1. Introduction
Foliage plant production represents an important agricultural industry worldwide with a net wholesale value of €50 billion [
1,
2]. The most-used lamps for greenhouse plant production are metal halide, high-pressure sodium, fluorescent lamps, and incandescent lamps [
3]. Currently, in a highly demanding market, growers are using light emitting diodes (LEDs) since they can produce more saleable plants with the desired characteristics, and LEDs may also optimize the spectra for each crop and stage of development [
4,
5]. The main advantages of LEDs in commercial plant production are the following: spectral specificity, relative energy efficiency, and longer lifetimes than the current industry standard lamps (e.g., fluorescent and high-pressure sodium ones) [
6,
7].
Light parameters such as spectral specificity and quantity strongly influence plant development and physiology through photosynthesis and photomorphogenetic photoreceptors [
8]. Wavelengths in red (R) and blue (B) regions are more effective in the photosynthetic process than wavelengths in other regions [
9]. Blue light plays a role in many physiological plant processes such as photomorphogenesis, phototropism, stomatal opening, and photosynthesis [
10]. Red light is involved in stem elongation, root-to-shoot ratio, chlorophyll content, and photosynthetic apparatus development [
11].
We investigated the responses of
Dieffenbachia maculata “Compacta” grown under different light treatments.
D. maculata, or “dumb cane”, is an ornamental perennial monocot plant native to the tropical Americas, belonging to the family Araceae. It is prized for its attractive variegated foliage, tolerance to interior environments, and ease of production [
12]. There is little information on the effects of light conditions on the growth of this species, with one of the few studies being an experiment conducted by Heo et al. [
13] concerning the effect of supplementary LED lighting on the biomass and physiological parameters in
Dieffenbachia amoena. Therefore, in this trial, a pot experiment with
D. maculata plants was carried out in order to determine the effects of different light treatments on growth, nutritional, physiological, and photosynthetic parameters and to establish a methodology to evaluate the effect of photosynthetically active radiation (PAR) emitted by lamps on biomass.
2. Materials and Methods
2.1. Plant Material and Experimental Conditions
One hundred rooted cuttings (plants) of D. maculata “Compacta” were acquired from a commercial nursery with an initial total dry weight (TDW) of 3.40 ± 0.20 g. Each plant was transplanted into 1.5-L plastic containers filled with Sphagnum peat-moss. During the trial (8 weeks), the pots were organized for the research in a growth chamber where they were randomly assigned to each light treatment. The growth chamber was set at a constant (day/night) temperature of 25 ± 2 °C, and a relative humidity of 55% ± 3% to minimize any potentially confounding effect of a diurnal difference in temperature or relative humidity on extension growth.
2.2. Experimental Design and Treatments
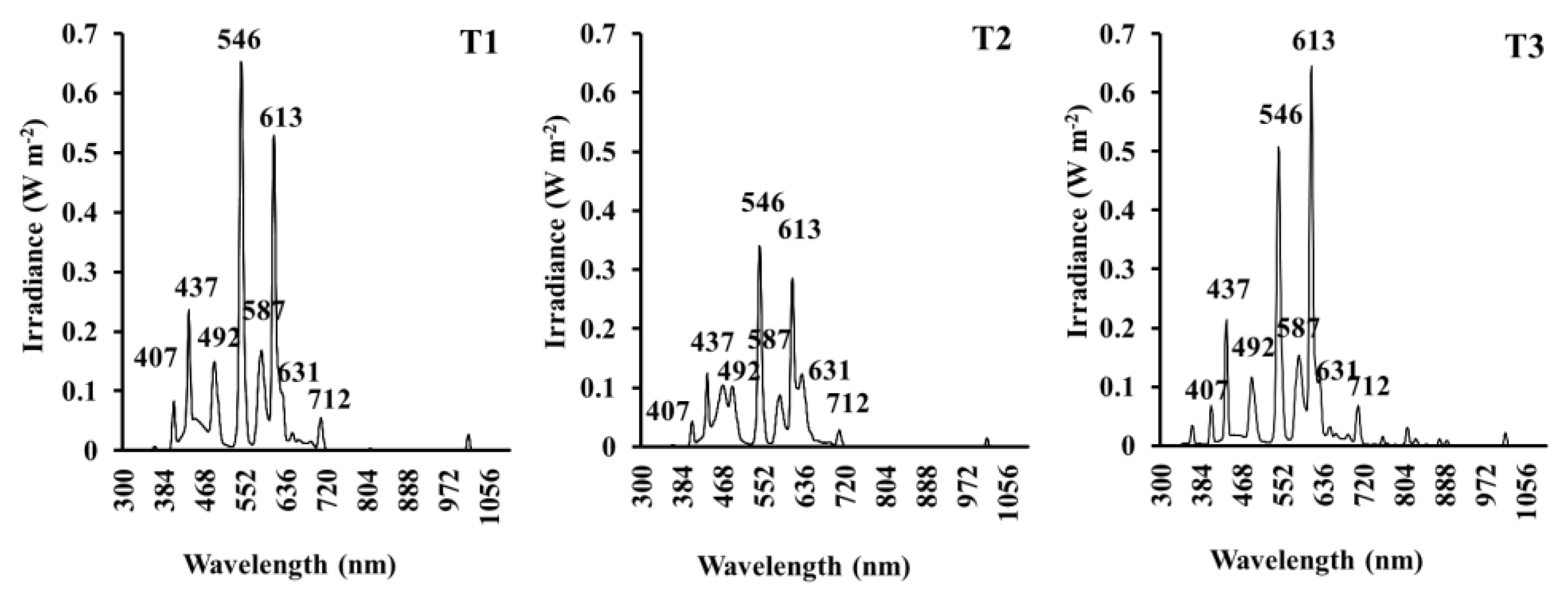
