Environmental Sustainability: A Review of Termite Mound Soil Material and Its Bacteria



Food Security and Safety Niche, Faculty of Natural and Agricultural Sciences, North-West University, Private Mail Bag X2046, Mmabatho 2735, South Africa
*
Author to whom correspondence should be addressed.
Sustainability 2019, 11(14), 3847; https://doi.org/10.3390/su11143847
Submission received: 30 May 2019
/
Revised: 25 June 2019
/
Accepted: 4 July 2019
/
Published: 15 July 2019
(This article belongs to the Section Environmental Sustainability and Applications)
Abstract
:The high quantity of nutrients accumulated in termite mound soils have placed termite mound as a ‘gold mine’ for bacteria concentrations. However, over the years, not much attention has been given to the bacteria present in termite mound soil. This is because many studies have focused on approaches to manage termites which they see as menace to agricultural crops and buildings. Therefore, we aimed to evaluate the potential application of termite mound soil material and its bacteria for biotechnological purposes. This review has been grouped into four key parts: The termite mound as hotspot for bacterial concentration, the degradation of lignocellulose for biofuel production, termite mound soil as a soil amendment, and the role of termite mound soil and its bacteria in bioremediation and bio-filtration. Therefore, the effective usage of the termite mound soil material and its bacteria in an ecofriendly manner could ensure environmental sustainability.
1. Introduction
In the agroindustry, mound-building termites are known to have a detrimental effects on a number of crops, forest trees, and manmade structures [1,2]. Mound-building termites are a group of termite species, such as fungus-growing termites, that live in mounds [3]. Because of the destructive effect arising from the feeding habit of termites, many research studies have concentrated on the pest management of termites. However, of the roughly 2250 taxonomically well-known termite species, comparatively few—around ten to twenty percent—can negatively impact humans socially and economically [4]. Termites have been used as a biological pointer to assess soil quality and fertility. This is because they play a key part in soil transportation, methanogenesis, nitrogen fixation, acetogenesis, and nutrient circulation, thus improving the soil water content, pH, porosity, and organic carbon content [5,6,7]. Some termite societies like Macrotermes and Coptotermes are found in a mound (Figure 1) which they built by excavating soil particles from depths using resources from the surrounding soil [8].
Termite mound soils serve as unique natural habitats which harbor and uphold several microorganisms, of which the predominant ones are bacteria [9]. A termite mound is built by a mixture of clay components and organic carbon cemented by secretions, excreta, or saliva deposited by termites [10]. The architectural shapes of termite mounds include cathedral, dome, conical, lenticular, and mushroom-like [11]. These variations in shape depend on species type, ecological temperature conditions, clay availability, and the level of termite disturbance in the environment [12]. It has been reported that soil nutrients are accumulated in a termite mound, and their turnover plays a vital function in the ecosystem [13,14,15]. Research has revealed that termite mound soil is a “gold mine” of bacterial communities [16,17,18,19]. Termite mound soils could be used as a bulking agent [20] and for silo construction for short-term grain storage [21]. Furthermore, some bacteria isolated from termite mound soil could be utilized in an ecofriendly way as a potential material for antimicrobial production [10,22], biofertilizers, and biocontrol [23]. This can increase soil fertility and enhance crop production, thereby guaranteeing environmental sustainability (Figure 1) [24,25]. Environmental sustainability recognizes the need to improve and maintain the biophysical systems that sustain both the short and long-term quality of all life on Earth without compromising the health and diversity of natural ecosystems [26]. Due to the ecosystem services rendered by some bacteria present in termite mound, Miyagawa et al. [27] and Kaiser et al. [28] have called for proper research on the factors that support mound restoration by termites and further encourage practices that improve mound conservation. Though bacteria found in termite mound soils contribute to ecological services, there is scarcity of information on the possibility of using them as biotechnological products. Generally, biotechnology is seen as a fast emerging and influential field of technology for its useful role in health, food, and environmental sustainability [29]. Therefore, this review summarizes the variety of bacteria in termite mound soils and their possible contributions to lignocellulose degradation, biofuel production, bioremediation, and bio-filtration, as well as their potential to be a soil amendment.
2. Termite Mound as Hotspot for Bacteria Concentration
Bacteria secrete extracellular hydrolytic enzymes into soil that are responsible for the breakdown and utilization of some vital elements, and, as a result, they are seen as a major driving force controlling terrestrial habitats [30,31]. The nutrient richness of termite mounds not only contributes to plant growth but also supports soil beneficial bacteria living in termite mound soil [32,33]. The little research conducted on termite mound soils have reported the existence of useful bacteria therein (Table 1) which are important for biotechnological applications. Spirochetes, which are capable of producing acetate from H2 or CO2, were observed in termite mounds [34]. Spain et al. [35] stated that there are higher populations of bacteria in termite mound soil than their surrounding soil. This was supported by the research of Kumar, Tilak, Sivakumar, and Saranya [16], who stated that bacterial populations in open and closed termite mound soils were 75.5 × 105 cfu/g of soil and 65.5 × 105 cfu/g of soil, respectively, and they were higher than the normal soil with 30.5 × 105 cfu/g of soil bacterial population. This high bacterial diversity in mound soils has been credited to the high level of humidity and accessibility of substrate.
