0.47 | −0.54 | 0.64 | |
| Grain protein | - | 0.52 | - | - | 0.30 | 0.36 | - | −0.30 | −0.35 | |
| Flour yield | 0.42 | - | 0.28 | - | - | −0.39 | 0.42 | 0.70 | - | |
| Conventional Management | ||||||||||
†All correlation coefficients shown are significant at P < 0.05;-indicates non-significance.
| PLFA | Origin | Mean | Indicator value | Monte Carlo | |
|---|---|---|---|---|---|
| Conv. | Organic | p < 0.05 | |||
| i14:0 | Gram+ | 35.4 (3.77) | 6 | 74 | 0.0002 |
| 16:1 w5c | Fungi | 50.8 (0.94) | 45 | 55 | 0.0002 |
| i17:0 | Gram+ | 50.4 (0.78) | 47 | 53 | 0.0002 |
| a17:0 | Gram+ | 50.3 (0.74) | 48 | 52 | 0.0002 |
| cyc17:0 | Bacteria | 50.4 (0.77) | 53 | 47 | 0.0002 |
| i18:0 | Gram+ | 19.8 (3.82) | 0 | 57 | 0.0002 |
| 18:1 w9c | Fungi | 50.4 (0.77) | 47 | 53 | 0.0002 |
| i19:0 | Gram+ | 27.0 (4.06) | 6 | 48 | 0.0006 |
| 20:1 w9c | Fungi | 44.8 (3.35) | 0 | 45 | 0.0002 |
| Se | Zn | Fe | Mn | Cu | Mg | K | Grain yield | Flour yield | |
|---|---|---|---|---|---|---|---|---|---|
| Gram– bacteria | −0.32 | 0.33 | - | −0.45 | - | - | - | −0.35 | - |
| Gram+ bacteria | −0.43 | - | - | - | - | - | - | - | - |
| Actinomycetes | −0.34 | 0.60 | 0.65 | −0.55 | −0.42 | 0.40 | −0.54 | −0.76 | −0.43 |
| AM Fungi | - | −0.44 | −0.47 | 0.44 | 0.41 | - | 0.46 | 0.63 | 0.38 |
| Richness | - | 0.33 | 0.38 | - | −0.48 | - | - | −0.59 | −0.47 |
| Evenness | −0.51 | 0.40 | 0.40 | −0.45 | - | - | - | −0.47 | - |
| Diversity | −0.47 | 0.46 | 0.47 | −0.34 | −0.35 | - | - | −0.62 | −0.45 |
| i14:0 (Gram+)‡ | - | - | - | 0.35 | - | - | - | 0.34 | - |
| 16:2 w5c (Fungi) | - | - | - | 0.35 | 0.41 | - | - | 0.35 | - |
| i17:0 (Gram+) | - | - | - | 0.35 | 0.41 | - | - | 0.32 | - |
| a17:0 (Gram+) | - | - | - | 0.35 | 0.47 | - | - | 0.33 | - |
| i18:0 (Gram+) | - | - | - | - | - | - | - | - | - |
| 18:1 w9c (Fungi) | - | - | - | - | 0.43 | - | - | - | - |
| i19:0 (Gram+) | - | - | - | - | 0.46 | - | 0.36 | - | - |
| 20:1 w9c (Fungi) | - | - | - | - | 0.66 | - | - | - | - |
†All correlation coefficients shown are significant at P < 0.05;-indicates non-significance;‡Brackets following specific indicator PLFA gives the microbial group associated with the indicator PLFA.
| Se | Zn | Fe | Mn | Cu | Mg | K | Grain yield | Grain protein | Flour yield | |
|---|---|---|---|---|---|---|---|---|---|---|
| Gram− bacteria | - | −0.37 | - | - | - | - | 0.32 | - | - | −0.41 |
| Gram− bacteria | −0.46 | - | - | 0.62 | −0.37 | 0.53 | 0.40 | −0.39 | - | −0.59 |
| Actinomycetes | −0.45 | - | −0.31 | 0.50 | -0.41 | 0.46 | 0.34 | −0.45 | - | −0.56 |
| AM Fungi | - | −0.65 | −0.34 | - | - | - | - | - | -0.46 | - |
| Richness | −0.52 | - | - | 0.67 | −0.40 | 0.55 | 0.42 | −0.42 | - | −0.58 |
| Evenness | −0.43 | - | - | 0.58 | −0.36 | 0.47 | 0.46 | −0.34 | - | −0.48 |
| Diversity | −0.48 | - | - | 0.63 | −0.40 | 0.53 | 0.46 | −0.40 | - | −0.56 |
| cyc17:0 (Bacteria)‡ | - | - | - | - | - | - | - | - | - | - |
†All correlation coefficients shown are significant at P < 0.05;-indicates non-significance;‡Brackets following specific indicator PLFA gives the microbial group associated with the indicator PLFA.
