Screening of Rice Endophytic Biofertilizers with Fungicide Tolerance and Plant Growth-Promoting Characteristics


1
Department of Soil and Environmental Sciences, National Chung-Hsing University, Taichung 40227, Taiwan
2
Innovation and Development Center of Sustainable Agriculture (IDCSA), National Chung-Hsing University, Taichung 40227, Taiwan
3
Department of Agricultural Chemistry, National Taiwan University, Taipei 10617, Taiwan
4
Department of Civil and Ecological Engineering, I-Shou University, Kaohsiung 84001, Taiwan
5
International Bachelor Program of Agribusiness, National Chung-Hsing University, Taichung 40227, Taiwan
*
Author to whom correspondence should be addressed.
Sustainability 2019, 11(4), 1133; https://doi.org/10.3390/su11041133
Submission received: 17 January 2019
/
Revised: 18 February 2019
/
Accepted: 19 February 2019
/
Published: 21 February 2019
(This article belongs to the Special Issue Sustainability of Microbial Ecosystems: Soil and Aquatic Microbial Diversity)
Abstract
:The application of pesticides is unavoidable in conventional agricultural practice. To develop effective biofertilizers, bacterial strains with both pesticide tolerance and plant growth-promoting (PGP) traits were isolated for further testing. Seedlings of rice (Oryza sativa) were planted in soil with 1, 5, or 10 times the recommended rates of the fungicides etridiazole, metalaxyl, and tricyclazole. Endophytic bacteria were isolated from roots of rice seedlings. The bacterial 16S rDNA sequences and related PGP characteristics including potential nitrogen fixation, phosphorus-solubilizing and indole acetic acid (IAA) production ability were further examined. In all, 17 different strains were obtained from rice seedling roots; five strains with both nitrogen fixation potential and IAA production ability included Rhizobium larrymoorei E2, Bacillus aryabhattai E7, Bacillus aryabhattai MN1, Pseudomonas granadensis T6, and Bacillus fortis T9. Except for T9, all strains could tolerate two or more fungicides. We inoculated rice roots with the endophytic bacteria and all conferred rice growth-promoting ability. Bacillus aryabhattai MN1 was further tested and showed high tryptophan dose-dependent IAA production ability, tolerance towards etridiazole and metalaxyl application and should be considered a potential bacterial biofertilizer.
1. Introduction
Rice is the largest crop cultivated in Taiwan and is a major source of food in Asia. To achieve high yield, considerable progress has been made in its cultivation, including breeding, nursery cultivation, fertilizer management and pest control, for one of the most representative crops in the world [1].
Bacterial biofertilizers are plant growth-promoting rhizobacteria (PGPR) applied in the field to promote crop growth and reduce the need for synthetic fertilizers. PGPR promote plant growth in many ways, including nitrogen fixation, phosphorus dissolution, secretion of chelating substances such as siderophores; cyanide and extracellular polysaccharide; and plant hormones such as indole acetic acid (IAA) [2].
Endophytic bacteria are defined as bacteria colonizing the internal tissues of plants (apoplasm or symplasm) without causing symptomatic infections or having negative effects on their host [3]. As compared with many rhizobacteria, endophytes have generally greater beneficial effects [4] and are less likely to encounter an extreme change of the soil environment because of the insulation of plants. Thus, to achieve better yield and protection of rice, the use of endophytic rather than rhizobacter biofertilizers, is preferred.
The possibility of inoculating rice endophytic bacteria strains has been studied. Diazotrophic endophyte Azoarcus sp. BH72 is one model species among many that was found capable of colonizing the interior of rice roots [5]. Rice endophyte Burkholderia vietnamensis was inoculated in pots and in the field to raise rice grain production significantly [6]. Stenotrophomonas maltophilia and Ochrobactrum sp. were isolated from rice seeds, and from the result of PCR-DGGE analysis, soil types alter rice endophytic bacterial diversity: the endophytes Pseudomonas oryzihabitans and Rhizobium radiobacter dominated in neutral pH soil, Enterobacter-like and Dyella ginsengisoli dominated in acid soil, and Stenotrophomonas maltophilia could be found in both soil types [7]. The production of 1-aminocyclopropane-1-caboxylate (ACC) deaminase was found to be a useful trait for elongation and endophytic colonization of rice roots under constant flooded conditions [8]. Therfore, endophytic bacteria could be used as biofertilizers in promoting rice growth and production.
However, to develop effective bacterial biofertilizers, PGP traits and also pesticide tolerance should be confirmed before their application because applying pesticides is inevitable in most agriculture practices [9]. The fungicides etridiazole, metalaxyl, and tricyclazole are recommended for rice leaf blast and rice blight in rice seedling boxes in Taiwan. Etridiazole [5-ethoxy-3(trichloromethyl)-1,2,4-thiadiazole] causes the hydrolysis of cell membrane phospholipids into free fatty acids and lysophosphatides, leading to the lysis of membranes in fungi [10,11]. Metalaxyl [methyl 2-(N-(2-methoxyacetyl)-2,6-dimethylanilino) propanoate] inhibits RNA and protein synthesis in many members of the Peronosporales [12]. Tricyclazole [5-methyl-1,2,4-triazolo[3,4-b] benzothiazole] is a systemic fungicide used to control Pyricularia oryzae (rice blast) in transplanted and direct-seeded rice [13]. Because of its systemic activity, it also reduces damage to roots as seedlings against Pythium spp. seed decay [14].
Fungicide application may have adverse effects on PGP traits in rhizobacteria. Although phosphate-solubilizing Pseudomonas putida isolated from mustard (Brassica compestris) rhizosphere could survive with high concentrations of fungicides hexaconazole, ketazin, metalaxyl, and tebuconazole, PGP attributes such as phosphate-solubilizing and IAA secretion were significantly and dose-dependently inhibited by fungicides at recommended rates [9]. Application of organophosphates on the PGPR Methylobacterium spp. and Bacillus spp., markedly reduced the microbial population, especially Methylobacterium spp. which was completely extinct in soil treated with chlorpyrifos, phorate and dimethoate [15]. The tolerance of 20 bacterial strains isolated from rhizosphere of okra plants with high doses of bifenthrin applied, demonstrated that the growth of most of the strains decreased with increasing pesticide dose [16]. It was reported that etridiazole and metalaxyl adversely affected ammonifying or nitrifying bacteria in soils [11]; therefore, the multiple or alternatively apply of different fungicides such as etridiazole, metalaxyl, and tricyclazole may also impede the functions of the non-target PGPR. The hypothesis of this study was that applying of biofertilizers with pesticide-tolerant ability would maintain their functions and would be capable of promoting the growth of the crops under the pesticides applied.
According to Taiwan’s plant protection manual [17], the recommended rates for etridiazole, metalaxyl, and tricyclazole are 6.7, 9.3, and 160.3 mg/kg, respectively in a 60 × 30 × 4 cm rice seedling box. From previous research, the EC50 on Pythium sp. isolated from carrot were 11.1 and 133.3 mg/L for metalaxyl and etridiazole, respectively [18]. The EC50 for tricyclazole on rice blast Magnaporthe oryzae was 100.41 mg/L [19]. Tricyclazole, at higher levels (20–40 mg/kg), reduced the growth and sporulation of the cadmium-tolerant fungi Exophiala pisciphil, but the toxic effects were not observed at a lower concentration, 2.5 mg/kg [20].
