Accelerating Capoeira Regeneration on Degraded Pastures in the Northeastern Amazon by the Use of Pigs or Cattle



1
Faculty of Resource Management, HAWK—University of Applied Sciences and Arts, Büsgenweg 1a, 37077 Göttingen, Germany
2
Embrapa Amazônia Oriental, Travessa Dr. Enéas Pinheiro, s/n, Marco, Belém, PA 66095-903, Brazil
*
Author to whom correspondence should be addressed.
Sustainability 2019, 11(6), 1729; https://doi.org/10.3390/su11061729
Submission received: 19 February 2019
/
Revised: 8 March 2019
/
Accepted: 19 March 2019
/
Published: 21 March 2019
(This article belongs to the Special Issue Sustainable Agriculture and the Integrated Management of Agricultural Weeds)
Abstract
:In the humid tropics of Latin America, considerable proportions of agro-scapes are covered with degraded pastures that were taken over by dense weedy shrub canopies hampering further forest succession. While tree seeds are still constantly dispersed by bats and birds, these often do not reach the soil but got stuck in the dense shrubby vegetation. While manual up-rooting of weedy shrubs or tree replantation is too expensive, we tested if burrowing pigs or trampling cattle can enhance proportions of bare soils for fallow restoration. These hypotheses were tested in on-farm experiments at Igarapé-Açu, northeastern Pará. Soil-opening effects of ten pigs (40 days + nights) and ten oxen (40 overnight stays), respectively, were tested against manual clearing and control on three plots per treatment, respectively. Ground cover percentages of bare soil, weedy shrubs, grasses, and tree species were visually determined in 40 plots/treatment before and directly after treatments, and half a year later (n = 480 samples). Both animal treatments could not really match manual clearing (62%) but pigs reached above 36% bare ground cover, while cattle just 20%. As pigs are almost omnipresent on Amazonian smallholdings and even give a modest economic refund, the use of pigs is recommended to smallholders who want to break up the lush weed layers for the benefit of forest restoration.
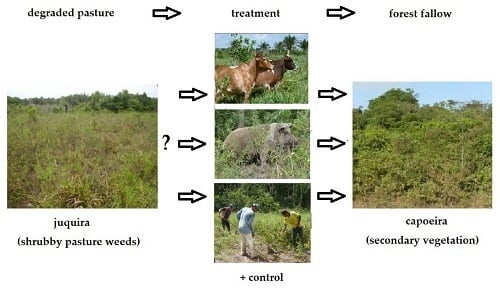
1. Introduction
In many humid tropical Central and South American regions, cattle pastures are ecologically not sustainable but enter into notorious degradation processes, leading to unproductive pastures after only 7–10 years of use [1,2,3]. Pasture degradation is thereby a complex agro-ecological process that is often early induced, e.g., by poor pasture establishment, and later by neglected controls and management activities, missing investments, or simply by mismanagement. Decreasing soil fertility, bush-encroachment, water deficiency, insect pests, and infection of fungi are the main ecological consequences of these shortcomings. In addition, technical aspects, such as inadequate forage grasses, overgrazing, inadequate fertilization, and underutilization of legumes, are responsible for early unproductiveness of pastures. Additionally, unfavorable socio-economic conditions like poor technical support, low return yields, inadequate development policies, and unsecure property rights lead to unsustainable cattle husbandry in these regions [2,3]. In the end, depleted soils, decreasing forage grass restoration and reiterating fencing costs force smallholders to abandon their pastures because further investment is not profitable anymore, even in short terms [3,4,5,6].
However, degraded pasturelands are difficult to turn into another land use, be it for another agronomic purpose or for forest use [3,7,8,9]. As manual restoration activities are generally too laborious, economically risky, and expensive [10,11], smallholders often simply abandon these degraded plots, trusting in the substantial biomass accumulation of the lush forest fallows in the humid tropics [7,12,13]. This secondary vegetation might develop into valuable forest fallows again [7,8,12,13,14,15]. After a few years, they will also be an effective tool to get rid of noxious agricultural weeds, which get shaded out by forest fallows [16]. Naturally, the dispersion of seeds by animals like bats, birds, and/or rodents is responsible for forest restoration [14,17,18,19,20,21,22,23]. However, this natural process can be quite slow, even in the tropics, especially when lofty isolated trees get uprooted and the next forest patches are far away. These plots in fragmented forest landscapes often need 25 years at most and up to 40 years to produce a useful amount of biomass for a subsequent cropping phase and to insure a complete suppression of shrubby weeds [4,7,23,24,25,26].
Besides time since abandonment, the overall pattern of forest restoration on degraded pastures is mainly related to the intensity of land-use history and decreasing distance to the next large forest patches [7,17,23,27]. Restoration happens due to slow secondary forest regeneration and its respective above-ground biomass accumulation, especially by spontaneously growing legumes that partly fix atmospheric N in the soils (Table 1a). Thus, natural vegetation succession should be artificially accelerated or even bypassed by farming activities, for instance by planting tree species or multi-purpose legumes [6,28,29,30,31,32]. First experimental trials in that direction showed promising results by using woody legumes that showed remarkable establishment rates even under unfertilized conditions [11,17,33,34,35]. However, one main drawback of this strategy is that legume planting includes high-input activities and are expensive [10,11,33,36]. Another problem is that plantation of exotic legumes would not restore the high biodiversity of the natural secondary vegetation with its diverse ecological adaptations and regional important ecological services [37,38]. Thus, there is still a need for cheap low-input technologies on smallholdings to bring degraded pastures back into the phytodiverse fallow-based smallholder production systems.
Therefore, the objective of this study is to restore the traditional forest fallow as the key agro-ecological basis for smallholder agriculture in the Amazon and perhaps the remainder of the humid Neotropics with low-input methods [12,13,39]. Traditionally, the whole sustainability of traditional smallholder agriculture in the Amazon fundamentally depends on this secondary forest, nationally called “capoeira” in Brazil. Its agricultural value is due to its dense woody regrowth of fast-growing secondary forest trees, herbaceous and woody lianas, shrubs, perennials, and herbs [12,33,37,40]. The prolific and mega-phytodiverse vegetation can accumulate up to 30 t/ha dry matter above-ground biomass in just four years and is traditionally slashed and burned or mulched for a subsequent cropping phase [40,41,42,43]. Thereby, the capoeira provides important habitat function for the indigenous flora and fauna and fulfills important ecological services in rural Amazonian regions [37,42,44]. Some of the most frequent capoeira species from the Bragantina region [12,37,45,46] are listed in Table 1b.
Traditionally, pastures without the possibility of re-sprouting capoeiras are defined as “biologically degraded pastures” [3,45]. They are ecologically characterized by chemically and physically depleted soils with low nutrient supply (especially in P and N), low cation exchange capacities, and high soil compaction [4,7,14,25]. The capoeira is almost completely uprooted and rather unpalatable native grasses, perennial herbs, herbaceous lianas, and shrubs that do not reach the biomass production of capoeira species are invading the areas [3,47,48]. This agricultural weed vegetation formation is locally called “juquira” in the Bragantina region. The most important juquira species (weedy shrubs) on pastures in the northeastern Amazon are Borreria verticillata (L.) G. Mey. (Rubiaceae) and Myrciaria tenella (DC.) O. Berg (Myrtaceae) but also contain other taxa (Table 1c). Unfortunately, biologically degraded pastures are not easily identifiable by remote sensing (patchy distribution; a similar spectral signature as intact pastures), but first attempts suggest that 8.6% (41,572 ha) of Bragantinian agro-scapes are covered with these weedy shrubs [49]. On these biologically degraded pastures, the main problem is often that arriving tree seeds never reach the ground for germination but are entrapped in the dense shrubby vegetation and thick litter layers [7,14,19,20,21,22,23,25,29,50,51,52,53].