3. Degradation of Lignocellulose for Biofuel Production
Currently, there are many calls for the search for renewable forms of energy resources. This is as a result of limited areas for petroleum-based fuel production, which are continually reducing [38,39]. Over the years, animal feed and plant built biological materials consisting of carbohydrate have been used as biomass energy resources for alternative fuel production, and the fuels generated from these resources are called biofuels [40,41]. A successful use of renewable fuels from biomass instead of petroleum-based automotive fuels will benefit many societies and environments [42]. Biofuel usage reduces oil reliance and the effects of greenhouse gases, improves air quality, and creates new job opportunities [43]. Plants are made up of lignocellulose, which is the principal structural component of their cell walls [44]. Lignocellulose, a major source of renewable organic biomass, is an economical source of energy that is composed of hemicellulose, cellulose, and lignin [45]. Despite lignocellulose abundances in nature, the high cost of hydrolyzing them into simpler monosaccharides has made the economics of utilizing them unattractive [46]. This has led to the search for cost-effective, biological means of hydrolyzing lignocellulose [47]. Overall, direct and indirect fermentation are the two main mechanisms (Figure 2) presently used in biofuel exploration by targeting alcohol production. Direct fermentation entails the breaking down of starting plant materials into fermentable sugars, which are later converted into alcohol. Indirect fermentation uses pyrolysis of the starting plant materials to produce a mixture of carbon dioxide, hydrogen, and carbon monoxide. Thereafter, acetogenic bacteria are used to convert the produced gas into ethanol [48].
Recently, scientific interest in termite mound soils has increased, because they accommodate vast numbers of bacteria that secrete unusual cellulolytic enzymes that are useful in biofuel industries. For example, Glycosyl hydrolases, a useful enzyme for bioethanol, could be obtained from a termite mound [37,39,49]. As early as 1985, Jaishree and his co-researchers reported that the Cellulomonas species, a cellulose degrading bacteria, was isolated in a termite mound occupied by Odontotermes obesus in a semi-arid region [9]. In addition, genes from the bacteria responsible for xylan and cellulose hydrolysis have been identified in bacteria isolated from termite mound soil [50]. Termite mound soils are laden with bacteria phyla like Acidobacteria, Firmicutes, Actinobacteria, and Proteobacteria [36]. Most strains of these bacteria phyla are good degraders of plant biomass polysaccharides [51], and they can also decompose phenolic composites and lignin [52].
Fermenting bacteria, which have the ability to convert complex lignocellulose polymers into monosaccharides using lignocellulolytic enzymes which are essential for several vital industrial processes, have been isolated from termite mound soils [16]. Varghese, Agrawal, Sharma, Mandhan, and Mahajan [50] reported that thirty bacterial isolates from termite mound soil showed xylanase and cellulase activities from 0.45 to 6.02 and 55 to 380 IU/mL, respectively. Simple sugar fermentation from the lignocellulose breakdown by these enzymes is a remarkable prospect in biofuel production [53]. Thus, understanding the bacterial activities and environmental conditions controlling the transformation of massive quantities of carbon materials in termite mound soils could lead to new opportunities that can benefit the environment [30,50,54].
4. Termite Mound Soil as Soil Amendment
Soil amendments are materials combined with soil for the improvement of physical quality and, consequently, plant growth and health. Due to the unique physical and chemical properties of termite mound soils (which are brought about by termite activities), they are ‘hotspots’ for nutrient concentration in tropical and subtropical soils [15]. The physiochemical parameters of termite mounds and their surrounding soils are relatively different (Figure 3) [55]. Dhembare [13] and Deke, as well as Adugna and Fite [4], revealed that termite mound soils are richer in properties like clay content, organic carbon, copper, iron, potassium, zinc, magnesium, and phosphorus relative to their surrounding soils. The investigation of Fall et al. [56], showed that termite mound soil had approximately two-to-three times calcium and phosphorus, five times carbon and nitrogen, and 50 times ammonia and organic matter than their surrounding soil. The clay content in termite mound soils increases their soil porosity and water-holding capacity [8]. This then increases microbial activities [8,57] and therefore assists in the growth of plants [33]. Owing to the uniqueness of termite mound soils, many researchers have recommended their use as a soil amendment in low-input cropping systems, as seen in Table 2 [4,15,58].
5. The Role of Termite Mound Soil and Its Bacteria in Bioremediation and Bio-Filtering
Current research has given intensive attention to bioremediation, which is a process used to treat contaminated media by altering environmental conditions to stimulate the growth of microorganisms that degrade target pollutants [68,69]. Some bacteria have the ability to directly interact and remove heavy metals from contaminated soils and increase the transfer of these metals to the above ground biomass of plants [70]. The efficacy of Fluorescent pseudomonads in bending heavy metals has been reviewed by Wasi et al. [71] and a lot of these Pseudomonas strains are found in termite mound soils [17]. For instance, Pseudomonas monteillii isolated from termite mound soil were inoculated to Sorghum bicolor, with soil amended with 560 mg Cd kg−1 soil in research conducted by Duponnois et al. [72]. Their findings revealed that Sorghum bicolor inoculated with Pseudomonas monteillii had a higher Cd uptake than those not inoculated. Furthermore, they also reported that soil samples inoculated with Pseudomonas monteillii had a higher use of hydroxybutiric and ketoglutaric against fumaric acid in soil samples not inoculated with Pseudomonas monteillii. They therefore recommended that Fluorescent pseudomonads could aid in the phyto-extraction of heavy metals from soil. The surface characteristics and mineralogical compositions of the termite mound soil materials position them as an excellent adsorbent of metals. This became evident in an experiment where Abdus-Salam and Itiola [73] used termite mound soil material as a prospective tool for removing lead metal (Pb(II)) from aqueous solutions. The results from their experiment revealed that termite mound materials adsorbed Pb(II) up to 15.5 mg/g, and they then concluded that termite mound soil material is an efficient adsorbent of oxides of metals and even manganite silicate. Additionally reported in literature is the use of termite mound materials for defluoridation [74], the removal of arsenate from water [75], Zn(II) removal from aqueous solutions [76], the removal of dyes from wastewater [77], and the removal of chromium from aqueous solutions [78].