From previous studies, it is important to examine the impact of pesticides during the process of developing biofertilizers in order to make sure the functions of the biofertilizers in the field. This study aims at screening potential endophytic biofertilizers with fungicides tolerance and plant growth-promoting characteristics.
2. Materials and Methods
2.1. Isolation of the Rice Endophytes
Following the suggestions from Taiwan’s Plant Protection Manual [17], we applied 1, 5, and 10 times the recommended rates of tricyclazole, metalaxyl and etridiazole to soils in plastic trays where rice (Oryza sativa, cultivar Tainan 11) was planted. We also followed the manual’s suggestions to apply the fungicides at different periods. Tricyclazole and etridiazole were applied before rice planting, and metalaxyl was applied a week after rice germination. Rice grains were surface-sterilized prior to the test by soaking them in 60 °C sterilized H2O for 10 min, then seeds were soaked in sterilized H2O for germination. The soils were from paddy fields in Hualien, with pH 6.7 and texture silt loam. After incubation in the plant growth chamber for 2 weeks at 25 °C, humidity 70%, with 12 h light: 12 h dark photoperiod, rice endophytes were isolated.
Rice plants were uprooted from the soil. The shoots and the roots were separated. Roots were soaked in sterilized H2O to remove soil particles, then in 70% ethanol solution for 30 s, rinsed with sterilized H2O again and the root surface was cleaned with 3% sodium hypochlorite solution for 3 min. Roots were washed thoroughly by using sterilized H2O several times. The final portion of the sterilized H2O washed through the root surface was collected. The water was spread onto a sterilized nutrient broth agar plate to ensure total sterilization of the root surface. A mortar was used to grind roots. Root debris was diluted by using sterilized H2O and the suspension was spread onto a sterilized nutrient broth agar plate.
Plates were incubated under 30 °C for several days after colonies were formed. Colonies with different morphology were isolated and incubated in a new sterilized nutrient broth agar plate, and the purity of the colony was confirmed by using the streak plates method twice.
2.2. Identification of Bacteria
DNA was extracted from isolated colonies by using the Exgene DNA Purification Kit (GeneAll Biotechnology). DNA fragments were amplified by using 16S rDNA universal primer 27 F and 1492 R [21] with a polymerase chain reaction (PCR) machine (Biometra T3000). The PCR reaction mixture contained 2x Taq PCR Master Mix (Ginomics BioScience and Technology) and 0.4 μM of the primers. The 16S rDNA was amplified with 95 °C denaturing for 5 min, followed by 35 cycles of 94 °C for 0.25 min, 60 °C for 1.25 min, 72 °C for 1.5 min, and with a final extension at 72 °C for 5 min. The DNA sequencing was performed with the ABI PRISM 3700 DNA Analyzer (Perkin Elmer Applied Biosystems) by Seeing Biosciences (Taiwan).
2.3. Phylogenic Tree
The DNA sequence was searched in the EzTaxon database on 27 July 2017. The phylogenetic tree was constructed by using Mega 7.0 software with the neighbor-joining method and maximum composite likelihood method confirmed by 1000 times of bootstrap analysis. DNA sequences containing gaps and missing data were eliminated [22,23,24,25].
2.4. Indole Acetic Acid (IAA) Production
IAA production was quantified as described as described by Gordon and Weber (1951) [26] with modification following Brick et al. (1991) [27]. In brief, we inoculated the bacteria strain into 100 mL Luria Bertani (LB) broth (g/L: tryptone 10; yeast extract 5; NaCl 10 and pH 7.5) with 100 μg/mL tryptophan amendment. Three replicates of the culture were agitated under 30 °C for 24 h. Then, 5 mL of the culture was centrifuged at 9000× g for 15 min. An amount of 1 mL supernatant was mixed with 1 mL Salkowsky reagent (2% 0.5 M FeCl3 in 35% perchloric acid) and incubated in the dark for 1 hr. The IAA concentration in the supernatant was determined at 540 nm by using a spectrophotometer (Thermo ScientificTM GENESYS 10S) against a standard curve.
2.5. Potential for Nitrogen-Fixing and Phosphate-Solubilizing Ability
The culture solution was spread onto a sterilized NFB medium plate [28,29,30] or a Pikovskaya plate [31]. The bacteria were considered potential nitrogen-fixing bacteria if the color of the plate turned from green to blue after 2 weeks of incubation on an NFB medium plate. The bacteria were considered potential phosphate-solubilizing bacteria if a clear zone around bacterial colony was formed after 7 days of cultivation on a Pikovskaya plate.
2.6. Fungicides on Bacteria Growth
Fungicides tricyclazole, metalaxyl and etridiazole were applied at recommended rates following Methods Section 2.1 into NB medium inoculated with the bacteria showing PGP characteristics. The absorbance was measured at OD600 to determine whether bacterial growth was affected by amendment with the fungicides.
2.7. Endophytic Colonization and the Effect on Rice Growth
We chose bacteria with both PGP and fungicide tolerance for the following test. Bacteria were inoculated in NB medium solution after colonies exceeded 106 colony forming units/mL. We centrifuged the culture solution and suspended the pellet by using an equal volume of 0.85% NaCl solution to become a mixture. Two mL of the mixture containing different bacteria pellets were then inoculated separately onto 15 rice seedlings (root length <1 cm) in a petri dish with a filter paper. Three replicates of the petri dishes were incubated in a plant growth chamber (conditions resembled those in Methods Section 2.1). The shoot and root length were measured after 4 days of incubation. Prior to the germination, the rice grains were surface-sterilized following the procedure in Methods Section 2.1 before the experiment was conducted.
We tested whether the bacteria were successfully inoculated in the rice roots by also following the isolation procedure from Methods Section 2.1. All strains were successfully isolated from rice roots after inoculation (data not shown).
2.8. IAA Production in MN1 Culture Medium with Different Tryptophan Concentration
We amended LB broth with 0, 100, 500, 1000 mg/L tryptophan and inoculated MN1 in the culture medium in three replicates. After 1, 5, and 7 days of cultivation, the supernatant was filtered by using a 0.22-μm membrane. The amount of IAA was quantified following Methods Section 2.9.
2.9. The Effect of Fungicides on IAA Production by MN1
We applied the recommended rates of metalaxyl and etridiazole to Bacillus aryabhattai MN1-inoculated LB broth containing 500 mg/L tryptophan. After 5 days, we determined IAA production by using high-performance liquid chromatography (HPLC) [32] instead of spectrophotometry due to the higher accuracy provided by the HPLC. Chromatographic separation was performed at ambient temperature on a C-18 column (5 μm, 250-4, Merck) using a gradient elution. Eluent A consisted of 0.1: 99.9% (v/v) acetic acid: H2O, pH 3.8 (pH was adjusted by the addition of 1 mol/L KOH) and eluent B consisted of 80%: 20% (v/v) acetonitrile: H2O, the HPLC gradient condition is listed in Table 1. UV wavelength was set to 249 nm. The absorbance wavelength of tricyclazole was close to that for IAA, so we did not test the effect of tricyclazole on IAA production in this experiment.