However, experiences from other neotropical regions, e.g., on montane pastures in Colombia [54], showed that a new short-time overgrazing of cattle create small weed gaps and thereby facilitate seed establishment of capoeira trees. This low-input option can be intensified by keeping cattle overnight or by using other domestic animal species like goats, sheep, or pigs. Domestic pigs, for instance, are well known to open soils by their intensive burrowing activities, also in the humid tropics [55,56,57]. In Pará, there are almost 560,000 pigs that are distributed on 153,273 smallholdings. A typical example of the rural landscape is the municipality of Igarapé-Açu where 689 pigs on 103 farms during the livestock census 2006 and 2014 were registered [58]. These numbers elucidate that pigs are commonly present on smallholdings and are an integral part within the northeastern Amazonian smallholder production systems. Thus, ecological services of these omnipresent pigs can be theoretically used on farms, for instance for the benefit of fallow restoration.
Thus, we assume that the ecological services of domestic animals (cattle, pigs) can substitute expensive manual hoeing by farmers to accelerate capoeira restoration. In more detail, we hypothesize that pigs and cattle increase the portion of bare soil on the plots to the same extent as manual clearing by day laborers. Furthermore, besides predicting that pigs are more efficient soil-breakers than cattle, we also hypothesize that they reduce soil cover of grasses and the shrub-layers, and especially that of B. verticillata and M. tenella. Even more, we feared that pig effects could be so strong that the animals would even damage and/or remove the desired tree saplings and would reduce their ground cover percentages. In terms of soil bulk density, we hypothesize that cattle increase soil compaction by trampling and values are also higher than control, while pigs and manual clearing will loosen the upper soil layers and values will be lower than control.
2. Materials and Methods
2.1. Study Area and Experimental Design
To test these hypotheses, cattle (trt1) and pig treatments (trt2) were tested against manual clearing (trt3) and control (trt4; Figure 1 and Figure 2). The experiment was replicated on three different smallholder farms (blocks) in the center of the Bragantina region, at Igarapé-Açu (1°03’ S and 47°30’ W), northeastern Pará, northern Brazil (n = 3 farms; n = 12 plots; n = 480 subplots). The region belongs to the per-humid warm low-land tropics, with a mean annual temperature between 21.0–22.3 °C and rainfall between 2000–2847 mm/year [12,37]. The rainy season lasts from January until June, while the dry season from July until December. The driest months October and November have less than 50 mm, respectively [37]. During the experimental time (August 2007 until May 2008), typical temperatures and rainfall patterns were observed. However, November, not October as usual, was the driest month in 2007 with just 2.6 mm rainfall. Therefore, December received more than the double amount of rainfall than the mean (231 mm instead of 110), and April 565 mm instead of 400 mm (mean), while the other months of the rainy season showed values minimally below the annual mean [59,60]. Typical soils of the region are poor Oxisols, Ultisols, and Entisols, characterized by a mean pH of 4.5, low nutrient fertility, especially in P and N, and a low cation exchange capacity [12,41,42,61].
Between August 2007 and May 2008, on each block (replication farm), 2 ha big plots of relatively homogenous biologically degraded pastures were selected and subdivided into four 0.5 ha (50 × 100 m) plots for treatment establishment. All plots had the similar land use history, typical for the Bragantinian rural region: after slashing-and-burning 4–10 years old capoeiras, crops like maize or beans had been cultivated for one or one-and-a-half years. Thereafter, traditional mixed Brachiaria-pastures were established which were then yearly slashed back from infesting juquira. As this was done for the last 8–10 years, forage grasses and capoeira trees were also increasingly eliminated by slashing and burning, so that mainly the robust juquira vegetation remained on the plots. Consequently, our selected plots were mainly dominated by B. verticillata and M. tenella shrub cover (Figure 1d). All treatments were fenced with five strands of barbwires, just trt2 receiving three strands more at the bottom to secure pigs on the plots. Animals were supplied with water and fodder ad libitum (pig fodder up to 7 kg/day plus kitchen scraps of the respective smallholding; around 0.5 kg/day for piglets). Palm-roofed shelters for pigs were located in one corner of the respective plots.
For trt1, ten mixed-bred cattle, including phenotypic characteristics of Holstein-Friesian, Nelore, and Gir breed (Figure 1a), with a mean body mass of around 400 kg, were put for 40 nights on the subplot in November 2007, between 6:00 p.m. until 8:00 a.m. Each block had its own herd of cattle and stocking rates were at 20 animals/ha (800 kg/ha) for the nighttime. The intention was that, during the early morning hours and also during the resting evening hours, cattle would roam through the degraded pasture to graze the remaining forage grasses and provoke a breaking-up of the juquira layers. During the days, animals grazed outside the experiment on distant pastures.
Trt2 received a pig family of domesticated pigs of the local Landrace breed of Pará, characterized by black skin spots (Figure 1b), for 40 days and nights. Three different pig families were put on the respective block in November 2007, December 2007, and January 2008. The pig family consisted of 11.7 animals (mean), with 2–3 adults, 3–4 piglets, and 5–6 baby piglets. Pigs were not withdrawn from plots in the daytime like cattle, as this was inoperable for farmers. Cattle could not stay on the degraded pasture plots during the daytime, as there were not sufficient forage grasses for animal alimentation. Trt3 was manually cleared by day laborers with the help of hoes (Figure 1c). Workers were advised to open the soil and uproot the weedy shrubs but neither to eradicate young capoeira saplings, nor lofty isolated seed trees (Figure 1c) nor the spontaneous herbaceous legume layer. Litter remained on the plots. The number of workers and hours/ha are indicated in the result chapter. Trt4 was control and was not altered. Plots were not fenced, as accidentally roaming cattle or wild herbivores belong to this traditional smallholder treatment. However, wild herbivores are extremely rare in the region.
2.2. Vegetation Sampling
The following six parameters were raised by estimating ground cover percentages of (a) bare soil, (b) B. verticillata, (c) M. tenella, (d) grasses- including remaining Brachiaria forage grasses but also all native spontaneously growing grasses (Table 1c), (e) native herbaceous legumes (cf. Table 1a), and (f) shrubs—all other weedy shrubs, lianas, and herbs, excluding B. verticillata, M. tenella, grasses, and herbaceous legumes. All variables were sampled at three times: time1 = before treatment from August until October 2007, time2 = directly after treatment effects in February–March 2008, and time3 = half a year after last treatment impact, in September–October 2008. Additionally, the number of tree individuals was counted, the mean of the heights of the five highest capoeira trees were calculated, and the ground cover percentage of native herbaceous legumes were estimated at time1 and time2. These variables were used as an indicator for possible animal damages on the young capoeira during treatments. Due to the high heterogeneity of the juquira and capoeira portions on degraded pastures in general, an extraordinary high number of 40 sampling subplots, i.e., four random transects of ten single 6.25 m2 sampling plots (2.5 × 2.5 m), with at least 5 m distance from fences and each other, were sampled for each parameter (Figure 2). Ground cover of bare soil and vegetation were visually estimated and categorized into eighth classes: 0.1–12.5%; 12.6–25%; 25.1–37.5%; 37.6–50%; 50.1–62.5%; 62.6–75%; 75.1–87.5%; 87.6–100%). Each class received a percent value for further calculation (the mean of the class): class1 = 6.25%; class2 = 18.75; class3 = 31.25; class4 = 43.75; class5 = 56.25; class6 = 68.75, class7 = 81.25; class8 = 93.75. To have a rough proxy for seed survival success (cf. 52), the number of capoeira saplings (<15 cm height) were also counted within the sampling plots at time2.