Acknowledgments
We thank Alireza Navabi, Klaus Strenzke, Derrick Kanashiro, Brian Bowen, Holley Yorston, Frances Reilly, Carla Ollenberger, Alana Kornelsen, Corrina Murdoch, and Madison Derworiz for technical support. We gratefully acknowledge the Natural Sciences and Engineering Research Council of Canada (NSERC), Alberta Ingenuity, and the University of Alberta for scholarship support to A. Nelson. We further gratefully acknowledge financial support to D. Spaner through a Discovery Grant from NSERC and a research grant from the Alberta Crop Industry Development Fund.
References
- Yiridoe, E.K.; Bonti-Ankomah, S.; Martin, R.C. Comparison of consumer perceptions and preference toward organic versus conventionally produced foods: A review and update of the literature. Renew. Agr. Food Syst. 2005, 20, 193–205. [Google Scholar]
- Lockie, S.; Haplin, D.; Gordon, R.; Pearson, D. Understanding the market for organic food. In Organic Agriculture: A Global Perspective; Kristiansen, P., Taji, A., Reganold, J., Eds.; Comstock Publishing Associates: Ithaca, NY, USA, 2006; pp. 245–258. [Google Scholar]
- Brandt, K.; Mølgaard, J.P. Food quality. In Organic Agriculture: A Global Perspective; Kristiansen, P., Taji, A., Reganold, J., Eds.; Comstock Publishing Associates: Ithaca, NY, USA, 2007; pp. 305–327. [Google Scholar]
- Ritson, C.; Oughton, E. Food consumers and organic agriculture. In Understanding Consumers of Food Products; Frewer, L., van Trijp, H., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 254–272. [Google Scholar]
- World Health Organization/Food and Agriculture Organization of the UN. Vitamin and Mineral Requirements in Human Nutrition, 2nd ed.; WHO: Bangkok, Thailand, 1998. [Google Scholar]
- Cordain, L.; Eaton, S.B.; Sebastian, A.; Mann, N.; Lindeberg, S.; Watkins, B.A.; O'Keefe, J.H.; Brand-Miller, J. Origins and evolution of the Western diet: Health implications for the 21st century. Am. J. Clin. Nutr. 2005, 81, 341–354. [Google Scholar]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets—iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar]
- Leonard, W.H.; Martin, J.H. Cereal Crops; MacMillan Publishing: New York, NY, USA, 1963. [Google Scholar]
- Worthington, V. Nutritional quality of organic versus conventional fruits, vegetables, and grains. J. Alternative Compl. Med. 2001, 7, 161–173. [Google Scholar]
- Murphy, K.M.; Reeves, P.G.; Jones, S.S. Relationship between yield and mineral nutrient concentrations in historical and modern spring wheat cultivars. Euphytica 2008, 163, 381–390. [Google Scholar]
- Hornick, S.B. Factors affecting the nutritional quality of crops. Am. J. Alternative Agr. 1992, 7, 63–68. [Google Scholar]
- Bourn, D.; Prescott, J. A comparison of the nutritional value, sensory qualities, and food safety of organically and conventionally produced foods. Crit. Rev. Food Sci. Nutr. 2002, 42, 1–34. [Google Scholar]
- Mäder, P.; Hahn, D.; Dubois, D.; Gunst, L.; Alföldi, T.; Bergmann, H.; Oehme, M.; Amadò, R.; Schneider, H.; Graf, U.; Velimirov, A.; Flieβbach, A.; Niggli., U. Wheat quality in organic and conventional farming: Results of a 21 year field experiment. J. Sci. Food Agr. 2007, 87, 1826–1835. [Google Scholar]
- Ryan, M.H.; Derrick, J.W.; Dann, P.R. Grain mineral concentrations and yield of wheat grown under organic and conventional management. J. Sci. Food Agr. 2004, 84, 207–216. [Google Scholar]
- Turmel, M.-S.; Entz, M.H.; Bamford, K.C.; Thiessen Martens, J.R. The influence of crop rotation on the mineral nutrient content of organic vs. conventionally produced wheat grain: Preliminary results from a long-term field study. Can. J. Plant Sci. 2009, 89, 915–919. [Google Scholar]