2.10. Statistical Analysis
Data were analyzed by one-way analysis of variance (ANOVA) with SAS v9.4 (SAS Institute Inc., Cary, NC, USA). Post-hoc analysis involved Fisher’s least significant difference (LSD) test for significant difference between treatments. p < 0.05 was considered statistically significant.
3. Results and Discussion
3.1. Endophytes’ Isolation with Application of Fungicides
We isolated bacteria with different morphology from the rice endophytes treated with 1–10 times the recommended rates of tricyclazole, metalaxyl and etridiazole applied in soil: 13 bacterial strains under tricyclazole, 10 with metalaxyl, and 11 under etridiazole were cultured and isolated. We eliminated high-homologous-sequence strains after sequencing. Five endophytic bacteria strains resulted from triclazole treatments, three from metalaxyl treatments and nine from etridiazole treatments (Table 2).
The phylogenetic tree of the fungicide-tolerant bacterial strains is shown in Figure 1. Three Mycobacerium spp.—Etri N1, Etri 09, and Etri 10—had 100% consensus rates, and three Pseudomonas spp.—Tric N1, Tric N3 and Tric 06—also had 100% consensus rate, indicating the robustness of the tree.
From Figure 1, the isolates were more abundant and diverse under 10 times the etridiazole recommended dose (67 mg/Kg) as compared with the other treatments. The isolates included Gram-positive bacteria Mycobacterium aromaticivorans, M. sphagni, M. vanbaalenii, and Bacillus aryabhattai, and Gram-negative Brevundimonas vesicularis, Flavobacterium ginsengiterrae, Pseudacidovorax intermedius, Rhizobium larrymoorei, and Sphingomonas aquatilis. The phenomenon was probably due to the low absorption/distribution rate by the plants or the lower toxicity of etridiazole towards soil biomes. Although etridiazole had been reported to reduce the nitrification rate of ammonium-oxidizing bacteria in soil [33], the nitrogen-fixing bacteria were isolated on high-dose applications in this study (Table 2).
All strains from metalaxyl treatments were Gram-positive Bacillus spp. including B. sporothermodurans, B. fortis, and B. aryabhattai. All three species were reported to be able to survive under adverse environment by producing resistant endospores [34,35,36].
Strains isolated from tricyclazole treatment were Bacillus spp. and Pseudomonas spp., including B. fortis and B. marisflavi, P. baetica, P. plecoglossicida, and P. granadensis. Among them, P. plecoglossicida was reported to be equipped with bioremediation potential and plant-promoting ability [37,38], and P. granadensis was reported to be isolated from pharmaceutical applied soil [39]. From previous research, several strains from the Bacillus and Pseudomonas genera were also found to use insecticide phorate and thiamethoxam as sole carbon source at the same time [40].
We tested the PGP characteristics of these bacteria strains (Table 2). Most of the above-mentioned bacteria strains had one or more PGP characteristics. Fourteen of 17 strains were potential nitrogen-fixing species, including Bacillus spp., Pseudomonas spp., Pseudacidovorax spp., Rhizobium spp., Sphingomonas spp. and Mycobacterium spp. Among them, Pseudomonas intermedius, Brevundimonas vesicularis, Bacillus aryabhattaihas, Bacillus marisflavi, and Pseudomonas baetica were previously reported to have nitrogen-fixing ability [41,42,43,44].
Six strains were found with phosphate-solubilizing ability, including Sphingomonas spp., Mycobacterium spp., and Pseudomonas spp. However, the phosphate-solubilizing clear zone was not large enough for SI index calculation.
Five strains had IAA secretion ability, including Rhizobium spp., Bacillus spp. and Pseudomonas spp. The IAA secretion rate was higher for B. aryabhattai MN1 and B. aryabhattai E7 than others and were 8.7 and 7.9 mg/L, respectively. B. aryabhattai, isolated from Crocus sativus rhizosphere, was previously found to produce IAA [45].
Previous research screening for endophytic bacteria with in vitro IAA production potential showed that they could provide a reliable base for selection of effective PGP bacteria [46]. Therefore, we chose R. larrymoorei E2, B. aryabhattai E7, B. aryabhattai MN1, P. granadensis T6 and B. fortis T9, with both nitrogen-fixing and IAA secretion ability for the fungicide tolerance test.
3.2. Fungicide Tolerance and Rice Inoculation Test
Table 3 shows the inhibition effect of fungicides on the growth of rice endophytes E2, E7, MN1,T6, T9 and the PGP effect of the endophytes on rice. Under 160 mg/L tricyclazole, only growth of MN1 was not inhibited. Under 9 mg/L metalaxyl, the growth of E2, E7, MN1, T6 and T9 was not significantly inhibited, whereas under 7 mg/L etridiazole, only the growth of T9 was significantly inhibited. The 3 bacterial strains E2, E7, and T6 were able to tolerate etridiazole and metalaxyl fungicides, whereas only MN1 was able to tolerate all tested fungicides. As a result, we tested the rice inoculation effect of E2, E7, MN1, and T6.
We combined two batches of experiments of inoculating endophytes on the growth of rice (Table 3). The average root length of E7 and MN1 was 26.8 and 31.2 mm, both significantly longer than that of the control (15.9 mm). The average shoot length of E7 and MN1 was 9.2 and 10.2 mm, although higher shoot lengths were observed, it was not significantly different from the control (7.9 mm) under 95% confidence level.
The average root lengths with E2 and T6 treatments were 40.3 and 39.2 mm, both significantly longer than the control (20.2 mm). The average shoot lengths with E2 and T6 treatments were 16.2 and 14.4 mm, also both significantly longer than the control (9.9 mm). We isolated the endophytes from the rice seedling and verified the successful inoculation (data not shown), thus assuming that the rice growth promotion was contributed by the inoculation of the endophytes E2, E7, MN1 and T6.
The above tests demonstrated that strain MN1 showed the highest IAA production rates as well as multiple plant growth promoting and fungicide tolerance ability; we therefore examine the growth curve, the tryptophan dose-dependent IAA production rates, and the fungicides’ amendment effect on IAA production of MN1 in latter experiments.