2.3. Soil Compaction
We measured soil compaction effects of the four treatments by using the hand penetrometer to 1 m (Eijkelkamp, Giesbeek, The Netherlands). In each corner of the 12 subplots, the penetrometer was pushed three times into the soil within a 30-cm diameter circle. To avoid experimental border effects, we kept a distance of at least 10 m from fences, and measured during time1 and time3 on all treatments. As we expected just significant animal effects within the upper soil layers, data of 2.5, 5, 10, 15, 20, and 40 cm depths were read out from paper sheets (n = 893). As thicker tree roots were still quite common on the plots, values of ≥5 MPa were removed from statistical analyses. Density data were analyzed for means, standard errors, and significant differences at p < 0.05. For each other parameter, the mean values plus standard deviation were calculated and differences were calculated for significance (p < 0.05) by conducting ANOVAs between treatments and times, using the “aov-procedure” of the interactive statistics platform “R” [62]. The following fixed linear model was used to represent the variation of differences:
where Ytfi = cover of bare soil, shrubs, B. verticillata, M. tenella, grasses, capoeira, legumes, number of trees, and tree heights:
Ytfi = μ + att + bf + ci + abtf + acti + dtfi,
- µ = overall mean,
- at = effect of treatment (i = trt1, trt2, trt3, trt4),
- bf = block effect (j = farm1, farm2, farm3),
- ci = effect of time (k = time1, time2, time3),
- abtf = interaction of treatment and block effect,
- acti = interaction of treatment and time effect,
- dtfi = residual deviation.
For the variables “legumes”, “number of trees”, and “tree heights”, data were determined just for time1 and time2.
2.4. Economic Costs and Benefits
It was originally not intended to completely compare economical features of all treatments in this study. However, to show a rough proxy of the slight economic gains of pig production, these animals were weighed before and after treatment, calculating also total weights/0.5 ha. Values were then multiplied by 2.5 R$ (BRL), the actual pig price per kg in October 2007, additionally multiplied by 2 to reach gains per hectare, and multiplied by 0.5 to get the economic gains in US$ (USD). The currency exchange rate was taken on 30th of August 2007 [63]. Pigs needed three more strands of barbwires to be held on degraded pastures, which needed two men for 30 minutes more than for the other treatments. These additional fencing costs were calculated with 0.94 US$ for each block (15 R$/8 working hours = 0.94 R$ for half-an-hour, multiplied by two day laborers) and then subtracted from the economic gains of each block. Costs for manual clearing were calculated by counting the needed working days multiplied by 15 R$ daily wage and the number of day laborers for the respective farm in US$. It was not useful to surveil cattle weight gains, as these animals spend the daytime on non-standardized different pastures and because cattle gains were not of interest in this ecological experiment.
3. Results
The results of the ground cover changes of the most important parameters under treatments are shown in Figure 3. Comparing the four treatments of the bare soil values (Figure 3a) against each other at time2 showed that all treatments had significant different values (p < 0.0001). As expected, day laborers achieved the highest clearing rates. However, as young target trees should be spared from clearing and not all inhibiting shrubs could have been completely eliminated in practice, values reached not more than 62.4%. Pigs reached a remarkably high value of 36.3% at time2. However, they could not match the trt3-values (p < 0.0001), meaning that our first hypothesis is rejected. Surprisingly, the soil opening effect of trt3 did not last for long, but values dropped back to below 20% at time3 and similar to that of trt1 (p = 0.611). However, trt3 at time3 was also significantly different from the value of time1 (p = 0.0003). In line with the bare soil parameter, some shrub-values decreased significantly, i.e., by trt3 and trt2 (Figure 3b). Trt2 did not eradicate the shrubs to the expected extent and reached just values equal to trt1 (p = 0.0692). However, while trt1 impact did not get significantly different between time1 and time2 (p = 0.2), this was the case for trt2 (p < 0.0001). Regarding the values of the B. verticillata-cover (Figure 3c), almost all values became significantly different between time1 and time2 (p < 0.0001). Just the values of trt4 remained statistically the same during the three times (all p > 0.26). In addition, trt2 stayed at the same level between time2 and time3 (p = 0.261). Besides trt3, there were no significant treatment effects observed on robust M. tenella-shrubs (Figure 3d). Trt1 and trt2 showed the same disappointing results and were significantly higher than that of trt3. Besides trt4, which stayed at the same level from time1 until time3, all grass treatments (Figure 3e) became significantly different at time2, albeit trt1 and trt4 that were quite similar (p = 0.0271). However, all grass cover values of the three treatments recuperated fast, and trt1-trt3 between time2 and time3 were all significantly different (trt1: p = 0.000129), so that all values became significantly higher than trt4 at time3. There were some observations of grass sods, turned by pigs, starting to spread roots again. Unfortunately, pigs dug up some valuable young target capoeira trees while burrowing up the plots, as trt2-values of the capoeira-data (Figure 3f) decreased from 42.9% to 34.1%. Thus, trt2 was the only treatment that had an almost significant impact on the capoeira (p = 0.000909). However, this seems to be just a side effect, as pigs did not intentionally destroy the target species but searched also the forested parts of the plots, after a few days.
As it is predictable that this disturbance effect gets stronger the longer the animals stay on the plots, the animals should be withdrawn from the plots when the juquira is almost uprooted because there seems to be no other practical solution to keep the pigs from young desirable capoeira trees. However, young trees recuperated fast after impact on trt2 and reached the level of time1 (44.9%; p = 0.451). Day laborers paid enough attention to sparing young target trees, which was not always an easy task, as B. verticillata and/or M. tenella weedy shrubs sometimes grew together with capoeira trees at the same location.
At time1, on all treatments, the same number of tree individuals per 6.25 m2 has been counted, except for trt3, which was significantly lower than trt2 (Table 2). Trt4 surprisingly decreased its values to 42%, probably due to the naturally closing shrub layers. The number of capoeira saplings was significantly lower on trt4, while all other numbers of saplings stayed at the same level (p > 0.05). Around 80% of the observed saplings of all treatments were of Vismia guianensis (Aubl.) Choisy (Clusiaceae) and around 5% of Lacistema pubescens Mart. (Lacistemataceae). Ground covers of herbaceous legumes were quite heterogeneous, indicated by the different values of the time1-values and high standard deviations (Table 3). However, after treatments, all legume values showed the same ground cover around 14.5% except for trt3, which showed almost the double amount (29.2%), as day laborers should spare legumes. On trt4, legume cover decreased significantly which might be due to out shading effects of the growing values of shrubs, M. tenella, and capoeira covers (cf. Figure 3). Results of soil compaction showed a significant increase of values with time, except the values in 2.5 cm of control and in 40 cm depth of trt2 (Table 4). Trt1 showed significant (p < 0.05) surface compaction after treatment in the upper 2.5 cm-soil layers. Trt2 showed significant higher not lower values, and values after treatment were statistically the same as trt4 (all p > 0.05). The number and mean weights of pigs are shown in Table 5. After the subtractions of the additional barbwire strand costs of 0.94 US$, therefore, the mean economic gain of the three farms was 682.39 US$/ha, reaching from 506.56 US$ to farm2 until 814.06 US$/ha on farm3.
Three day laborers on farm1 needed nine days to do the work of trt3 (=405 R$), while on farm2 just four day laborers spent six days (=360 R$), and on farm3 three day laborers seven days (=315 R$). Thus, there were mean costs of 360 US$/ha with a standard deviation of 45 US$. Trt1 needed just some financial investments for the joint installation of the whole experiment. However, in reality, there would be just minor costs for farms, as cowboys just had to get cattle from the degraded plots in the morning and bring them back in the evening. This action would consume at maximum one hour per day, morning and evening herding together, which would imply (15 R$/8 working hours = 1,88 R$ per day), again 0.94 US$ extra-costs per day.
4. Discussion
The results mean that the use of domestic pigs, to break up the dense weed canopies (juquira), can be a promising and cheap option for Amazonian smallholders. Although pigs could not entirely match manual clearing with hoes, their impacts on bare soils and the weedy shrub vegetation were considerable in major parts of the plots which, however, is not so striking in the presented data. The presentation problem was that pigs burrowed the soil not as evenly as day laborers but were most active along fences and in corners of the plots. Here, they cleared almost the complete area, showing strong border effects. However, as agricultural experimental stations, in general, are designed to try to blank out border effects, like it was also intended in this trial, the burrowing effects of pigs are therefore underrepresented in the data. Next to corners and fences, the animals showed also excellent clearing results around their shelters and along animal tracks, which was done as efficiently as by manual hoeing. While human activity is rather expensive, pigs returned at least a modest economic gain during the same time by gaining weight. However, these gains are mainly caused by additional feed supply that was provided, as it is usually done on smallholdings. It remains unclear how much pigs really foraged from degraded pastures and how much the provided feed contributed to weight gains. Data also indicate that trt2 kept open the bare soil patches for a much longer time than trt1 or even trt3, enlarging the period of favorable conditions for sexual reproduction of the capoeira trees, i.e., seed-germination and seedling growth of target tree species.