- Davis, J.; Abbott, L. Soil fertility in organic farming systems. In Organic Agriculture: A Global Perspective; Kristiansen, P., Taji, A., Reganold, J., Eds.; Comstock Publishing Associates: Ithaca, NY, USA, 2006; pp. 25–51. [Google Scholar]
- Shannon, D.; Sen, A.M.; Johnson, D.B. A comparative study of the microbiology of soils managed under organic and conventional regimes. Soil Use Manag. 2002, 18, 274–283. [Google Scholar]
- Siciliano, S.D.; Theoret, C.M.; de Freitas, J.R.; Hucl, P.J.; Germida, J.J. Differences in the microbial communities associated with the roots of different cultivars of canola and wheat. Can. J. Microbiol. 1998, 44, 844–851. [Google Scholar]
- Entz, M.H.; Guilford, R.; Gulden, R. Crop yield and soil nutrient status on 14 organic farms in the eastern portion of the northern Great Plains. Can. J. Plant Sci. 2001, 81, 351–354. [Google Scholar]
- Snyder, C.; Spaner, D. The sustainability of organic grain production on the Canadian Prairies—a review. Sustainability 2010, 2, 1016–1034. [Google Scholar]
- Bossio, D.A.; Scow, K.M.; Gunapala, N.; Graham, K.J. Determinants of soil microbial communities: Effects of agricultural management, season, and soil type on phospholipid fatty acid profiles. Microb. Ecol. 1998, 36, 1–12. [Google Scholar]
- Shepherd, M.A.; Harrison, R.; Webb, J. Managing soil organic matter—implications for soil structure on organic farms. Soil Use Manag. 2002, 18, 284–292. [Google Scholar]
- Lundquist, E.J.; Scow, K.M.; Jackson, L.E.; Uesugi, S.L.; Johnson, C.R. Rapid response of soil microbial communities from conventional, low input, and organic farming systems to a wet/dry cycle. Soil Biol. Biochem. 1999, 31, 1661–1675. [Google Scholar]
- Wander, M.M.; Hedrick, D.S.; Kaufman, D.; Traina, S.J.; Stinner, B.R.; Kehrmeyer, S.R.; White, D.C. The functional significance of the microbial biomass in organic and conventionally managed soils. Plant Soil 1995, 170, 87–97. [Google Scholar]
- Girvan, M.S.; Bullimore, J.; Pretty, J.N.; Osborn, A.M.; Ball, A.S. Soil type is the primary determinant of the composition of the total and active bacterial communities in arable soils. Appl. Environ. Microbiol. 2003, 69, 1800–1809. [Google Scholar]
- Nelson, A.G.; Quideau, S.; Frick, B.; Niziol, D.; Clapperton, J.; Spaner, D. Spring wheat genotypes differentially alter soil microbial communities and wheat breadmaking quality in organic and conventional systems. Can. J. Plant Sci. 2011. in press. [Google Scholar]
- Dalpé, Y.; Hamel, C. Arbuscular mycorrhizae. In Soil Sampling and Methods of Analysis, 2nd ed.; Carter, M.R., Gregorich, E.G., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 355–378. [Google Scholar]
- DeGrood, S.H.; Claassen, V.P.; Scow, K.M. Microbial community composition on native and drastically disturbed serpentine soils. Soil Biol. Biochem. 2005, 37, 1427–1435. [Google Scholar]
- Thavarajah, D.; Thavarajah, P.; Sarker, A.; Vandenberg, A. Lentils (Lens culinaris Medikus Subspecies culinaris): A whole food for increased iron and zinc intake. J. Agric. Food Chem. 2009, 57, 5413–5419. [Google Scholar]
- Thavarajah, D.; Vandenberg, A.; George, G.N.; Pickering, I.J. Chemical form of selenium in naturally selenium-rich lentils (Lens culinaris L.) from Saskatchewan. J. Agric. Food Chem. 2007, 55, 7337–7341. [Google Scholar]
- Steel, R.G.D.; Torrie, J.H.; Dickey, D.A. Principles and Procedures of Statistics: A Biometrical Approach; WCB McGraw-Hill: Madison, WI, USA, 1997. [Google Scholar]
- Zhao, F.J.; Su, Y.H.; Dunham, S.J.; Rakszegi, M.; Bedo, Z.; McGrath, S.P.; Shewry, P.R. Variation in mineral micronutrient concentrations in grain of wheat lines of diverse origin. J. Cereal Sci. 2009, 49, 290–295. [Google Scholar]
- Mishra, B.N.; Prasad, R.; Gangaiah, B.; Shivakumar, B.G. Organic manures for increased productivity and sustained supply of micronutrients Zn and Cu in a rice-wheat cropping system. J. Sustain. Agr. 2006, 28, 55–66. [Google Scholar]