Fluorescent lamps have traditionally been used in indoor garden designs due to their low power consumption and acceptable light conditions for the human eye. Nevertheless, the implementation of new lighting technologies (LEDs) may allow for an improvement in the spectral light available in indoor gardens. The experiment was made up of three light treatments: treatment T1 (fluorescent TLD 18 W), treatment T2 (fluorescent TLD 18 W + Pure Blue and Pure Red Mix-Light-Emitting Diodes (BR-LEDs), and treatment T3 (high-efficiency fluorescent TL5 35 W). The first treatment used 4 light Philips TCS097 luminaires × 2 lamps (Philips Lighting Spain, Madrid, Spain) with a total power of 144 W. The second treatment used 3 light Philips TCS097 luminaires × 2 lamps (Philips Lighting Spain, Madrid) + BR-LEDs RGB (4 lines ALUM 40 x 25 LED SMD RGB x 9 W with console DN-RGB FIBER LIGHT, Modular Signs, Murcia, Spain) with the same power as T1. The third treatment used 2 light MAXOS 4M691 luminaires × 2 lamps (Philips Lighting Spain, Madrid, Spain) with a total power of 140 W. To determine the spectral distribution scans of the different light treatments, recordings were conducted at 400–1100 nm with 1-nm steps using a calibrated spectroradiometer (LI-COR 1800, Lincoln, NE, USA) at the canopy level.
Agronomic characterization of each light treatment was assessed following the methodology established by Baille et al. [
14]. The plants were grown under a photoperiod of 16 h of light and 8 h of dark and were irrigated manually every day with a Steiner nutrient solution [
15] in order to avoid any water or nutrient limitation. The experimental design consisted of three light treatments, four blocks for each treatment, and five plants (one plant per pot) per block, giving a total of 60 plants plus border plants. The selection of these light treatments was undertaken following the recommendations given by local nursery growers for
Dieffenbachia plants.
The quantum flux is defined as the number of photons (in µmol) per second and unit area on a surface (µmol·m
−2·s
−1). To calculate the quantum flux considering the value of each wavelength, we used the following abbreviated equation:
where I is irradiance expressed in W·m
−2.
To calculate the integral quantum flux of PAR, we used the following equation:
According to these equations, the values of the integral quantum flux were the following: 112.38, 77.37, and 101.22 µmol·m−2·s−1 for treatments T1, T2, and T3, respectively.
2.3. Growth Parameters
At the end of the experimental period, the plants were harvested, and the substrate was gently washed from the roots of five plants per treatment. The plants were divided into roots (R), stems (S), and leaves (L), and the respective fresh weights (FW) were measured. Roots, stems, and leaves were then oven-dried at 60 °C until they reached a constant weight to measure their respective dry weights (DW). These dry weights were used to calculate several plant parameters as indicated by Ryser and Lambers [
16] and García-Caparrós et al. [
2]: the leaf weight ratio (LWR; leaf DW per unit plant DW), the stem weight ratio (SWR; stem DW per unit plant DW), and the relative root weight ratio (RWR; root DW per unit plant DW). The TDW was calculated as the sum of leaf, stem, and root DWs. The fresh and dry weights of roots, stems, and leaves were used to calculate the water content (WC) as indicated by Ben Amor et al. [
17]:
2.4. Analysis of Mineral Elements
The determination of nutrient concentrations in roots, stems, and leaves was carried out on oven-dried samples which were ground in a mill and divided into two subsamples. In one subsample, the soluble ionic form of nitrate (NO
3−) was quantified using water extraction and HPLC, as described by Csáky and Martínez-Grau [
18]. The other subsample was mineralized with sulphuric acid (H
2SO
4, 96%) in the presence of P-free hydrogen peroxide (H
2O
2, 30% (w/v)) at 300 °C and used for the determination of organic N [
19], total P [
20], and K
+ [
21] concentrations. The total N concentration was calculated as the sum of the organic N and NO
3−-N concentration.