Vegetation succession and development after trt2 are well known to be highly dynamic and heterogeneous and usually showed a significant increase of species richness [55,56,57,64]. Some experiences from the humid tropics showed that pig-scarification can even lead to an establishment of special pioneer tree species [57,65]. However, most of the benefiting plants are noxious weeds that are well adapted to unstable and dynamic environments [56,57,64]. Although grass and B. verticillata-layers were significantly reduced, trt1 impact was negligible and open spots were distributed rather patchy on the plots. Therefore, ecological effects were not strong enough to induce considerable open areas for tree regeneration, and, on the few open patches, soils got more compacted by trampling or resting cattle (Figure 4a; see below). It remains to be seen if the additional dung import from adjacent pasture grazing during the day will enhance at least capoeira seed rain via cowpats. Nutrients, however, are expected to be imported by both domestic animal species, in the case of pigs via fodder or by the organic household wastes.
Our soil compaction data revealed the well-known trampling effect of cattle that lead to compacted upper soil layers [14]. However, soil compaction by trt1, as made evident by soil physical properties, can be accepted just for the 2.5 cm variables, i.e. surface compaction. It should be also noticed that trt1 started from the lowest value of all treatments in the 2.5 cm-layer (e.g., 1.88 MPa). Trt2 did not show the desired effects, as soil compaction was higher after treatment effect and the same as control (all p > 0.05). This trt2-effect can be probably explained by the above-mentioned border effects, as pigs preferred to mainly burrow along fences and started from slightly higher values (Figure 4b). Values of manual clearing were mostly in between the other treatments but showed much higher values in the 5-cm layer, probably due to the fact that the uppermost soil layer was scratched by day laborers to eliminate the juquira (Figure 4c). Significant higher compaction values on all treatment might be explained by season, as soil compaction often depends on the soil moisture contents and thus on rainy or dry seasons [14,19,42]. The period for seed germination and establishment of target forest trees seem to be limited under all treatments, indicated by the distinctively re-bouncing shrub-values at time3. This was found also in other studies, where livestock grazing did change soil infiltration rates, soil bulk density, and soil porosity, but the effects were quickly reversed following cessation of grazing and had little detrimental effect on tree production [66]. The present study aimed to evaluate the chances for germination and sprouting of seedlings and sapling from seeds, which depends on the conditions in which the land was abandoned after the agricultural cultivation (juquira phase). The further vegetal succession progress [67] should be monitored through the coming years (conclusion section).
Extra-fencing with three additional strands of barbwires was necessary to hold pigs on the plots, needing extra labor/money of farmers. However, in practice, even five wires were not able to completely avoid escapes from plots. Especially, piglets can hardly be secured but also mature pigs tried to refuge the longer they stayed there. However, while piglets willingly come back later to their mother pigs, mature pigs will not return and can cause severe damages on adjacent fields. Additionally, once pigs escape, it is difficult to catch them again and it often takes half a day for three agricultural workers at a minimum to bring all escaped animals back to plots. Once a plot is sufficiently burrowed, shifting to the next plots is necessary and causes extra-fencing, pig catching, and labor time again. However, the modest economic gains can be used to compensate parts of these additional costs. However, as management time is always scarce on smallholdings and pastures have to be checked almost on a daily basis, it will be a difficult task for busy Amazonian peasant farmers to optimize the ecological services of these animals.
Another promising alternative animal species for the desired ecological effects is perhaps the Collared Peccary (Pecari tajacu L., Tayassuidae, Portuguese: cateto) that might provide even better ecological effects and production features than domestic pigs [68,69]. However, as there are almost no practical experiences with the species on-farm, and animals can cause serious damage in case of escapes in the agro-scapes, or can even be dangerous for people, this idea was disregarded for this experiment. However, we would like to recommend research upon this species, hypothesizing that P. tajacu might create higher proportions of small gaps in weed canopies (juquiras) than domestic pigs, while taking better advantage of the nutrient-poor natural resources on degraded tropical pastures than the Landrace pigs. It assumedly will receive higher prices on local markets, and will be better adapted to the ecological conditions of the northeastern Amazon [68,69].
Instructing day laborers to hoe the shrubs was easy and there were no further problems to slash the shrubs while sparing the capoeira trees and legumes. Difficulties just occurred when weedy shrub patches grew in conjunction with valuable capoeira trees so that there was the danger to damage target trees while withdrawing disturbing shrubs. However, clearing activities can be weary after some hours so that tired workers unintentionally damage target forest trees. In practice, it is not possible to completely eliminate shrubs from plots, as some up-rooted shrubs start to root again. However, removing the shrub litter from plots is too expensive. Initially, all replication blocks seemed to have the same degradation stage, fully covered with B. verticillata and M. tenella and very few capoeira trees. However, at time3, it became clear that farm1 had much more capoeira trees rooting firmly in the ground, while farm3 almost had none.
5. Conclusions
The proposed agricultural technique to use pigs or cattle to break up the juquira layers for accelerated capoeira restoration is a low-input method and thus suitable for smallholders of the NE-Amazon. Large agricultural enterprises will prefer to plant tree saplings or to dig up and fertilize lands with big agricultural implements, but these high-input agricultural management tools are hardly acceptable for smallholders, as the direct financial return is often low. However, as generally reforestation techniques are not easily realizable for Amazonian peasant farmers, adoption rates of this technique might be critical, especially if the success is not fully guaranteed after one year or even endangered to be destroyed by uncontrolled fires of neighboring farms [11,41]. Moreover, in the humid tropics, smallholders usually have to combat against the lush vegetation and a high amount of labor and money is permanently spent to keep the vegetation under control. Thus, motivation to invest labor and money to combat the aggressive and fast growing weed canopies (juquira) aiming at fostering a successful establishment of capoeira on fallow land is limited. However, a kick-off action to break once the weedy shrub canopies might be acceptable for smallholders if an obvious acceleration of the establishment of capoeiras can be achieved. Because once smallholders understood the importance of agricultural systems as multifunctional landscapes, they also will be convinced about the necessity to invest in the agronomic and economic benefits of secondary forest fallow dynamics [35]. This is all the more applicable because pigs are almost omnipresent on Amazonian smallholdings [58] and farmers like to keep them. Besides protecting their areas from fire [8,41,43], the investigation into the capoeira will not only secure livelihoods of farmers [13], but will also substantially contribute to the conservation of the indigenous flora and fauna [37,44]. At first glance, the results are just valid for a limited region in the center of the Bragantina region. However, our approach is in best accordance with global efforts of ‘provisioning, regulating and maintaining cultural ecosystem services’ [70] that might be suitable also for other Amazonian regions or beyond, in the humid neotropics [71]. Finally, the further vegetal succession process, subsequent to the four different starting conditions/treatments of juquira, requires the implementation of standardized methods of surveillance and monitoring to assess the progress of further successional phases (capoeira rala—actual capoeira—capoeirão) [67], until the stage of the secondary forest reaching the maximum succession is completed.
Author Contributions
S.H. and H.W. wrote the paper; S.H. sampled data in the field; O.R.K. supervised data sampling and gave substantial expert input.
Funding
The work has been funded by the German Research Foundation (WO-913/2-2) with manifold support of the Embrapa Amazônia Oriental in Belém do Pará. We also acknowledge the support of the Open Access Publication Funds of the HAWK, Hildesheim/Holzminden/Göttingen, Germany.