3.3. The Growth Curve of MN1 under Fungicides Application
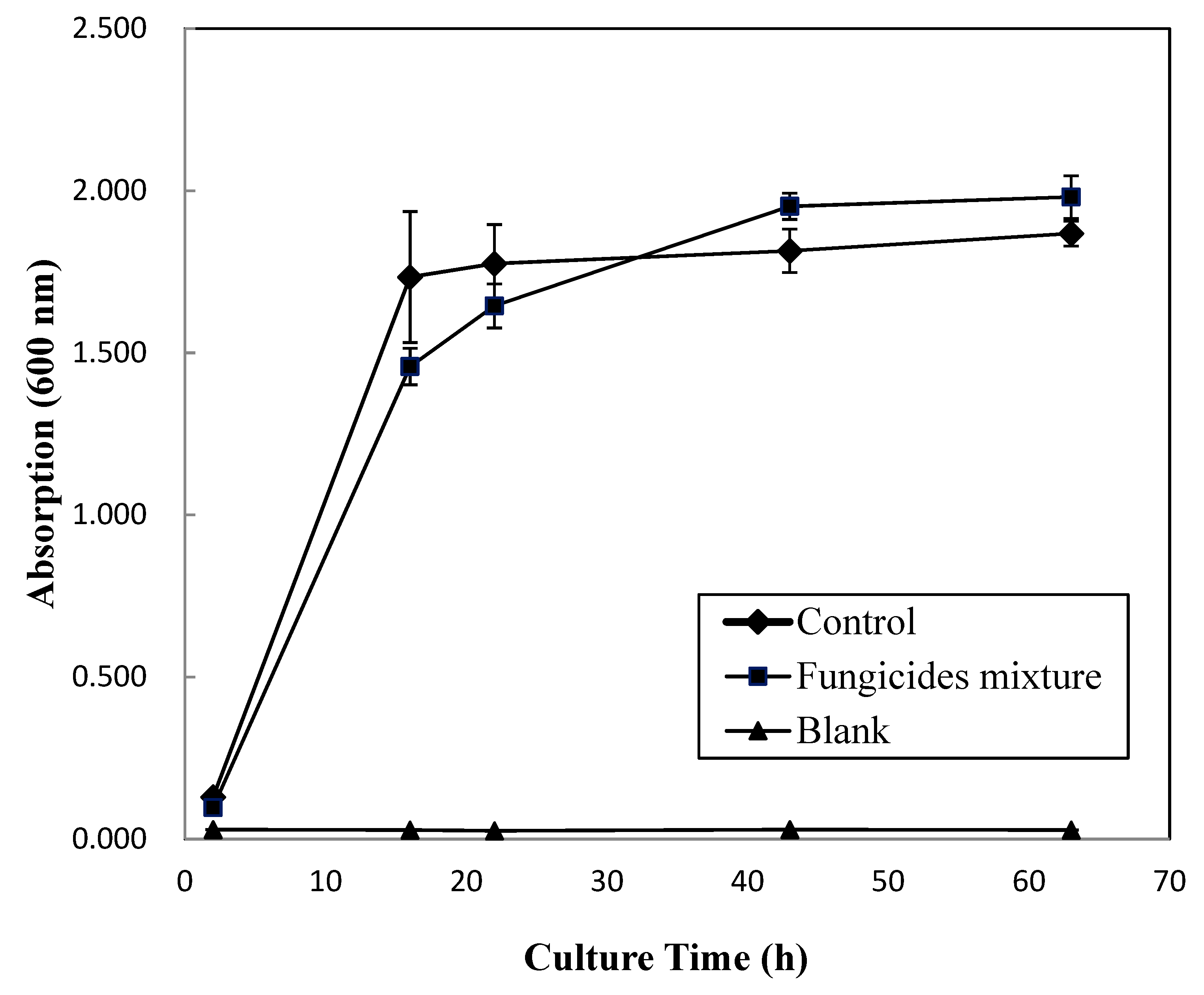
The MN1 culture medium was amended with a mixture of the recommended rates of the three fungicides. The fungicides’ mixture only slightly retarded the growth of MN1 within 35 h of incubation (Figure 2); however, after 35 h, the population of MN1 was slightly augmented with the amendment. From previous studies, the application of metalaxyl (both soil and foliar applied) and tricyclazole (foliar applied) at field-recommended rates did not affect the structure and diversity of bacteria and fungi communities [47,48]. Etridiazole at 100 mg/L application (approximately 14 times the recommended rate) was apparently fatal to ammonium-oxidizing bacteria [33]. In our study, the growth of MN1 was slightly augmented with the fungicides mixture amendment, making it among one of the possible bacterial members capable of utilizing or co-metabolizing the fungicides. From a previous study, one B. arybhattai strain isolated from rice rhizospheres was found to be equipped with arsenic absorption and accumulation abilities [49]. However, further analysis still needed to verify its fungicide utilizing ability.
3.4. Tryptophan Dose-Dependent IAA Production Rates of MN1
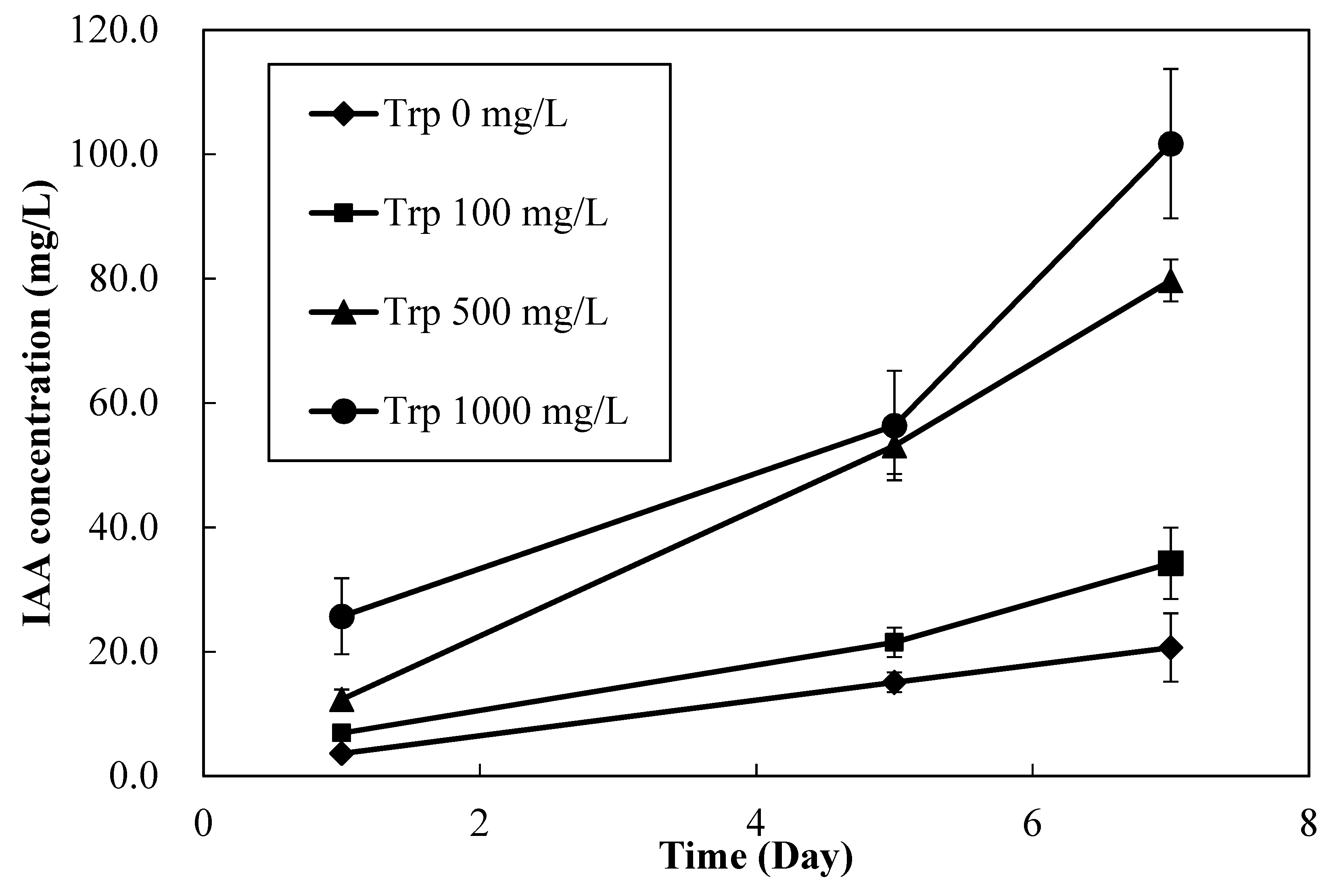
Figure 3 shows the IAA-production ability of MN1 with different concentrations of L-tryptophan amendment. After seven days of inoculation, the IAA concentrations in the supernatant with 100, 500, and 1000 mg/L tryptophan treatments were 34.2 ± 5.7, 79.7 ± 3.4, and 101.7 ± 12.0 mg/L, respectively. The IAA production ability of MN1 increased with increasing amount of tryptophan amendment. Since the concentrations of tryptophan did not significantly alter the growth of MN1 (data not shown), we assumed that the IAA production rate of MN1 species increased with increasing concentration of tryptophan. Similar findings were reported previously [50]: Higher concentrations of tryptophan amendment were previously reported to increase the IAA production rates of Azotobacter spp., fluorescent Pseudomonas spp., and Enterobacter spp. I-3. However, if considering the IAA production rate from unit tryptophan amended at day seven, it was 18.6% for 500 mg/L tryptophan applied and 11.9% for 1000 mg/L tryptophan.