Termites can significantly decompose plant materials, and, through the process of decomposition, a significant amount of greenhouse gases are produced [79,80]. Ho et al. [81] reported that termite-resultant methane adds roughly 3–4% to the entire global methane budget, and mound-building termites contribute less than 1% of the total global emission (although this amount depends on the type of mound-building termite species) [82]. However, termite mound soils harbor a considerable number of methane oxidizing bacteria like Methylocystaceae and Methylococcaceae that act as biofilters. This bacteria can consume the methane produced by termites [36,81,82] and therefore reduce the amount of methane finally released to the surroundings [83,84]. In an experiment to test if the bacteria in termite mound soil can mitigate methane emission, Nauer et al. [85] reported that termite mounds with the help of the methanotrophic bacteria living in them were able to oxidize half of the methane formed by termites before it was released. Methanotrophs use enzymes—mainly methane monooxygenase—as a strategy to oxidize the methane produced by termites. This is because some genes (like the mmoX and pmoA genes) which encode subunits of methane monooxygenase have a strong affinity for methane and, even at a low concentration of less than 40 ppm, can oxidize methane [86].
6. Conclusions
Globally, biotechnology is seen as a fast developing and significant field of technology for its proficient part in health, food, and environmental sustainability. With the pursuit of using natural materials for biotechnological purposes, we therefore reviewed the potentiality of termite mound soil materials as a worthy tool. This is because they host beneficial bacteria that can be useful in the bioremediation of heavy metals, the bio-filtering of gases, and biofuel production, and they can serve as soil amendments. For the optimal utilization of termite mound materials for biotechnological purposes, there is a need for further and extensive research to exploit the metabolic capabilities of the bacteria present in termite mound soils, thereby uncovering their full potential. Furthermore, to ensure a sustainable termite mound materials application in the future, research should also be carried out to improve factors that stimulate mound restoration by termites, as well as to improve termite mound conservation.
Author Contributions
B.J.E., managed the literature searches and wrote the first draft of the manuscript, while O.O.B. provided academic input and thoroughly critiqued and proof read the draft. Both authors approved the article for publication.
Funding
This research received no external funding.
Acknowledgments
B.J.E. thanks South Africa’s National Research Foundation/The World Academy of Science African Renaissance grant (UID110909) that was of great help during his Doctoral programme. O.O.B. would like to thank the National Research Foundation, South Africa for the grant (UID81192) that has supported research in her lab.
Conflicts of Interest
The authors declare no potential conflict of interest.
Ethical Approval
This article does not contain any studies with human participants or animals performed by any of the authors.
References
- Bhardwaj, N.; Tomar, S. Indigenous knowledge of termite (Odontotermes obesus) control: A Review. Int. J. Adv. Sci. Res. 2018, 1, 124–127. [Google Scholar]
- Negassa, W.; Sileshi, G.W. Integrated soil fertility management reduces termite damage to crops on degraded soils in western Ethiopia. Agric. Ecosyst. Environ. 2018, 251, 124–131. [Google Scholar] [CrossRef]
- Calovi, D.S.; Bardunias, P.; Carey, N.; Scott Turner, J.; Nagpal, R.; Werfel, J. Surface curvature guides early construction activity in mound-building termites. Philos. Trans. R. Soc. B 2019, 374, 20180374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deke, A.L.; Adugna, W.T.; Fite, A.T. Soil physic-chemical properties in termite mounds and adjacent control soil in Miyo and Yabello Districts of Borana Zone, Southern Ethiopia. Amer. J. Agric. For. 2016, 4, 69–74. [Google Scholar]
- Dawes, T.Z. Reestablishment of ecological functioning by mulching and termite invasion in a degraded soil in an Australian savanna. Soil Biol. Biochem. 2010, 42, 1825–1834. [Google Scholar] [CrossRef]
- Khucharoenphaisan, K.; Sripairoj, N.; Sinma, K. Isolation and identification of actinomycetes from termite’s gut against human pathogen. Asian J. Anim. Vet. Adv. 2012, 7, 68–73. [Google Scholar] [CrossRef]