Acknowledgments
We thank Ednaldo Augusto Pinheiro Nascimento, Marildo Teixeira da Silva, and Jucelino Machado for technical support and the Tipitamba team (Embrapa) in Belém for the collaboration and discussions. We are especially grateful to Reginaldo da Silva Coelho, José Sténio Mendes Machado, and E.C. Santana for permitting the experiments on their farms.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Serrão, E.A.S.; Falesi, I.C.; da Veiga, J.B.; Teixeira Neto, J.F. Productivity of Cultivated Pastures on Low Fertility Soils of the Amazon Brazil. In Pasture Production in Acid Soils of the Tropics; Sánchez, P.A., Tergas, L.E., Eds.; Centro Internacional de Agricultura Tropical: Cali, Colombia, 1979; pp. 195–225. [Google Scholar]
- Serrão, E.A.S.; Nepstad, D.C. Pastures on Amazonian forestlands: A review of environmental and economic performance. In Interdisciplinary Research on the Conservation and Sustainable Use of the Amazonian Rainforest and Its Information Requirements; Lieberei, R., Reisdorff, C., Dantas Machado, A., Eds.; SBCS: Geesthacht, Germany; Stern-Werke: Hamburg, Germany, 1996; pp. 221–239. ISBN 3-00-000909-4. [Google Scholar]
- Dias-Filho, M.B. Degradação de pastagens: Processos, causas e estratégias de recuperação, 4th ed.; MBDF: Belém, Brazil, 2015. [Google Scholar]
- Buschbacher, R.; Uhl, C.; Serrão, E.A.S. Abandoned pastures in Eastern Amazonia. II. Nutrient stocks in the soil and vegetation. J. Ecol. 1988, 76, 682–699. [Google Scholar] [CrossRef]
- Müller, M.M.L.; Guimarães, M.F.; Desjardins, T.; Mitja, D. The relationship between pasture degradation and soil properties in the Brazilian Amazon: A case study. Agric. Ecosyst. Environ. 2004, 103, 279–288. [Google Scholar] [CrossRef]
- Davidson, E.A.; Reis de Carvalho, C.J.; Figueira, A.M.; Ishida, F.Y.; Ometto, J.P.H.B.; Nardoto, G.B.; Sabá, R.T.; Hayashi, S.N.; Leal, E.C.; Vieira, I.C.G.; et al. Recuperation of nitrogen cycling in Amazonian forests following agricultural abandonment. Nature 2007, 447, 995–998. [Google Scholar] [CrossRef]
- Uhl, C.; Buschbacher, R.; Serrão, E.A.S. Abandoned pastures in Eastern Amazonia. I. Patterns of plant succession. J. Ecol. 1988, 76, 663–681. [Google Scholar] [CrossRef]
- Aide, T.M.; Zimmerman, J.K.; Pascarella, J.B.; Rivera, L.; Marcano-Vega, H. Forest Regeneration in a Chronosequence of Tropical Abandoned Pastures: Implications for Restoration Ecology. Rest. Ecol. 2000, 8, 328–338. [Google Scholar] [CrossRef]
- Lamb, D.; Erskine, P.D.; Parrotta, J.A. Restoration of degraded tropical forest landscape. Science 2005, 310, 1628–1632. [Google Scholar] [CrossRef]
- Bruel, B.O.; Marques, M.C.M.; Britez, R.M. Survival and Growth of Tree Species under Two Direct Seedling Planting Systems. Rest. Ecol. 2010, 18, 414–417. [Google Scholar] [CrossRef]
- Bertacchi, M.I.F.; Amazonas, N.T.; Brancalion, P.H.; Brondani, G.E.; Oliveira, A.; Pascoa, M.A.; Rodrigues, R.R. Establishment of tree seedlings in the understory of restoration plantations: Natural regeneration and enrichment plantings. Rest. Ecol. 2015, 24, 100–108. [Google Scholar] [CrossRef]
- Denich, M. Estudo da importância de uma vegetação secundária nova para o incremento da produtividade do sistema de produção na Amazônia oriental brasileira, 2nd ed.; GTZ, EMBRAPA/CPATU: Eschborn, Germany; Belém, Brazil, 1991. [Google Scholar]
- Chazdon, R.L. Second Growth: The Promise of Tropical Forest Regeneration in an Age of Deforestation; University of Chicago Press: Chicago, IL, USA, 2014. [Google Scholar]
- Holl, K.D. Factors Limiting Tropical Rain Forest Regeneration in Abandoned Pasture: Seed Rain, Seed Germination, Microclimate, and Soil. Biotropica 1999, 31, 229–242. [Google Scholar] [CrossRef]
- Feldpausch, T.R.; Rondon, M.A.; Fernandes, E.C.M.; Riha, S.J.; Wandelli, E. Carbon and nutrient accumulation in secondary forests regenerating on pastures in central Amazonia. Ecol. Appl. 2004, 14, 164–176. [Google Scholar] [CrossRef]
- De Rouw, A. The fallow period as a weed-break in shifting cultivation (tropical wet forests). Agric. Ecosyst. Environ. 1995, 54, 31–43. [Google Scholar] [CrossRef]
- Nepstad, D.C.; Uhl, C.; Pereira, C.A.; Cardoso da Silva, J.M. A comparative study of tree establishment in abandoned pasture and mature forest of eastern Amazonia. Oikos 1996, 76, 25–39. [Google Scholar] [CrossRef]
- Uhl, C.; Nepstad, D.; da Silva, J.M.C.; Vieira, I. Restauração da floresta em pastagens degradadas. Ciência Hoje 1991, 13, 23–31. [Google Scholar]
- Aide, T.M.; Cavelier, J. Barriers to lowland tropical forest restoration in the Sierra Nevada de Santa Marta Colombia. Rest. Ecol. 1994, 2, 219–229. [Google Scholar] [CrossRef]
- Da Silva, J.M.C.; Uhl, C.; Murray, G. Plant Succession, Landscape Management, and the Ecology of Frugivorous Birds in Abandoned Amazonian Pastures. Conserv. Biol. 1996, 10, 491–543. [Google Scholar] [CrossRef]
- Muscarella, R.; Fleming, T.H. The role of frugivorous bats in tropical forest succession. Biol. Rev. 2007, 82, 573–590. [Google Scholar] [CrossRef]
- Reid, J.L.; Holl, K.D.; Zahawi, R.A. Seed dispersal limitations shift over time in tropical forest restoration. Ecol. Appl. 2015, 25, 1072–1082. [Google Scholar] [CrossRef] [Green Version]
- Aide, T.M.; Zimmerman, J.K.; Herrera, L.; Rosario, M.; Serrano, M. Forest recovery in abandoned tropical. pastures in Puerto Rico. For. Ecol. Man. 1995, 77, 77–86. [Google Scholar] [CrossRef]
- Engel, V.L.; Parrotta, J.A. An evaluation of direct seeding for reforestation of degraded lands in central São Paulo state Brazil. For. Ecol. Man. 2001, 152, 169–181. [Google Scholar] [CrossRef]
- Zahawi, R.A.; Augspurger, C.K. Early plant succession in abandoned pastures in Ecuador. Biotropica 1999, 31, 540–552. [Google Scholar] [CrossRef]
- Aide, T.M.; Zimmerman, J.K.; Rosario, M.; Marcano, H. Forest recovery in abandoned cattle pastures along an elevational gradient in northeastern Puerto Rico. Biotropica 1996, 28, 537–548. [Google Scholar] [CrossRef]
- Uhl, C. Factors controlling succession following slash-and-burn agriculture in Amazonia. J. Ecol. 1987, 75, 377–407. [Google Scholar] [CrossRef]
- Lindell, C.A.; Reid, J.L.; Cole, R.J. Planting design effects on avian seed dispersers in a tropical forest restoration experiment. Rest. Ecol. 2013, 21, 515–522. [Google Scholar] [CrossRef]