No tryptophan amendment of MN1 produced 3.6 ± 0.6, 15.1 ± 1.6, and 20.7 ± 5.5 mg/L IAA at days 1, 5 and 7, MN1, respectively (Figure 3), showing the reasonable assumption that NB broth should contain trace amounts of tryptophan, although relevant tests should be conducted.
The corresponding IAA production by MN1 also showed that MN1 may benefit different plant species by various concentrations of tryptophan amendment, because for each specific plant or even each plant cultivar, there should be a suitable PGP exogenous IAA concentration produced by MN1 with different concentrations of tryptophan applied for future biofertilizer application.
3.5. Effect of Fungicides on IAA Production of MN1
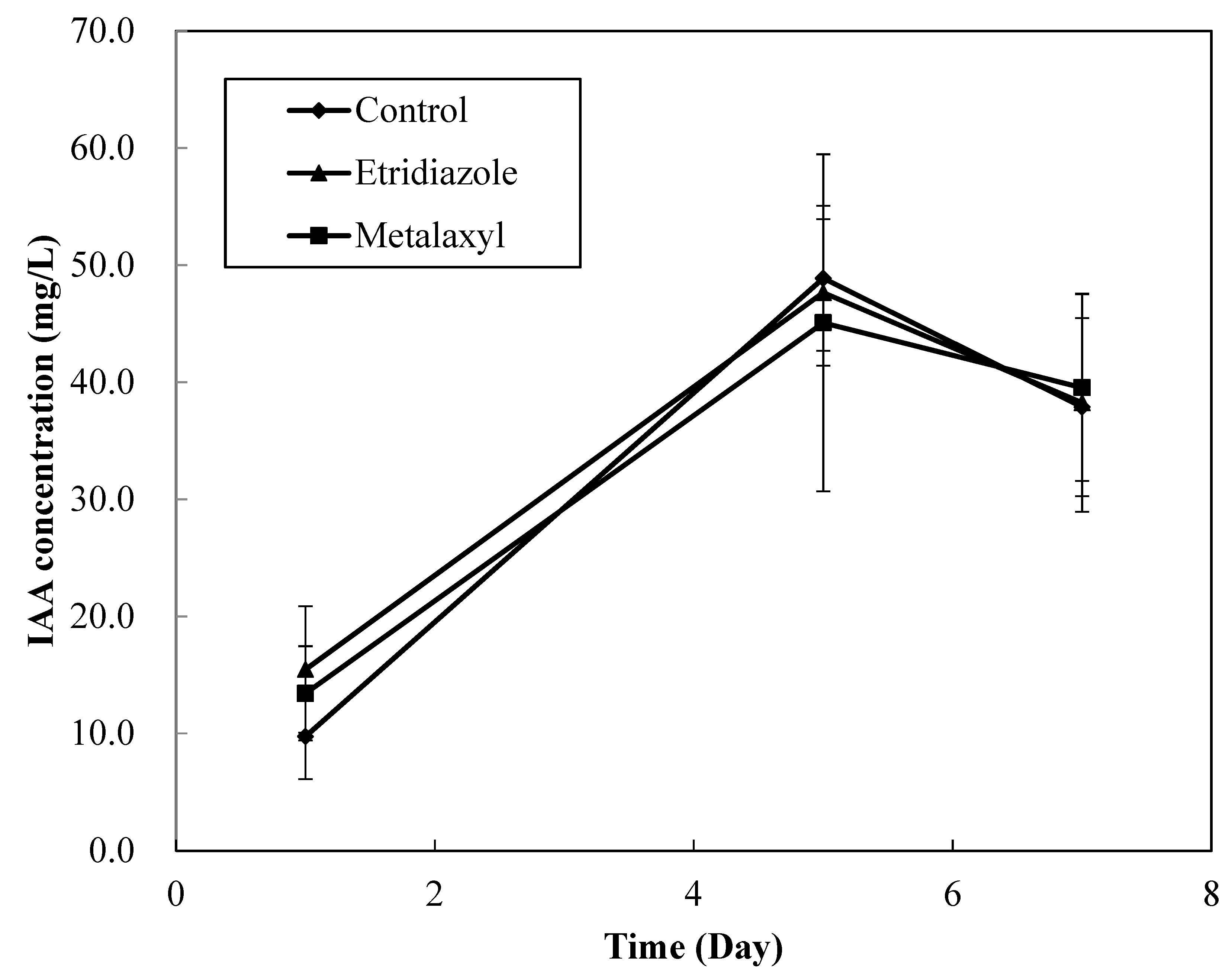
We inoculated MN1 in LB culture medium containing 500 mg/L tryptophan and etridiazole and metalaxyl (Figure 4). We examined IAA production only with 500 mg/L tryptophan because this conferred the highest growth rate per unit of tryptophan applied. Data for tricyclazole amended are also not shown because under the current HPLC analytic condition, the peaks for IAA and tricyclazole overlapped.
The IAA production of MN1 with control, etridiazole and metalaxyl treatments after 1 day of cultivation were 9.8, 15.5 and 13.4 mg/L, respectively. Higher IAA production rates were observed at day 5, which were 48.9, 47.7, and 45.1 mg/L, respectively. Although we observed varied IAA production with both fungicide treatments, we found no significant differences at the 95% confidence level. Thus, we assumed that the fungicides applied had no significant effect on IAA production by MN1.
From previous research, a specific B. arybhattai strain isolated from grapevine rhizospheres was considered a potential biofertilizer because it showed various PGP abilities such as nitrogen fixation and the production of IAA, siderophores, 1-aminocyclopropane-1-carboxylate (ACC) deaminase, chitinase, and protease [51]. Another B. arybhattai strain SanPS1 also isolated from soil showed chlorpyrifos and malathion degradation ability [52]. Being among one of the potential bioremediation members as well, it was reported that one B. arybhattai strain isolated from rice rhizospheres was equipped with arsenic absorption and accumulation abilities [49]. The above findings and our research together show that B. arybhattai has diverse functions in various applications.