- Nithyatharani, R.; Kavitha, U.S. Termite soil as bio-Indicator of soil fertility. Int. J. Res. Appl. Sci. Eng. Technol. 2018, 6, 659–661. [Google Scholar] [CrossRef]
- Jouquet, P.; Guilleux, N.; Caner, L.; Chintakunta, S.; Ameline, M.; Shanbhag, R. Influence of soil pedological properties on termite mound stability. Geoderma 2016, 262, 45–51. [Google Scholar] [CrossRef]
- Kumari, R.; Sachdev, M.; Prasad, R.; Garg, A.P.; Sharma, S.; Giang, P.H.; Varma, A. Microbiology of termite hill (mound) and soil. In Intestinal Microorganisms of Termites and Other Invertebrates; Springer: Berlin, Germany, 2006; pp. 351–372. [Google Scholar]
- Sujada, N.; Sungthong, R.; Lumyong, S. Termite nests as an abundant source of cultivable actinobacteria for biotechnological purposes. Microbes Environ. 2014, 29, 211–219. [Google Scholar] [CrossRef]
- Abe, S.S.; Watanabe, Y.; Onishi, T.; Kotegawa, T.; Wakatsuki, T. Nutrient storage in termite (Macrotermes bellicosus) mounds and the implications for nutrient dynamics in a tropical savanna Ultisol. Soil Sci. Plant Nutr. 2011, 57, 786–795. [Google Scholar] [CrossRef]
- Arhin, E.; Boadi, S.; Esoah, M.C. Identifying pathfinder elements from termite mound samples for gold exploration in regolith complex terrain of the Lawra belt, NW Ghana. J. Afr. Earth. Sci. 2015, 109, 143–153. [Google Scholar] [CrossRef]
- Dhembare, A. Physico-chemical properties of termite mound soil. Arch. Appl. Sci. Res. 2013, 5, 123–126. [Google Scholar]
- Jouquet, P.; Guilleux, N.; Shanbhag, R.R.; Subramanian, S. Influence of soil type on the properties of termite mound nests in Southern India. Appl. Soil Ecol. 2015, 96, 282–287. [Google Scholar] [CrossRef]
- Menichetti, L.; Landi, L.; Nannipieri, P.; Katterer, T.; Kirchmann, H.; Renella, G. Chemical properties and biochemical activity of colonized and abandoned litter-feeding termite (Macrotermes spp.) mounds in chromic Cambisol Area on the Borana Plateau, Ethiopia. Pedosphere 2014, 24, 399–407. [Google Scholar]
- Kumar, P.; Tilak, M.; Sivakumar, K.; Saranya, K. Studies on the assessment of major nutrients and microbial population of termite mound soil. Int. J. For. Crop Improv. 2018, 9, 13–17. [Google Scholar] [CrossRef]
- Devi, R.; Thakur, R. Screening and identification of bacteria for plant growth promoting traits from termite mound soil. J. Pharm. Phytochem. 2018, 7, 1681–1686. [Google Scholar]
- Poulsen, M.; Hu, H.; Li, C.; Chen, Z.; Xu, L.; Otani, S.; Nygaard, S.; Nobre, T.; Klaubauf, S.; Schindler, P.M.; et al. Complementary symbiont contributions to plant decomposition in a fungus-farming termite. Proc. Natl. Acad. Sci. USA 2014, 111, 14500–14505. [Google Scholar] [CrossRef] [Green Version]
- Benndorf, R.; Guo, H.; Sommerwerk, E.; Weigel, C.; Garcia-Altares, M.; Martin, K.; Hu, H.; Küfner, M.; de Beer, Z.; Poulsen, M. Natural products from actinobacteria associated with fungus-growing termites. Antibiotics 2018, 7, 83. [Google Scholar] [CrossRef]
- Karak, T.; Sonar, I.; Paul, R.K.; Das, S.; Boruah, R.K.; Dutta, A.K.; Das, D.K. Composting of cow dung and crop residues using termite mounds as bulking agent. Bioresour. Technol. 2014, 169, 731–741. [Google Scholar] [CrossRef]
- Omobowale, M.O.; Armstrong, P.R.; Mijinyawa, Y.; Igbeka, J.C.; Maghirang, E.B. Maize storage in termite mound clay, concrete, and steel silos in the humid tropics: Comparison and effect on bacterial and fungal counts. Trans. Asabe 2016, 59, 1039–1048. [Google Scholar]
- Krishanti, N.P.R.A.; Zulfina, D.; Wikantyoso, B.; Zulfitri, A.; Yusuf, S. Antimicrobial production by an actinomycetes isolated from the termite nest. J. Trop. Life Sci. 2018, 8, 279–288. [Google Scholar]
- Enagbonma, B.J.; Babalola, O.O. Potentials of termite mound soil bacteria in ecosystem engineering for sustainable agriculture. Ann. Microbiol. 2019, 69, 211–219. [Google Scholar] [CrossRef]
- Yêyinou Loko, L.E.; Orobiyi, A.; Agre, P.; Dansi, A.; Tamò, M.; Roisin, Y. Farmers’ perception of termites in agriculture production and their indigenous utilization in Northwest Benin. J. Ethnobiol. Ethnomed. 2017, 13, 64. [Google Scholar] [CrossRef]
- Bama, P.S.; Ravindran, A.D. Dynamics of P sorption and solubilising activity in termite nest material. Asian J. Res. Soc. Sci. Hum. 2012, 2, 231–237. [Google Scholar]
- Smith, F. Environmental Sustainability: Practical Global Applications; CRC Press: Boca Raton, FL, USA, 2019; p. 279. [Google Scholar]