- Nepstad, D.C.; Uhl, C.; Serrão, E.A.S. Surmounting barriers to forest regeneration in abandoned, highly degraded pastures: A case study from Paragominas, Pará, Brazil. In Alternatives to Deforestation: Steps toward Sustainable Use of the Amazon Rain Forest; Anderson, A.B., Ed.; Columbia University Press: New York, NY, USA, 1990; pp. 215–229. [Google Scholar]
- Nepstad, D.C.; Uhl, C.; Serrão, E.A.S. Restoration of degraded Amazonian landscape: Forest recovery and agricultural restoration. Ambio 1991, 20, 248–255. [Google Scholar]
- Zahawi, R.A.; Holl, K.D. Comparing the performance of tree stakes and seedlings to restore abandoned tropical pastures. Rest. Ecol. 2009, 17, 854–864. [Google Scholar] [CrossRef]
- Zahawi, R.A.; Holl, K.D.; Cole, R.J.; Reid, J.L. Testing applied nucleation as a strategy to facilitate tropical forest recovery. J. Appl. Ecol. 2013, 50, 88–96. [Google Scholar] [CrossRef]
- Hohnwald, S.; Rischkowsky, B.; Camarão, A.P.; Schultze-Kraft, R.; Rodrigues Filho, J.A.; King, J.M. Integrating cattle into the slash-and-burn cycle on smallholdings in the Eastern Amazon: Grass-capoeira pasture versus grass-legume pasture. Agric. Ecosyst. Environ. 2006, 117, 266–276. [Google Scholar] [CrossRef]
- Hohnwald, S.; Trautwein, J.; Camarão, A.P.; Wollny, C.B.A. Relative palatability and growth performance of capoeira species as supplementary forages in the NE-Amazon. Agric. Ecosyst. Environ. 2016, 218, 107–115. [Google Scholar] [CrossRef]
- Vieira, D.L.M.; Holl, K.D.; Peneireiro, F.M. Agro-Successional Restoration as a Strategy to Facilitate Tropical Forest Recovery. Rest. Ecol. 2009, 17, 451–459. [Google Scholar] [CrossRef]
- Hohnwald, S.; Rischkowsky, B.; Schultze-Kraft, R.; Rodrigues-Filho, J.A.; Camarão, A.P. Experiences with legumes as part of a ley pasture in a low input farming system of North-Eastern Pará, Brazil. Pasturas Trop. 2005, 27, 2–12. [Google Scholar]
- Baar, R.; dos Reis Cordeiro, M.; Denich, M.; Fölster, H. Floristic inventory of secondary vegetation in agricultural systems of East-Amazonia. Biodivers. Conserv. 2004, 13, 501–528. [Google Scholar] [CrossRef]
- Rodrigues, R.R.; Lima, R.A.F.; Gandolfi, S.; Nave, A. On the restoration of high diversity forests: 30 years of experience in the Brazilian Atlantic Forest. Biol. Conserv. 2009, 142, 1242–1251. [Google Scholar] [CrossRef]
- Brown, S.; Lugo, A.E. Tropical secondary forests. J. Trop. Ecol. 1990, 6, 1–32. [Google Scholar] [CrossRef]
- Denich, M.; Vielhauer, K.; Kato, M.D.A.; Block, A.; Kato, O.R.; de Sá Abreu, T.D.; Lücke, W.; Vlek, P.L.G. Mechanized land preparation in forest-based fallow systems: The experience from Eastern Amazonia. Agric. Syst. 2004, 61, 91–106. [Google Scholar] [CrossRef]
- Kato, M.S.A.; Kato, O.R.; Denich, M.; Vlek, P.L.G. Fire-free alternatives to slash and-burn for shifting cultivation in the eastern Amazon region: The role of fertilizers. Field Crops Res. 1999, 62, 225–237. [Google Scholar] [CrossRef]
- Sommer, R.; Vlek, P.L.G.; de Sá, T.D.A.; Vielhauer, K.; Rodrigues Coelho, R.F.; Fölster, H. Nutrient balance of shifting cultivation by burning or mulching in the Eastern Amazon—Evidence for subsoil nutrient accumulation. Nutr. Cycl. Agroecosyst. 2004, 68, 257–271. [Google Scholar] [CrossRef]
- Denich, M.; Vlek, P.L.G.; de Sá, T.D.A.; Vielhauer, K.; Lücke, W. A concept for the development of fire-free fallow management in the Eastern Amazon, Brazil. Agric. Ecosyst. Environ. 2005, 110, 43–58. [Google Scholar] [CrossRef]
- Hohnwald, S. Bird Records from the Rural Landscape of Igarapé-Açu Municipality, Northeastern Pará. Boletim do Museu Paraense Emílio Goeldi. Ciências Naturais 2009, 4, 119–131. [Google Scholar]
- Hohnwald, S.; Acioli de Abréu, E.; Krummel, T.; da Veiga, J.B.; Wollny, C.B.A.; Calandrini de Azevedo, C.M.B.; Gerold, G. Degraded Pasture Expansion and Woody Enrichment Strategies for Pasture Fertility Preservation in the Bragantina Region, North-Eastern Amazon. Erdkunde 2010, 64, 17–31. [Google Scholar] [CrossRef]
- Hohnwald, S.; Rischkowsky, B.; King, J.M.; Camarão, A.P.; Rodrigues Filho, J.A.; Zeppenfeld, T. Intensive Cattle Browsing Did Not Prevent Fallow Recuperation on Smallholder Grass-Capoeira Pastures in the NE-Amazon. Agric. Syst. 2015, 89, 813–828. [Google Scholar] [CrossRef]
- Staver, C. The Role of Weeds in the Productivity of Amazonian Bush Fallow Agriculture. Exp. Agric. 1991, 27, 287–304. [Google Scholar] [CrossRef]
- Dias-Filho, M.B. Germination and emergence of Stachytarpheta cayennensis and Ipomoea asarifolia. Planta Daninha 1996, 14, 118–126. [Google Scholar] [CrossRef]
- Krummel, T.; Hohnwald, S.; Gerold, G. Biologisch degradierte Rinderweiden und ihre agrarökologischen Regradationsmöglichkeiten für Kleinbauern in Nordost-Amazonien. Innsbr. Geogr. Stud. 2011, 38, 43–56. [Google Scholar]
- Vieira, I.C.G.; Uhl, C.; Nepstad, D.C. The role of shrub Cordia multispicata Cham. as a ‘succession facilitator’ in an abandoned pasture, Paragominas, Amazônia. Vegetatio 1994, 115, 91–99. [Google Scholar]
- Holl, K.D.; Loik, M.E.; Lin, E.H.V.; Samuels, I.A. Tropical Montane Forest Restoration in Costa Rica: Overcoming Barriers to Dispersal and Establishment. Rest. Ecol. 2000, 8, 339–349. [Google Scholar] [CrossRef]
- Reid, J.L.; Holl, K.D. Arrival ≠ Survival. Rest. Ecol. 2013, 21, 153–155. [Google Scholar] [CrossRef]
- Peña-Domene, M.; Martínez-Garza, C.; Palmas-Pérez, S.; Rivas-Alonso, E.; Howe, H.F. Roles of Birds and Bats in Early Tropical-Forest Restoration. PLoS ONE 2014, 9, e104656. [Google Scholar] [CrossRef]
- Posada, J.M.; Aide, T.M.; Cavalier, J. Cattle and Weedy Shrubs as Restoration Tools of Tropical Montane Rainforest. Rest. Ecol. 2000, 8, 370–379. [Google Scholar] [CrossRef]
- Elledge, A.E.; McAlpine, C.A.; Murray, P.J.; Gordon, I.J. Modelling habitat preferences of feral pigs for rooting in lowland rainforest. Biol. Invasion 2013, 15, 1523–1535. [Google Scholar] [CrossRef]
- Nogueira-Filho, S.L.G.; Nogueira, S.S.C.; Fragoso, J.M.V. Ecological impacts of feral pigs in the Hawaiian Islands. Biodivers. Conserv. 2009, 18, 3677–3683. [Google Scholar] [CrossRef]
- Spatz, G.; Mueller-Dombois, D. Succession patterns after pig diggings in grassland communities on Mauna Loa, Hawaii. Phytocoenologia 1975, 3, 346–373. [Google Scholar]
- Censo Agropecuário 2006 and Produção da Pecuária Municipal-2014 of Igarapé-Açu; IBGE: Brasilia, Brazil, 2017. Available online: http://cod.ibge.gov.br/2358p (accessed on 15 December 2017).