4. Conclusions
From the above experiments, the bacterial strain Bacillus arybhattai MN1 proved to have nitrogen-fixing and IAA-production abilities. With application of the fungicides metalaxyl and etridiazole, the IAA production ability was not adversely affected. Previous results showed that B. arybhattai has diverse functions in various applications, but from our findings, B. arybhattai MN1was the first rice endophyte reported to possess PGP characteristics under multiple fungicides applied, and can promote the growth of rice seedlings. As compared with the isolates from the plant rhizosphere, bacterial endophytes have an advantage because living within a plant’s tissues represents an opportunity to always be in contact with plant cells and, therefore, more readily exerts a direct beneficial effect [53]. Thus, in accordance with our hypothesis that applying biofertilizers with pesticide-tolerant ability would maintain their functions and would be capable of promoting the growth of the crops under the pesticides applied, B. arybhattai MN1 should be a potential endophytic biofertilizer with fungicide tolerance ability and suitable for application in conventional rice seedling boxes.
Author Contributions
Conceptualization, F.-T.S. and W.-C.C.; Data curation, J.-H.Y. and Y.-T.C.; Formal analysis, J.-H.Y., W.-C.C. and Y.-T.C.; Funding acquisition, W.-C.C.; Investigation, Y.-T.C.; Methodology, F.-T.S., W.-C.C. and Y.-T.C.; Project administration, W.-C.C.; Resources, J.-H.Y. and C.-S.L.; Supervision, W.-C.C.; Validation, F.-T.S., W.-C.C. and Y.-T.C.; Writing—original draft, W.-C.C. and Y.-T.C.; Writing—review and editing, C.-S.L. and W.-C.C.
Acknowledgments
This research work was kindly supported by grants from the Ministry of Science and Technology (105-2313-B-005-005-) and in part by the Ministry of Education, Taiwan, R.O.C. under the Higher Education Sprout Project.
Conflicts of Interest
The authors submitted the MN1 bacteria patent application in Taiwan on 22 June 2018.
References
- Peng, S.; Tang, Q.; Zou, Y. Current Status and Challenges of Rice Production in China. Plant Prod. Sci. 2009, 12, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.; Boyle, C. The endophytic continuum. Mycol. Res. 2005, 109, 661–687. [Google Scholar] [CrossRef] [PubMed]
- Pillay, V.K.; Nowak, J. Inoculum density, temperature, and genotype effects on in vitro growth promotion and epiphytic and endophytic colonization of tomato (Lycopersicon esculentum L.) seedlings inoculated with a pseudomonad bacterium. Can. J. Microbiol. 1997, 43, 354–361. [Google Scholar] [CrossRef]
- Hurek, T.; Reinhold-Hurek, B. Azoarcus sp. strain BH72 as a model for nitrogen-fixing grass endophytes. J. Biotechnol. 2003, 106, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, M.; Balandreau, J.; Kwon, S.W.; Weon, H.Y.; Lakshminarasimhan, C. Effects of the inoculation of Burkholderia vietnamensis and related endophytic diazotrophic bacteria on grain yield of rice. Microb. Ecol. 2008, 55, 21–37. [Google Scholar] [CrossRef]
- Hardoim, P.R.; Hardoim, C.C.P.; van Overbeek, L.S.; van Elsas, J.D. Dynamics of seed-borne rice endophytes on early plant growth stages. PLoS ONE 2012, 7, e30438. [Google Scholar] [CrossRef]
- Etesami, H.; Hosseini, H.M.; Alikhani, H.A. Bacterial biosynthesis of 1-aminocyclopropane-1-caboxylate (ACC) deaminase, a useful trait to elongation and endophytic colonization of the roots of rice under constant flooded conditions. Physiol. Mol. Biol. Plants 2014, 20, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Ahemad, M.; Khan, M.S. Effect of fungicides on plant growth promoting activities of phosphate solubilizing Pseudomonas putida isolated from mustard (Brassica compestris) rhizosphere. Chemosphere 2012, 86, 945–950. [Google Scholar] [CrossRef]
- Radzuhn, B.; Lyr, H. On the mode of action of the fungicide etridiazole. Pestic. Biochem. Physiol. 1984, 22, 14–23. [Google Scholar] [CrossRef]
- Yang, C.; Hame, C.; Vujanovic, V.; Gan, Y. Fungicide: Modes of action and possible impact on nontarget microorganisms. ISRN Ecol. 2011, 2011, 1–8. [Google Scholar] [CrossRef]
- Fisher, D.J.; Hayes, A.L. Mode of action of the systemic fungicides furalaxyl, metalaxyl, and ofurace. Pestic. Sci. 1982, 13, 330–339. [Google Scholar] [CrossRef]
- Krieger, M.S.; Cook, W.L.; Kennard, L.M. Extraction of tricyclazole from soil and sediment with subcritical water. J. Agric. Food Chem. 2000, 48, 2178–2183. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.F.; Gossen, B.D.; Chang, K.F.; Turnbull, G.D.; Howard, R.J. Effect of seed damage and metalaxyl seed treatment on pythium seedling blight and seed yield of field pea. Can. J. Plant Sci. 2001, 81, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Sultan, N.S.; Raipat, B.S.; Sinha, M.P. Survival of plant growth promoting rhizobacteria (PGPR) in soil under pesticide stress. ISESCO J. Sci. Technol. 2013, 9, 82–88. [Google Scholar]
- Najam-ul-Sehar Ahmad, M.; Akhtar, M.F.Z.; Jamil, M.; Latif, M.; Ahmad, I. Pesticide tolerant plant growth promoting rhizobacteria isolated from rhizosphere of okra. Soil Environ. 2015, 34, 111–118. [Google Scholar]
- Council of Agriculture Executive Yuan, R.O.C. (Taiwan). Plant Protection Manual. 2018. Available online: https://otserv2.tactri.gov.tw/ppm/ (accessed on 15 November 2018).