- Miyagawa, S.; Koyama, Y.; Kokubo, M.; Matsushita, Y.; Adachi, Y.; Sivilay, S.; Kawakubo, N.; Oba, S. Indigenous utilization of termite mounds and their sustainability in a rice growing village of the Central Plain of Laos. J. Ethnobiol. Ethnomed. 2011, 7, 24. [Google Scholar] [CrossRef]
- Kaiser, D.; Lepage, M.; Konaté, S.; Linsenmair, K.E. Ecosystem services of termites (Blattoidea: Termitoidae) in the traditional soil restoration and cropping system Zaï in northern Burkina Faso (West Africa). Agric. Ecosyst. Environ. 2017, 236, 198–211. [Google Scholar] [CrossRef]
- Ahmad, T.; Nabi, S.; Humera, Q. Biotechnology: A tool in termite management. In Termites and Sustainable Management: Economic Losses and Management; Khan, M.A., Ahmad, W., Eds.; Springer International Publishing: Cham, Germany, 2018; Volume 2, pp. 289–315. [Google Scholar]
- Gougoulias, C.; Clark, J.M.; Shaw, L.J. The role of soil microbes in the global carbon cycle: Tracking the below-ground microbial processing of plant-derived carbon for manipulating carbon dynamics in agricultural systems. J. Sci. Food Agric. 2014, 94, 2362–2371. [Google Scholar] [CrossRef]
- Haynes, R.J. Nature of the belowground ecosystem and its development during pedogenesis. Adv. Agron. 2014, 127, 43–109. [Google Scholar]
- Fall, S.; Hamelin, J.; Ndiaye, F.; Assigbetse, K.; Aragno, M.; Chotte, J.L.; Brauman, A. Differences between bacterial communities in the gut of a soil-feeding termite (Cubitermes niokoloensis) and its mounds. Appl. Environ. Microbiol. 2007, 73, 5199–5208. [Google Scholar] [CrossRef]
- Chauhan, A.K.; Maheshwari, D.K.; Dheeman, S.; Bajpai, V.K. Termitarium-inhabiting Bacillus spp. enhanced plant growth and bioactive component in turmeric (Curcuma longa L.). Curr. Microbiol. 2017, 74, 174–192. [Google Scholar] [CrossRef]
- Manjula, A.; Pushpanathan, M.; Sathyavathi, S.; Gunasekaran, P.; Rajendhran, J. Comparative analysis of microbial diversity in termite gut and termite nest using ion sequencing. Curr. Microbiol. 2016, 72, 267–275. [Google Scholar] [CrossRef]
- Spain, A.V.; Gordon, V.; Reddell, P.; Correll, R. Ectomycorrhizal fungal spores in the mounds of tropical Australian termites (Isoptera). Eur. J. Soil Biol. 2004, 40, 9–14. [Google Scholar] [CrossRef]
- Makonde, H.M.; Mwirichia, R.; Osiemo, Z.; Boga, H.I.; Klenk, H.P. 454 Pyrosequencing-based assessment of bacterial diversity and community structure in termite guts, mounds and surrounding soils. SpringerPlus 2015, 4, 471. [Google Scholar] [CrossRef]
- Manjula, A.; Sathyavathi, S.; Pushpanathan, M.; Gunasekaran, P.; Rajendhran, J. Microbial diversity in termite nest. Curr. Sci. 2014, 106, 1430–1434. [Google Scholar]
- Yilmaz, N.; Atmanli, A. Sustainable alternative fuels in aviation. Energy 2017, 140, 1378–1386. [Google Scholar] [CrossRef]
- Batista-García, R.A.; Del Rayo Sánchez-Carbente, M.; Talia, P.; Jackson, S.A.; O’Leary, N.D.; Dobson, A.D.; Folch-Mallol, J.L. From lignocellulosic metagenomes to lignocellulolytic genes: Trends, challenges and future prospects. Biofuels Bioprod. Bioref. 2016, 10, 864–882. [Google Scholar] [CrossRef]
- Williams, C.L.; Westover, T.L.; Emerson, R.M.; Tumuluru, J.S.; Li, C. Sources of biomass feedstock variability and the potential impact on biofuels production. Bioenergy Res. 2016, 9, 1–14. [Google Scholar] [CrossRef]
- Molino, A.; Larocca, V.; Chianese, S.; Musmarra, D. Biofuels production by biomass gasification: A review. Energies 2018, 11, 811. [Google Scholar] [CrossRef]
- Goldemberg, J.; Coelho, S.T.; Guardabassi, P.; Nastari, P.M. Bioethanol from Sugar: The Brazilian Experience; Springer Nature: New York, NY, USA, 2019; pp. 925–954. [Google Scholar]
- Nigam, P.S.; Singh, A. Production of liquid biofuels from renewable resources. Prog. Energy Com. Sci. 2011, 37, 52–68. [Google Scholar] [CrossRef]
- Sethi, A.; Scharf, M.E. Biofuels: Fungal, bacterial and insect degraders of lignocellulose. In e LS; John Wiley & Sons, Ltd.: Chichester, UK, 2013; p. 19. [Google Scholar]
- Sharma, H.K.; Xu, C.; Qin, W. Biological pretreatment of lignocellulosic biomass for biofuels and bioproducts: An overview. Waste Biomass Valorization 2019, 10, 235–251. [Google Scholar] [CrossRef]
- Putro, J.N.; Soetaredjo, F.E.; Lin, S.-Y.; Ju, Y.-H.; Ismadji, S. Pretreatment and conversion of lignocellulose biomass into valuable chemicals. RSC Adv. 2016, 6, 46834–46852. [Google Scholar] [CrossRef]
- Moreno, A.D.; Ibarra, D.; Alvira, P.; Tomás-Pejó, E.; Ballesteros, M. A review of biological delignification and detoxification methods for lignocellulosic bioethanol production. Crit. Rev. Biotechnol. 2015, 35, 342–354. [Google Scholar] [CrossRef]