- Pachêco, N.A.; Bastos, T.X. Boletim Agrometeorológico de 2007 para Igarape-Açu, PA. In Documentos 354; Embrapa Amazônia Oriental: Belém, Brazil, 2009. [Google Scholar]
- Pachêco, N.A.; Bastos, T.X. Boletim Agrometeorológico de 2008 para Igarape-Açu, PA. In Documentos 372; Embrapa Amazônia Oriental: Belém, Brazil, 2011. [Google Scholar]
- Baena, A.R.C.; Falesi, I.C.; Dutra, S. Características Físio-Químicas do Solo em Diferentes Agroecossistemas na Região Bragantina do Nordeste Paraense; Embrapa Amazônia Oriental: Belém, Brazil, 1998. [Google Scholar]
- R: A Language and Environment for Statistical Computing; R Core Team R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 17 January 2018).
- Oanda cooperation; Currency converter. 2019. Available online: https://www.oanda.com/currency/converter/ (accessed on 15 February 2019).
- Kotanen, P.M. Responses of vegetation to a changing regime of disturbance: Effects of feral pigs in a Californian coastal prairie. Ecography 1995, 18, 190–198. [Google Scholar] [CrossRef]
- Aplet, G.H.; Anderson, S.J.; Stone, C.P. Association between feral pig disturbance and the composition of some alien plant assemblages in Hawaii Volcanoes National Park. Plant Ecol. 1991, 95, 55–62. [Google Scholar] [CrossRef]
- Sharrow, S.H. Soil compaction by grazing livestock in silvopastures as evidenced by changes in soil physical properties. Agric. Syst. 2007, 71, 215–223. [Google Scholar] [CrossRef]
- Veloso, H.P. As communidades e as estações botânicas de Teresópolis, Estado do Rio de Janeiro. Boletim do Museu Nacional. Botânica. 1945, 3, 1–95. [Google Scholar]
- Albuquerque, N.I.; Guimarães, D.A.; Tavares Dias, H.L.; le Pendu, Y.; Reis Kahwage, P.; Rossetto Garcia, A. Intensive production system of collared peccary (Pecari tajacu) in Brazilian Amazon. Adv. Anim. Biosci. 2010, 1, 480–481. [Google Scholar] [CrossRef]
- Garnero, A.D.V.; Marcondes, C.R.; Albuquerque, N.I.; Araújo, R.O.; de Pendu, Y.; Guimarães, D.A. Growth curve of female collared peccaries (Pecari tajacu) raised in captivity in the Brazilian Amazon Region. Arquivo Brasileiro de Medicina Veterinária e Zootecnia 2013, 65, 961–966. [Google Scholar] [CrossRef]
- Haines-Young, R.; Potschin-Young, M. Revision of the Common International Classification for Ecosystem Services (CICES V5. 1): A Policy Brief. One Ecosyst. 2018, 3, e27108. [Google Scholar] [CrossRef]
- Díaz, S.; Pascual, U.; Stenseke, M.; Martín-López, B.; Watson, R.T.; Molnár, Z.; Hill, R.; Chan, K.M.; Baste, I.A.; Brauman, K.A. Assessing nature’s contributions to people. Science 2018, 359, 270–272. [Google Scholar] [CrossRef]
Figure 1.
Illustration of the four treatments: (a) mixed-bred cattle (upper left photo); (b) domesticated pigs (upper right; in front of the pig there is a B. verticillata weed); (c) manual soil opening (lower left) with hoes and tolerated capoeira trees in front of the day laborers; (d) control (lower right) = biologically degraded pastures, with just typical shrub vegetation of B. verticillata and M. tenella. In the right background, an example of a capoeira can be seen.
Figure 1.
Illustration of the four treatments: (a) mixed-bred cattle (upper left photo); (b) domesticated pigs (upper right; in front of the pig there is a B. verticillata weed); (c) manual soil opening (lower left) with hoes and tolerated capoeira trees in front of the day laborers; (d) control (lower right) = biologically degraded pastures, with just typical shrub vegetation of B. verticillata and M. tenella. In the right background, an example of a capoeira can be seen.

Figure 2.
Design of the experimental plot with the four treatments (trt = subplots) of which three were fenced + control (unfenced). This design was repeated on three different smallholder farms in the Bragantina region. Within the subplots, there were four rows of ten sampling plots that were investigated at three different times (n = 480).
Figure 2.
Design of the experimental plot with the four treatments (trt = subplots) of which three were fenced + control (unfenced). This design was repeated on three different smallholder farms in the Bragantina region. Within the subplots, there were four rows of ten sampling plots that were investigated at three different times (n = 480).

Figure 3.
(a–f). Ground cover changes of bare soil, shrubs, B. verticillata, M. tenella, grasses, and capoeira trees of trt1 until trt4, from time1 to time3 plus standard errors (n = 8640).
Figure 3.
(a–f). Ground cover changes of bare soil, shrubs, B. verticillata, M. tenella, grasses, and capoeira trees of trt1 until trt4, from time1 to time3 plus standard errors (n = 8640).

Figure 4.
Selected examples of effective soil opening effects of the treatment (without control): (a) trt1 (left) trampled patches inside the plots during staying overnight; (b) trt2 (centre) burrowed the soils especially along the fences; (c) trt3 (right) with spared young target trees of capoeira succession and remaining litter on the ground.
Figure 4.
Selected examples of effective soil opening effects of the treatment (without control): (a) trt1 (left) trampled patches inside the plots during staying overnight; (b) trt2 (centre) burrowed the soils especially along the fences; (c) trt3 (right) with spared young target trees of capoeira succession and remaining litter on the ground.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Lists of characteristic spontaneous native legume species (a), capoeira tree species (b), and juquira (c), ordered by its frequency on pastures in the northeastern Amazon, in its respective group.
Table 1.
Lists of characteristic spontaneous native legume species (a), capoeira tree species (b), and juquira (c), ordered by its frequency on pastures in the northeastern Amazon, in its respective group.
| Species Name | Plant (Sub) Family | Life-Form |
|---|---|---|
| a) Spontaneous Legume Species | ||
| Zornia latifolia Sm. | Papilionoideae | herb |
| Mimosa pudica L. | Mimosoideae | herb |
| Stylosanthes gracilis Kunth | Papilionoideae | shrub |
| Senna chrysocarpa (Desv.) H.S. Irwin & Barneby | Caesalpinioideae | shrub |
| Mimosa quadrivalvis L. | Mimosoideae | herb |
| Machaerium madeirense Pittier | Papilionoideae | liana |
| Machaerium froesii Rudd | Papilionoideae | liana |
| Desmodium barbatum (L.) Benth. | Papilionoideae | herb |
| Desmodium canum (J.F. Gmel.) Schinz & Thell. | Papilionoideae | herb |
| Bauhinia guianensis Aubl. | Caesalpinioideae | liana |
| b) Capoeira | ||
| Vismia guianensis (Aubl.) Choisy | Clusiaceae | tree |
| Lacistema pubescens Mart. | Lacistemataceae | tree |
| Myrcia sylvatica (G. Mey.) DC. | Myrtaceae | shrub |
| Myrcia deflexa (Poir.) DC. | Myrtaceae | shrub |
| Myrcia bracteata (Rich.) DC. | Myrtaceae | shrub |
| Banara guianensis Aubl. | Connaraceae | tree |
| Lecythis lurida (Miers) S.A. Mori | Lecythidaceae | tree |
| Abarema cochleata (Willd.) Barneby & J.W. Grimes | Mimosoideae | tree |
| c) Juquira | ||
| Borreria verticillata (L.) G. Mey. | Rubiaceae | shrub |
| Myrciaria tenella (DC.) O. Berg | Myrtaceae | shrub |
| Borreria latifolia (Aubl.) K. Schum. | Rubiaceae | shrub |
| Stachytarpheta cayennensis (Rich.) M. Vahl | Verbenaceae | shrub |
| Paspalum maritimum Trin. | Poaceae | herb |
| Paspalum conjugatum P.J. Bergius | Poaceae | herb |
| Panicum pilosum Sw. | Poaceae | herb |
| Scleria pterota C. Presl | Cyperaceae | herb |
| Rourea ligulata Baker | Connaraceae | liana |
| Rolandra argentea Rottb. | Asteraceae | shrub |
| Rollinia exsucca (DC. ex Dunal) A. DC. | Annonaceae | tree |
| Hyptis atrorubens Poit. | Lamiaceae | herb |
| Imperata brasiliensis Trin. | Poaceae | herb |
| Emilia sonchifolia (L.) DC. | Asteraceae | herb |
| Ipomoea asarifolia (Desr.) Roem. & Schult. | Verbenaceae | liana |
| Borreria suaveolens G. Mey. | Rubiaceae | shrub |
| Andropogon bicornis L. | Poaceae | herb |
| Andropogon leucostachyus Kunth | Poaceae | herb |
Table 2.