- Martinez, C.; Levesque, C.A.; Belanger, R.R.; Tweddell, R.J. Evaluation of fungicides for the control of carrot cavity spot. Pest Manag. Sci. 2005, 61, 767–771. [Google Scholar] [CrossRef] [PubMed]
- Kunova, A.; Pizzatti, C.; Cortesi, P. Impact of tricyclazole and azoxystrobin on growth, sporulation and secondary infection of the rice blast fungus, Magnaporthe oryzae. Pest Manag. Sci. 2013, 69, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Zhan, F.; He, Y.; Yang, Y.; Li, Y.; Li, T.; Zhao, Z. Effects of tricyclazole on cadmium tolerance and accumulation characteristics of a dark septate endophyte (DSE), Exophiala pisciphila. Bull. Environ. Contam. Toxicol. 2016, 96, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Felsenstein, J. Confidence-limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, S.; Weber, R.P. The colorimetric estimation of IAA. Plant Physiol. 1951, 26, 192–195. [Google Scholar] [CrossRef]
- Brick, J.M.; Bostock, R.M.; Silversone, S.E. Rapid in situ assay for indole acetic acid production by bacteria immobilized on nitrocellulose membrane. Appl. Environ. Microbiol. 1991, 57, 535–538. [Google Scholar]
- Eckford, R.; Cook, F.D.; Saul, D.; Aislabie, J.; Foght, J. Free-living heterotrophic nitrogen-fixing bacteria isolated from fuel-contaminated Antarctic soils. Appl. Environ. Microbiol. 2012, 68, 5181–5185. [Google Scholar] [CrossRef]
- Ladha, J.K.; Barraquio, W.L.; Watanabe, I. Isolation and identification of nitrogen fixing Enterobacter cloacae and Klebsiella planticola associated with rice plants. Can. J. Microbiol. 1983, 29, 1301–1308. [Google Scholar] [CrossRef]
- Colnaghi, R.; Green, A.; He, L.; Rudnick, P.; Kennedy, C. Strategies for increased ammonium production in free-living or plant associated nitrogen-fixing bacteria. Plant Soil 1997, 194, 145–154. [Google Scholar] [CrossRef]
- Pikovskaya, R.I. Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Microbiologiya 1948, 17, 362–370. [Google Scholar]
- Szkop, M.; Bielawski, W. A simple method for simultaneous RP-HPLC determination of indolic compounds related to bacterial biosynthesis of indole-3-acetic acid. Antonie Van Leeuwenhoek 2013, 103, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, G.A.; Ashworth, J. Bacteriostatic action of nitrification inhibitors. Can. J. Microbiol. 1982, 28, 1093–1100. [Google Scholar] [CrossRef]
- Scheldeman, P.; Rodríguez-Díaz, M.; Goris, J.; Pil, A.; De Clerck, E.; Herman, L.; De Vos, P.; Logan, N.A.; Heyndrickx, M. Bacillus farraginis sp. nov., Bacillus fortis sp. nov. and Bacillus fordii sp. nov., isolated at dairy farms. Int. J. Syst. Evol. Microbiol. 2004, 54, 1355–1364. [Google Scholar] [CrossRef]
- Shivaji, S.; Chaturvedi, P.; Begum, Z.; Pindi, P.K.; Manorama, R.; Padmanaban, D.A.; Shouche, Y.S.; Pawar, S.; Vaishampayan, P.; Dutt, C.B.; et al. Janibacter hoylei sp. nov., Bacillus isronensis sp. nov. and Bacillus aryabhattai sp. nov., isolated from cryotubes used for collecting air from the upper atmosphere. Int. J. Syst. Evol. Microbiol. 2009, 59, 2977–2986. [Google Scholar] [CrossRef]
- Silva, M.P.; Pereira, C.A.; Junqueira, J.C.; Jorge, A.O.C. Methods of destroying bacterial spores. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2013; pp. 490–496. [Google Scholar]
- Boricha, H.; Fulekar, M.H. Pseudomonas plecoglossicida as a novel organism for the bioremediation of cypermethrin. Biol. Med. 2009, 1, 1–10. [Google Scholar]
- Rahmoune, B.; Morsli, A.; Khelifi-Slaoui, M.; Khelifi, L.; Strueh, A.; Erban, A.; Kopka, J.; Prell, J.; van Dongen, J.T. Isolation and characterization of three new PGPR and their effects on the growth of Arabidopsis and Datura plants. J. Plant Interact. 2017, 12, 1–6. [Google Scholar] [CrossRef]
- Pascual, J.; García-López, M.; Bills, G.F.; Genilloud, O. Pseudomonas granadensis sp. nov., a new bacterial species isolated from the Tejeda, Almijara and Alhama Natural Park, Granada, Spain. Int. J. Syst. Evol. Microbiol. 2015, 65, 625–632. [Google Scholar] [CrossRef]
- Rana, S.; Jindal, V.; Mandal, K.; Kaur, G.; Gupta, V.K. Thiamethoxam degradation by Pseudomonas and Bacillus strains isolated from agricultural soils. Environ. Monit. Assess. 2015, 187, 300. [Google Scholar] [CrossRef]
- Kämpfer, P.; Thummes, K.; Chu, H.I.; Tan, C.C.; Arun, A.B.; Chen, W.M.; Lai, W.A.; Shen, F.T.; Rekha, P.D.; Young, C.C. Pseudacidovorax intermedius gen. nov., sp. nov., a novel nitrogen-fixing betaproteobacterium isolated from soil. Int. J. Syst. Evol. Microbiol. 2008, 58, 491–495. [Google Scholar] [CrossRef]
- Liaqat, F.; Eltem, R. Identification and characterization of endophytic bacteria isolated from in vitro cultures of peach and pear rootstocks. 3 Biotech 2016, 6, 120. [Google Scholar] [CrossRef]
- Ding, Y.; Wang, J.; Liu, Y.; Chen, S. Isolation and identification of nitrogen-fixing bacilli from plant rhizospheres in Beijing region. J. Appl. Microbiol. 2005, 99, 1271–1281. [Google Scholar] [CrossRef] [Green Version]
- Nongkhlaw, F.M.; Joshi, S.R. Epiphytic and endophytic bacteria that promote growth of ethnomedicinal plants in the subtropical forests of Meghalaya, India. Rev. Biol. Trop. 2014, 62, 1295–1308. [Google Scholar] [CrossRef]
- Ambardar, S.; Vakhlu, J. Plant growth promoting bacteria from Crocus sativus rhizosphere. World J. Microbiol. Biotechnol. 2013, 29, 2271–2279. [Google Scholar] [CrossRef]
- Etesami, H.; Alikhani, H.A.; Hosseini, H.M. Indole-3-acetic acid (IAA) production trait, a useful screening to select endophytic and rhizosphere competent bacteria for rice growth promoting agents. MethodsX 2015, 2, 72–78. [Google Scholar] [CrossRef]
- García-Jaramillo, M.; Redondo-Gómez, S.; Barcia-Piedras, J.M.; Aguilar, M.; Jurado, V.; Hermosín, M.C.; Cox, L. Dissipation and effects of tricyclazole on soil microbial communities and rice growth as affected by amendment with alperujo compost. Sci. Total Environ. 2016, 550, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Moulas, C.; Petsoulas, C.; Rousidou, K.; Perruchon, C.; Karas, P.; Karpouzas, D.G. Effects of systemic pesticides imidacloprid and metalaxyl on the phyllosphere of pepper plants. Biomed. Res. Int. 2013, 2013, 969750. [Google Scholar] [CrossRef]
- Singh, N.; Gupta, S.; Marwa, N.; Pandey, V.; Verma, P.C.; Rathaur, S.; Singh, N. Arsenic mediated modifications in Bacillus aryabhattai and their biotechnological applications for arsenic bioremediation. Chemosphere 2016, 164, 524–534. [Google Scholar] [CrossRef]
- Ahmad, F.; Ahmad, I.; Khan, M.S. Indole acetic acid production by the indigenous isolates of Azotobacter and fluorescent Pseudomonas in the presence and absence of tryptophan. Turk. J. Biol. 2005, 29, 29–34. [Google Scholar]
- Liu, M.; Liu, X.; Cheng, B.S.; Ma, X.L.; Lyu, X.T.; Zhao, X.F.; Ju, Y.L.; Min, Z.; Fang, Y.L. Selection and evaluation of phosphate-solubilizing bacteria from grapevine rhizospheres for use as biofertilizers. Span. J. Agric. Res. 2016, 14, e1106. [Google Scholar] [CrossRef] [Green Version]
- Pailan, S.; Gupta, D.; Apte, S.; Krishnamurthi, S.; Saha, P. Degradation of organophosphate insecticide by a novel Bacillus aryabhattai strain SanPS1, isolated from soil of agricultural field in Burdwan, West Bengal, India. Int. Biodeterior. Biodegrad. 2015, 103, 191–195. [Google Scholar] [CrossRef]
- Santoyo, G.; Moreno-Hagelsieb, G.; Orozco-Mosqueda Mdel, C.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]
Figure 1.