- McCallum, B.D.; DePauw, R.M. A review of wheat cultivars grown in the Canadian prairies. Can. J. Plant Sci. 2008, 88, 649–677. [Google Scholar]
- Fan, M.S.; Zhao, F.J.; Fairweather-Tait, S.J.; Poulton, P.R.; Dunham, S.J.; McGrath, S.P. Evidence of decreasing mineral density in wheat grain over the last 160 years. J. Trace Elem. Med. Biol. 2008, 22, 315–324. [Google Scholar]
- Rawat, N.; Tiwari, V.K.; Singh, N.; Randhawa, G.S.; Singh, K.; Chhuneja, P.; Dhaliwal, H.S. Evaluation and utilization of Aegilops and wild Triticum species for enhancing iron and zinc content in wheat. Genet. Resour. Crop Evol. 2009, 56, 53–64. [Google Scholar]
- Al-Karaki, G.; McMichael, B.; Zak, J. Field response of wheat to arbuscular mycorrhizal fungi and drought stress. Mycorrhiza 2004, 14, 263–269. [Google Scholar]
- Cruz, C.; Green, J.J.; Watson, C.A.; Wilson, F.; Martins-Loução, M.A. Functional aspects of root architecture and mycorrhizal inoculation with respect to nutrient uptake capacity. Mycorrhiza 2004, 14, 177–184. [Google Scholar]
- Mohammad, M.J.; Malkawi, H.I.; Shibli, R. Effects of arbuscular mycorrhizal fungi and phosphorus fertilization on growth and nutrient uptake of barley grown on soils with different levels of salts. J. Plant Nutr. 2003, 26, 125–137. [Google Scholar]
- Mohammad, M.J.; Pan, W.L.; Kennedy, A.C. Chemical alteration of the rhizosphere of the mycorrhizal-colonized wheat root. Mycorrhiza 2005, 15, 259–266. [Google Scholar]
- Habte, M. The roles of arbuscular mycorrhizas in plant and soil health. In Biological Approaches to Sustainable Soil Systems; Uphoff, N., Ball, A.S., Fernandes, E.C.M., Herren, H., Husson, O., Laing, M., Palm, C., Pretty, J., Sanchez, P., Eds.; Taylor & Francis Group LLC: Boca Raton, FL, USA, 2006; pp. 129–148. [Google Scholar]
- Mäder, P.; Edenhofer, S.; Boller, T.; Wiemken, A.; Niggli, U. Arbuscular mycorrhizae in a long-term field trial comparing low-input (organic, biological) and high-input (conventional) farming systems in a crop rotation. Biol. Fertil. Soils 2000, 31, 150–156. [Google Scholar]
- Entz, M.H.; Penner, K.R.; Vessey, J.K.; Zelmer, C.D.; Thiessen Martens, J.R. Mycorrhizal colonization of flax under long-term organic and conventional management. Can. J. Plant Sci. 2004, 84, 1097–1099. [Google Scholar]
- Oehl, F.; Sieverding, E.; Mäder, P.; Dubois, D.; Ineichen, K.; Boller, T.; Wiemken, A. Impact of long-term conventional and organic farming on the diversity of arbuscular mycorrhizal fungi. Oecologia 2004, 138, 574–583. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nelson, A.G.; Quideau, S.A.; Frick, B.; Hucl, P.J.; Thavarajah, D.; Clapperton, M.J.; Spaner, D.M. The Soil Microbial Community and Grain Micronutrient Concentration of Historical and Modern Hard Red Spring Wheat Cultivars Grown Organically and Conventionally in the Black Soil Zone of the Canadian Prairies. Sustainability 2011, 3, 500-517. https://doi.org/10.3390/su3030500
Nelson AG, Quideau SA, Frick B, Hucl PJ, Thavarajah D, Clapperton MJ, Spaner DM. The Soil Microbial Community and Grain Micronutrient Concentration of Historical and Modern Hard Red Spring Wheat Cultivars Grown Organically and Conventionally in the Black Soil Zone of the Canadian Prairies. Sustainability. 2011; 3(3):500-517. https://doi.org/10.3390/su3030500
Chicago/Turabian StyleNelson, Alison G., Sylvie A. Quideau, Brenda Frick, Pierre J. Hucl, Dil Thavarajah, M. Jill Clapperton, and Dean M. Spaner. 2011. "The Soil Microbial Community and Grain Micronutrient Concentration of Historical and Modern Hard Red Spring Wheat Cultivars Grown Organically and Conventionally in the Black Soil Zone of the Canadian Prairies" Sustainability 3, no. 3: 500-517. https://doi.org/10.3390/su3030500