Number of capoeira tree individuals/6.25 m2, mean tree heights of the five highest trees at time1 and time2, and mean number of capoeira saplings (just time3) plus standard deviations (n = 3929). a–c Values in the same column with the different superscript letters are significantly different from each other at p < 0.05. 1–2 Values in the same row with the different superscript numbers are significantly different from each other at p < 0.05 (for capoeira tree number, mean tree heights, and capoeira saplings number, calculated, respectively).
Table 2.
Number of capoeira tree individuals/6.25 m2, mean tree heights of the five highest trees at time1 and time2, and mean number of capoeira saplings (just time3) plus standard deviations (n = 3929). a–c Values in the same column with the different superscript letters are significantly different from each other at p < 0.05. 1–2 Values in the same row with the different superscript numbers are significantly different from each other at p < 0.05 (for capoeira tree number, mean tree heights, and capoeira saplings number, calculated, respectively).
| Capoeira Trees time1 [#] | Capoeira Trees time2 [#] | Mean Tree Heights time1 [m] | Mean Tree Heights time2 [m] | Capoeira Saplings time3 [#] | |
|---|---|---|---|---|---|
| n = 1440 | n = 1129 | n = 960 | |||
| trt1 | 5.84 (0.47) ab1 | 6.13 (0.54) a1 | 1.0 a1 (0.6) | 1.1 a1 (0.4) | 2.56 (2.8) a |
| trt2 | 7.07 (0.45) a1 | 6.42 (0.41) a1 | 1.3 b1 (0.7) | 1.3 b1 (0.6) | 2.64 (2.8) a |
| trt3 | 5.52 (0.40) b1 | 4.55 (0.45) b1 | 0.9 a1 (0.7) | 1.3 b2 (0.4) | 2.84 (3.0) a |
| trt4 | 6.52 (0.41) ab1 | 3.77 (0.29) b2 | 1.1 a1 (0.6) | 1.5 c2 (0.5) | 1.33 (1.8) b |
Table 3.
Ground cover of spontaneously growing herbaceous legumes at time1 and time2 plus standard deviations (n = 3929). a–c Values in the same column with the different superscript letters are significantly different from each other at p < 0.05, 1–2 Values in the same row with different superscript numbers are significantly different from each other at p < 0.05.
Table 3.
Ground cover of spontaneously growing herbaceous legumes at time1 and time2 plus standard deviations (n = 3929). a–c Values in the same column with the different superscript letters are significantly different from each other at p < 0.05, 1–2 Values in the same row with different superscript numbers are significantly different from each other at p < 0.05.
| Legume Cover time1 [%] | Legume Cover time2 [%] | |
|---|---|---|
| n = 1360 | ||
| trt1 | 6.9 a1 (11.0) | 14.4 a2 (19.4) |
| trt2 | 15.6 b1 (20.6) | 15.4 a1 (18.5) |
| trt3 | 20.3 bc1 (25.1) | 29.2 b2 (27.0) |
| trt4 | 24.0 c1 (28.4) | 14.4 a2 (19.7) |
Table 4.
Soil bulk densities [MPa = 100 N/cm2] at time1 and time2 in six different soil depths with standard errors in brackets (n = 893). a–d values within the same depths (time1 + time2 together) with different superscripts are significantly different at p < 0.05 (experimental unit n = 3).
Table 4.
Soil bulk densities [MPa = 100 N/cm2] at time1 and time2 in six different soil depths with standard errors in brackets (n = 893). a–d values within the same depths (time1 + time2 together) with different superscripts are significantly different at p < 0.05 (experimental unit n = 3).
| time1 | ||||
| Depths | trt1 | trt2 | trt3 | trt4 |
| 2.5 cm | 1.88 (0.13) a | 1.95 (0.13) a | 1.71 (0.14) a | 2.05 (0.14) a |
| 5 cm | 2.07 (0.13) ab | 2.21 (0.13) ab | 1.82 (0.15) a | 2.25 (0.12) b |
| 10 cm | 2.21 (0.13) ab | 2.37 (0.11) a | 1.90 (0.16) b | 2.43 (0.10) a |
| 15 cm | 2.18 (0.13) ab | 2.32 (0.16) ab | 1.90 (0.16) a | 2.40 (0.11) b |
| 20 cm | 2.12 (0.14) a | 2.24 (0.18) a | 1.68 (0.12) b | 1.98 (0.14) ab |
| 40 cm | 1.56 (0.09) a | 1.34 (0.17) a | 1.38 (0.11) a | 1.40 (0.05) a |
| time2 | ||||
| Depths | trt1 | trt2 | trt3 | trt4 |
| 2.5 cm | 2.95 (0.18) b | 3.06 (0.19) bc | 2.63 (0.19) bc | 2.44 (0.15) ac |
| 5 cm | 3.04 (0.15) cd | 3.51 (0.19) bcd | 3.43 (0.20) c | 2.84 (0.17) bd |
| 10 cm | 3.06 (0.14) c | 3.77 (0.15) c | 4.20 (0.18) c | 4.01 (0.19) c |
| 15 cm | 3.26 (0.15) c | 3.86 (0.17) c | 4.05 (0.25) c | 4.67 (0.14) c |
| 20 cm | 3.34 (0.20) c | 3.61 (0.19) c | 3.45 (0.27) c | 3.42 (0.29) c |
| 40 cm | 2.51 (0.16) b | 3.26 (0.22) ab | 4,29 (0.19) b | 4.21 (0.21) b |
Table 5.
Pig weights [in kg] at time1 and time2 with standard deviations in brackets.
| Number of Pigs | Mean Pig Weights [kg/pig] | Total Pig Weight before [kg/0.5 ha] | Total Pig Weight after [kg/0.5 ha] | Total Weight Gain [kg; %] | Economic Gain from Pigs [US$/ha] | |
|---|---|---|---|---|---|---|
| farm1 | 14 | 20.1 | 281 (20.0) | 572 (25.0) | 291; +104% | 727.50 |
| farm2 | 11 | 24.0 | 264 (17.3) | 467 (18.7) | 203; +77% | 507.50 |
| farm3 | 10 | 29.8 | 298 (15.5) | 624 (21.6) | 326; +109% | 815.00 |
| mean | 11.7 | 24.6 | 281.0 | 554.3 | 273.3 | 683.33 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hohnwald, S.; Kato, O.R.; Walentowski, H. Accelerating Capoeira Regeneration on Degraded Pastures in the Northeastern Amazon by the Use of Pigs or Cattle. Sustainability 2019, 11, 1729. https://doi.org/10.3390/su11061729
AMA Style
Hohnwald S, Kato OR, Walentowski H. Accelerating Capoeira Regeneration on Degraded Pastures in the Northeastern Amazon by the Use of Pigs or Cattle. Sustainability. 2019; 11(6):1729. https://doi.org/10.3390/su11061729
Chicago/Turabian StyleHohnwald, Stefan, Osvaldo Ryohei Kato, and Helge Walentowski. 2019. "Accelerating Capoeira Regeneration on Degraded Pastures in the Northeastern Amazon by the Use of Pigs or Cattle" Sustainability 11, no. 6: 1729. https://doi.org/10.3390/su11061729
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.