The phylogenetic tree for the isolated endophytes with fungicide tolerance. Numbers shown on branches indicate the percentage of resembling the consensus branch obtained from 1000 times bootstrap analysis. Branches with <50% resembling rate were eliminated beforehand.
Figure 1.
The phylogenetic tree for the isolated endophytes with fungicide tolerance. Numbers shown on branches indicate the percentage of resembling the consensus branch obtained from 1000 times bootstrap analysis. Branches with <50% resembling rate were eliminated beforehand.

Figure 2.
Bacterial growth curve of Bacillus aryabhattai MN1 amended with the recommended dose of fungicides mixture.
Figure 2.
Bacterial growth curve of Bacillus aryabhattai MN1 amended with the recommended dose of fungicides mixture.

Figure 3.
Indole acetic acid (IAA) production in MN1 culture medium with different tryptophan concentrations.
Figure 3.
Indole acetic acid (IAA) production in MN1 culture medium with different tryptophan concentrations.

Figure 4.
IAA production of Bacillus aryabhattai MN1 under fungicide treatments.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Gradient program for indole acetic acid (IAA) detection with high-performance liquid chromatography (HPLC).
Table 1.
Gradient program for indole acetic acid (IAA) detection with high-performance liquid chromatography (HPLC).
| Time (Min) | Eluent A (%) | Eluent B (%) |
|---|---|---|
| 0.1% acetic acid (v/v, aqueous) | 80:20 Acetonitrile: H2O (v/v) | |
| 0 | 80 | 20 |
| 13 | 50 | 50 |
| 18 | 0 | 100 |
| 22 | 80 | 20 |
| 26 | Stop |
Table 2.
The 16S rDNA sequence analysis and plant growth-promoting activities of isolated rice endophytes.
Table 2.
The 16S rDNA sequence analysis and plant growth-promoting activities of isolated rice endophytes.
| Treatment † | Strain | Closest Identificaton | Similarity (%) | Nitrogen Fixation | Phosphate Solubilization | IAA ‡ (mg/L) |
|---|---|---|---|---|---|---|
| 10× Etridiazole | E1 | Pseudacidovorax intermedius | 100 | + | - | <1 |
| 10× Etridiazole | E2 | Rhizobium larrymoorei | 99.78 | + | - | 4 |
| 10× Etridiazole | E4 | Sphingomonas aquatilis | 100 | + | + | <1 |
| 10× Etridiazole | E5 | Flavobacterium ginsengiterrae | 97.1 | - | - | <1 |
| 10× Etridiazole | E6 | Brevundimonas vesicularis | 99.63 | - | - | <1 |
| 10× Etridiazole | E7 | Bacillus aryabhattai | 100 | + | - | 7.9 |
| 10× Etridiazole | E9 | Mycobacterium sphagni | 99.42 | + | + | <1 |
| 10× Etridiazole | E10 | Mycobacterium vanbaalenii | 99.71 | + | - | <1 |
| 10× Etridiazole | EN1 | Mycobacterium aromaticivorans | 99.06 | + | + | <1 |
| 10× Metalaxyl | M1 | Bacillus sporothermodurans | 98.6 | + | - | <1 |
| 5× Metalaxyl | M7 | Bacillus fortis | 98.17 | - | - | <1 |
| 1× Metalaxyl | MN1 | Bacillus aryabhattai | 100 | + | - | 8.7 |
| 5× Tricyclazole | T6 | Pseudomonas granadensis | 99.49 | + | + | 2.0 |
| 5× Tricyclazole | T9 | Bacillus fortis | 98.38 | + | - | 1.1 |
| 5× Tricyclazole | TN1 | Pseudomonas baetica | 99.57 | + | + | <1 |
| 5× Tricyclazole | TN3 | Pseudomonas plecoglossicida | 99.01 | + | + | <1 |
| 1× Tricyclazole | T3 | Bacillus marisflavi | 99.72 | + | - | <1 |
† The endophytes were isolated from rice seedlings grown on soils amended with different times of recommended doses of fungicides. ‡ IAA production ability was tested with 100 mg/L typtophan application and 24-h incubation period.
Table 3.
The growth of potential rice endophytic biofertilizers with fungicide amendment and their effect on rice germination.
Table 3.
The growth of potential rice endophytic biofertilizers with fungicide amendment and their effect on rice germination.
| Bacterial Strain | Fungicide Inhibition | Inoculation Test | ||||
|---|---|---|---|---|---|---|
| Etridiazole 7 mg/Kg | Metalaxy 19 mg/Kg | Tricyclazole 160 mg/Kg | Fungicides Mixture | Shoots Growth Promoting | Roots Growth Promoting | |
| Rhizobium larrymoorei E2 | − | − | + | + | + | + |
| Bacillus aryabhattai E7 | − | − | + | N/A | = | + |
| Bacillus aryabhattai MN1 | − | − | − | − | = | + |
| Pseudomonas granadensis T6 | − | − | + | N/A | + | + |
| Bacillus fortis T9 | + | − | N/A | N/A | N/A | N/A |
+: bacteria growth inhibited by fungicide or seedling growth promotion observed; −: bacteria growth not inhibited by fungicide, or no seedling growth promotion observed; =: not significantly different from the control; “N/A”: the experiments were not conducted.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shen, F.-T.; Yen, J.-H.; Liao, C.-S.; Chen, W.-C.; Chao, Y.-T. Screening of Rice Endophytic Biofertilizers with Fungicide Tolerance and Plant Growth-Promoting Characteristics. Sustainability 2019, 11, 1133. https://doi.org/10.3390/su11041133
AMA Style
Shen F-T, Yen J-H, Liao C-S, Chen W-C, Chao Y-T. Screening of Rice Endophytic Biofertilizers with Fungicide Tolerance and Plant Growth-Promoting Characteristics. Sustainability. 2019; 11(4):1133. https://doi.org/10.3390/su11041133
Chicago/Turabian StyleShen, Fo-Ting, Jui-Hung Yen, Chien-Sen Liao, Wen-Ching Chen, and Yi-Ting Chao. 2019. "Screening of Rice Endophytic Biofertilizers with Fungicide Tolerance and Plant Growth-Promoting Characteristics" Sustainability 11, no. 4: 1133. https://doi.org/10.3390/su11041133
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.