- Elshahed, M.S. Microbiological aspects of biofuel production: Current status and future directions. J. Adv. Res. 2010, 1, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Talia, P.; Arneodo, J. Lignocellulose degradation by termites. In Termites and Sustainable Management; Springer: Cham, Germany, 2018; pp. 101–117. [Google Scholar]
- Varghese, L.M.; Agrawal, S.; Sharma, D.; Mandhan, R.P.; Mahajan, R. Cost-effective screening and isolation of xylano-cellulolytic positive microbes from termite gut and termitarium. Biotechnology 2017, 7, 108. [Google Scholar] [CrossRef] [Green Version]
- Koeck, D.E.; Pechtl, A.; Zverlov, V.V.; Schwarz, W.H. Genomics of cellulolytic bacteria. Curr. Opin. Biotechnol. 2014, 29, 171–183. [Google Scholar] [CrossRef]
- Bandounas, L.; Wierckx, N.J.; De Winde, J.H.; Ruijssenaars, H.J. Isolation and characterization of novel bacterial strains exhibiting ligninolytic potential. BMC Biotechnol. 2011, 11, 94. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, J.; Baskar, C. Lignocellulosic biomass for bioethanol production through microbes: Strategies to improve process efficiency. In Prospects of Renewable Bioprocessing in Future Energy Systems; Rastegari, A.A., Yadav, A.N., Gupta, A., Eds.; Springer International Publishing: Cham, Germany, 2019; pp. 357–386. [Google Scholar]
- You, Y.; Wang, J.; Huang, X.; Tang, Z.; Liu, S.; Sun, O.J. Relating microbial community structure to functioning in forest soil organic carbon transformation and turnover. Ecol. Evol. 2014, 4, 633–647. [Google Scholar] [CrossRef]
- Traoré, S.; Bottinelli, N.; Aroui, H.; Harit, A.; Jouquet, P. Termite mounds impact soil hydrostructural properties in southern Indian tropical forests. Pedobiologia 2019, 74, 1–6. [Google Scholar] [CrossRef]
- Fall, S.; Brauman, A.; Chotte, J.-L. Comparative distribution of organic matter in particle and aggregate size fractions in the mounds of termites with different feeding habits in Senegal: Cubitermes niokoloensis and Macrotermes bellicosus. Appl. Soil Ecol. 2001, 17, 131–140. [Google Scholar] [CrossRef]
- Fall, S.; Nazaret, S.; Chotte, J.L.; Brauman, A. Bacterial density and community structure associated with aggregate size fractions of soil-feeding termite mounds. Microb. Ecol. 2004, 48, 191–199. [Google Scholar] [CrossRef]
- Dhembare, A.; Pokale, A. Physico-Chemical Properties of Termite Mound Soil from Lohare, Ahmednagar Maharashtra; International E Publication: Ahmednagar Maharashtra, India, 2013; p. 243. [Google Scholar]
- Muvengwi, J.; Mbiba, M.; Nyenda, T. Termite mounds may not be foraging hotspots for mega-herbivores in a nutrient-rich matrix. J. Trop. Ecol. 2013, 29, 551–558. [Google Scholar] [CrossRef]
- Jouquet, P.; Tessier, D.; Lepage, M. The soil structural stability of termite nests: Role of clays in Macrotermes bellicosus (Isoptera, Macrotermitinae) mound soils. Eur. J. Soil Biol. 2004, 40, 23–29. [Google Scholar] [CrossRef]
- Adhikary, N.; Erens, H.; Weemaels, L.; Deweer, E.; Mees, F.; Mujinya, B.B.; Baert, G.; Boeckx, P.; Van Ranst, E. Effects of spreading out termite mound material on ferralsol fertility, Katanga, DR Congo. Comm. Soil Sci. Plant Anal. 2016, 47, 1089–1100. [Google Scholar] [CrossRef]
- Garba, M.; Cornelis, W.M.; Steppe, K. Effect of termite mound material on the physical properties of sandy soil and on the growth characteristics of tomato (Solanum lycopersicum L.) in semi-arid Niger. Plant Soil 2011, 338, 451–466. [Google Scholar] [CrossRef]
- Batalha, L.; Da Silva Filho, D.; Martius, C. Using termite nests as a source of organic matter in agrosilvicultural production systems in Amazonia. Sci. Agric. 1995, 52, 318–325. [Google Scholar] [CrossRef] [Green Version]
- Watson, J. The use of mounds of the termite Macrotermes falciger (Gerstäcker) as a soil amendment. Eur. J. Soil Sci. 1977, 28, 664–672. [Google Scholar] [CrossRef]
- Bama, P.S.; Ravindran, A.D. Influence of Combined Termite Mound Materials and Inorganic Fertilizers on Growth Parameters of Maize under Non Sterilized Pot Culture Study. Elixir Appl. Zool. 2018, 125, 52303–52305. [Google Scholar]
- Kisa, M.; Duponnois, R.; Assikbetse, K.; Ramanankierana, H.; Thioulouse, J.; Lepage, M. Litter-forager termite mounds enhance the ectomycorrhizal symbiosis between Acacia holosericea A. Cunn. Ex G. Don and Scleroderma dictyosporum isolates. FEMS Microbiol. Ecol. 2006, 56, 292–303. [Google Scholar]
- Suzuki, S.; Noble, A.D.; Ruaysoongnern, S.; Chinabut, N. Improvement in water-holding capacity and structural stability of a sandy soil in Northeast Thailand. Arid Land Res. Manag. 2007, 21, 37–49. [Google Scholar] [CrossRef]
- Das, S.; Dash, H.R.; Chakraborty, J. Genetic basis and importance of metal resistant genes in bacteria for bioremediation of contaminated environments with toxic metal pollutants. Appl. Microbiol. Biotechnol. 2016, 100, 2967–2984. [Google Scholar] [CrossRef]
- Dangi, A.K.; Sharma, B.; Hill, R.T.; Shukla, P. Bioremediation through microbes: Systems biology and metabolic engineering approach. Crit. Rev. Biotechnol. 2019, 39, 79–98. [Google Scholar] [CrossRef]
- Burd, G.I.; Dixon, D.G.; Glick, B.R. Plant growth-promoting bacteria that decrease heavy metal toxicity in plants. Can. J. Microbiol. 2000, 46, 237–245. [Google Scholar] [CrossRef]
- Wasi, S.; Tabrez, S.; Ahmad, M. Use of Pseudomonas spp. for the bioremediation of environmental pollutants: A review. Environ. Monit. Assess. 2013, 185, 8147–8155. [Google Scholar] [CrossRef]
- Duponnois, R.; Kisa, M.; Assigbetse, K.; Prin, Y.; Thioulouse, J.; Issartel, M.; Moulin, P.; Lepage, M. Fluorescent pseudomonads occuring in Macrotermes subhyalinus mound structures decrease Cd toxicity and improve its accumulation in sorghum plants. Sci. Total Environ. 2006, 370, 391–400. [Google Scholar] [CrossRef]
- Abdus-Salam, N.; Itiola, A.D. Potential application of termite mound for adsorption and removal of Pb(II) from aqueous solutions. J. Iran. Chem. Soc. 2012, 9, 373–382. [Google Scholar] [CrossRef]
- Fufa, F.; Alemayehu, E.; Lennartz, B. Defluoridation of groundwater using termite mound. Water Air Soil Pollut. 2013, 224, 1552. [Google Scholar] [CrossRef]
- Fufa, F.; Alemayehu, E.; Lennartz, B. Sorptive removal of arsenate using termite mound. J. Environ. Manag. 2014, 132, 188–196. [Google Scholar] [CrossRef]
- Abdus-Salam, N.; Bello, M.O. Kinetics, thermodynamics and competitive adsorption of lead and zinc ions onto termite mound. Int. J. Environ. Sci. Technol. 2015, 12, 3417–3426. [Google Scholar] [CrossRef] [Green Version]
- Ayanda, O.S.; Amodu, O.S.; Adubiaro, H.; Olutona, G.O.; Ebenezer, O.T.; Nelana, S.M.; Naidoo, E.B. Effectiveness of termite hill as an economic adsorbent for the adsorption of alizarin red dye. J. Water Reuse Desalin. 2019, 9, 83–93. [Google Scholar] [CrossRef]
- Araújo, B.; Reis, J.; Rezende, E.; Mangrich, A.; Wisniewski, A., Jr.; Dick, D.; Romão, L. Application of termite nest for adsorption of Cr (VI). J. Environ. Manag. 2013, 129, 216–223. [Google Scholar] [CrossRef]
- Da Costa, R.R.; Hu, H.; Pilgaard, B.; Vreeburg, S.M.; Schückel, J.; Pedersen, K.S.; Kračun, S.K.; Busk, P.K.; Harholt, J.; Sapountzis, P. Enzyme activities at different stages of plant biomass decomposition in three species of fungus-growing termites. Appl. Environ. Microbiol. 2018, 84, e01815–e01817. [Google Scholar] [CrossRef]
- Gomati, V.; Ramasamy, K.; Kumar, K.; Sivaramaiah, N.; Mula, R.V. Green house gas emissions from termite ecosystem. Afr. J. Environ. Sci. Technol. 2011, 5, 56–64. [Google Scholar]
- Ho, A.; Erens, H.; Mujinya, B.B.; Boeckx, P.; Baert, G.; Schneider, B.; Frenzel, P.; Boon, N.; Van Ranst, E. Termites facilitate methane oxidation and shape the methanotrophic community. Appl. Environ. Microbiol. 2013, 79, 7234–7240. [Google Scholar] [CrossRef]
- Jamali, H.; Livesley, S.; Hutley, L.B.; Fest, B.; Arndt, S. The relationships between termite mound CH4/CO2 emissions and internal concentration ratios are species specific. Biogeosciences 2013, 10, 2229–2240. [Google Scholar] [CrossRef]
- Jamali, H.; Livesley, S.J.; Dawes, T.Z.; Hutley, L.B.; Arndt, S.K. Termite mound emissions of CH4 and CO2 are primarily determined by seasonal changes in termite biomass and behaviour. Oecologia 2011, 167, 525–534. [Google Scholar] [CrossRef]
- Sugimoto, A.; Inoue, T.; Kirtibutr, N.; Abe, T. Methane oxidation by termite mounds estimated by the carbon isotopic composition of methane. Global Biogeochem. Cycles 1998, 12, 595–605. [Google Scholar] [CrossRef]
- Nauer, P.A.; Hutley, L.B.; Arndt, S.K. Termite mounds mitigate half of termite methane emissions. Proc. Nat. Acad. Sci. USA 2018, 115, 13306–13311. [Google Scholar] [CrossRef] [Green Version]
- Knief, C.; Dunfield, P.F. Response and adaptation of different methanotrophic bacteria to low methane mixing ratios. Environ. Microbiol. 2005, 7, 1307–1317. [Google Scholar] [CrossRef]
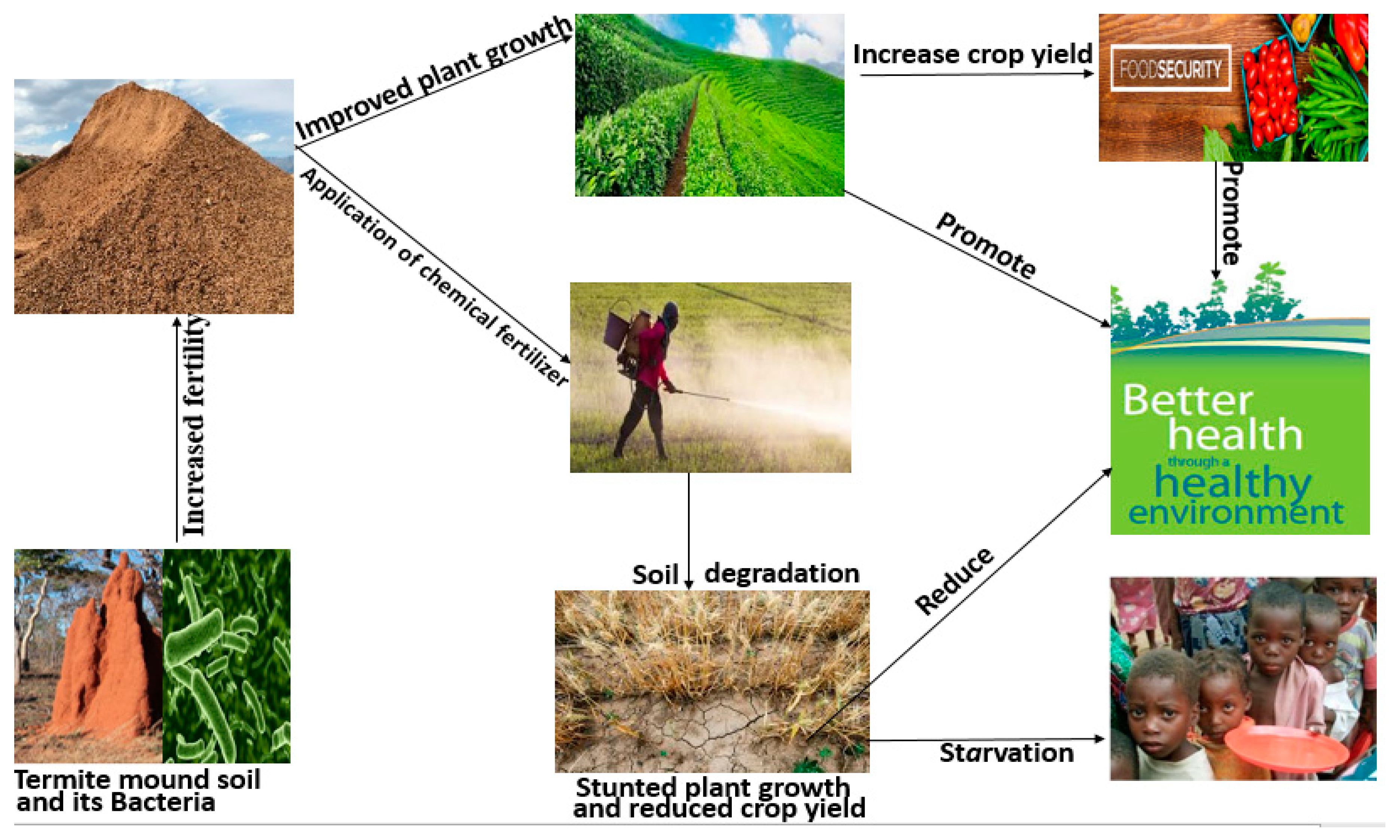
Figure 1.
Termite mound soil and its bacteria’s role on human health and environmental sustainability.
Figure 1.
Termite mound soil and its bacteria’s role on human health and environmental sustainability.

Figure 2.
Direct and indirect fermentation mechanisms for biofuel production.

Figure 3.
Some soil physiochemical parameter differences between termite mound soil and surrounding soil. References: A = [59], B = [13], C = [4], D = [60], E = [61], and F = [62].


{kind=link}
{kind=link}
{kind=link}
Table 1.
Bacteria reported present in termite mound soil and the method used in identifying them.
| Molecular Method Used | Bacteria Reported | Reference |
|---|---|---|
| 454-pyrosequencing based analysis of 16S ribosomal ribonucleic acid gene sequences | Nitrospirae, Cyanobacteria, Bacteroidetes, Spirochaetes, Actinobacteria, Fibrobacteres, Candidate division TM7, Chloroflexi, Proteobacteria, WCHB1-60, Elusimicrobia, Planctomycetes, Spirochaetes, Chlorobi, Gemmatimonadetes, Armatimonadetes, Acidobacteria, SM2F11, Firmicutes, Candidate division WS3 | [36] |
| PCR-denaturing gradient gel electrophoresis analysis, cloning and sequencing of PCR-amplified 16S rRNA gene fragments | Firmicutes, Actinobacteria, Chloroflexi | [32] |
| Ion Sequencing of V3 hypervariable region of 16S ribosomal deoxyribonucleic acid | Cyanobacteria, Proteobacteria, Chlorobi, Actinobacteria, Deinococcus-Thermus, Firmicutes, Synergistetes, Spirochaetes, and Bacteroidetes, | [34] |
| Bacterial tag encoded amplicon pyrosequencing | Gemmatimonadetes, Fibrobacteres, Nitrospirae, Verrucomicrobia, Bacteroidetes, Actinobacteria, Deinococcus, Planctomycetes, Chlamydiae, Firmicutes, Chloroflexi, Tenericutes, Acidobacteria, and Proteobacteria | [37] |
| Isolation and cultivation | Pseudonocardia, Nocardia, Micromonospora, Streptomyces, and Amycolatopsis | [10] |
| Isolation and cultivation | Bacillus, Pseudomonas, Nocardia, and Streptomyces | [16] |
Table 2.
Termite mound soil application as a soil amendment.
| Test Plant | Effect on the Plant | Reference |
|---|---|---|
| Solanum lycopersicum | The soil amended with termite mound soils resulted in better plant height, as well as an increase in leave number, fruits, and dry matter than those plant grown on unamended soil. | [62] |
| Oryza sativa L. and Phaseolus vulgaris L. | Improve the growth of Oryza sativa L. and Phaseolus vulgaris L. | [27] |
| Solatium melongena | Combined use of 200 g of termite mound material with NPK led to a substantial increase in Solatium melongena production. | [63] |
| Lolium perenne | Lolium perenne gave higher dry-matter yields with substrates derived from termite mounds than the comparable soil. | [64] |
| Zea mays | Combined use of termite mound materials and inorganic fertilization significantly increased the Zea mays growth and yield. | [65] |
| Acacia holosericea | Termite mound materials consisting of Pseudomonas monteilii species enhanced the ectomycorrhizal development between Acacia holosericea and Scleroderma dictyosporum. | [66] |
| Sorghum sudanensis | Combining sandy soil with termite mound materials at a proportion of 120 Mg/ha improved porosity and transformed the pore size distribution, thus causing a stepping up in the obtainable water content for the crop growth. | [67] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Enagbonma, B.J.; Babalola, O.O. Environmental Sustainability: A Review of Termite Mound Soil Material and Its Bacteria. Sustainability 2019, 11, 3847. https://doi.org/10.3390/su11143847
AMA Style
Enagbonma BJ, Babalola OO. Environmental Sustainability: A Review of Termite Mound Soil Material and Its Bacteria. Sustainability. 2019; 11(14):3847. https://doi.org/10.3390/su11143847
Chicago/Turabian StyleEnagbonma, Ben Jesuorsemwen, and Olubukola Oluranti Babalola. 2019. "Environmental Sustainability: A Review of Termite Mound Soil Material and Its Bacteria" Sustainability 11, no. 14: 3847. https://doi.org/10.3390/su11143847
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.