2.5. Physiological Parameters
The concentrations of proline, total soluble sugars (TSS), starch, and indole 3-acetic acid (IAA) were determined in the leaves of five plants randomly selected per treatment at harvest. The free proline, TSS, starch, and IAA concentrations were determined in the alcoholic extract supernatant. The free proline concentration was determined by the ninhydrin reagent method. Free proline concentration was expressed as µg·g
−1·FW. The total soluble sugars concentration was determined by the anthrone reagent method. The starch concentration was determined using the oven-dried residue of total soluble sugars determination. The total soluble sugars and starch concentration was expressed as mg glucose g
−1·FW [
22]. The IAA concentration was determined by the Salkowski reagent method and was expressed as mg·g
−1·FW [
23].
2.6. Pigment Concentrations
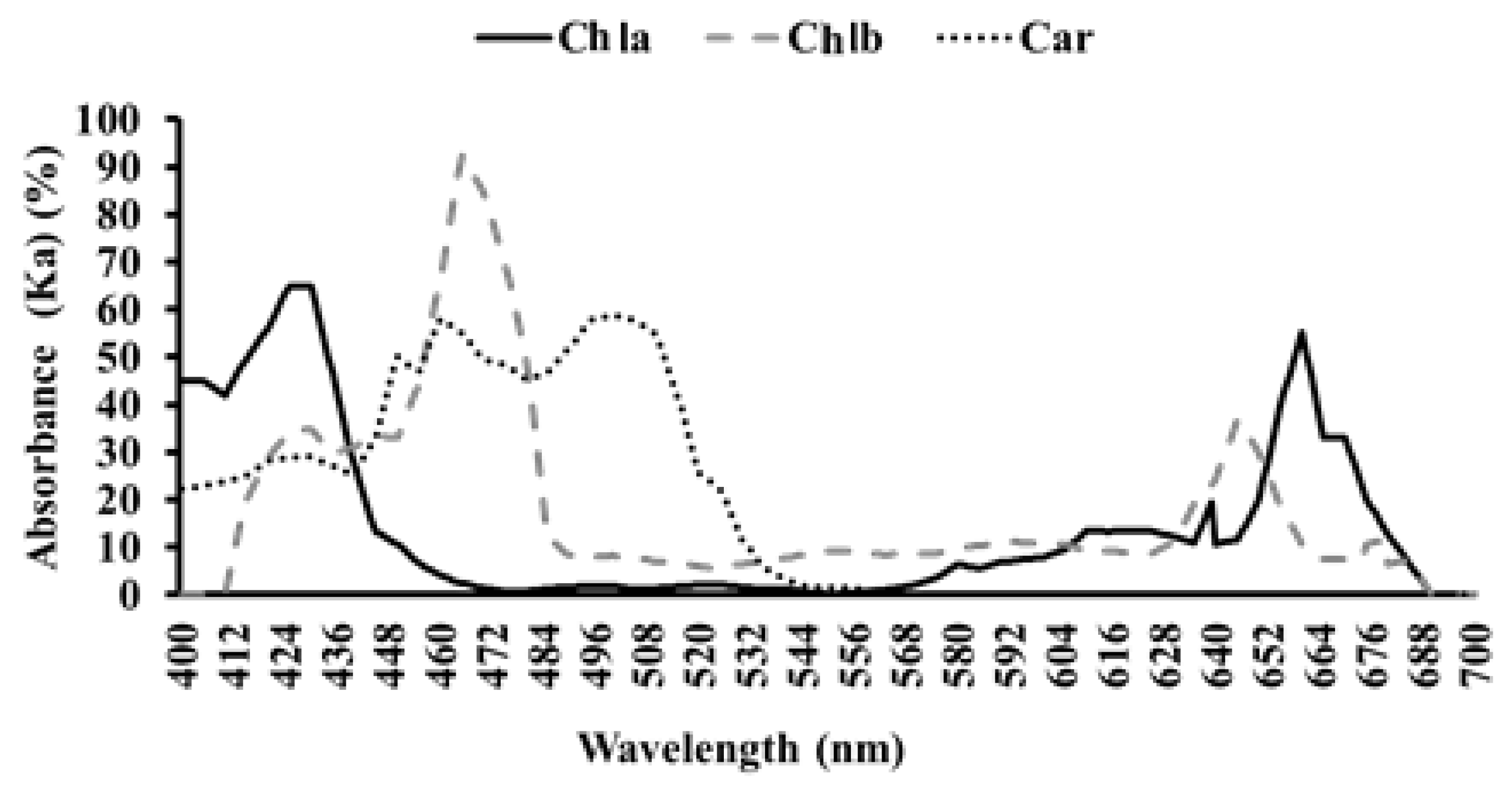
To determine the concentrations of pigments (chlorophylls and carotenoids) in leaves, five plants were randomly selected per treatment at harvest. Extraction of chlorophyll a and b (Chl a and Chl b) and carotenoids were performed by submerging 0.2 g of fresh mature leaf material in methanol in the dark at room temperature (15 °C) for 24 h. The supernatant was removed, and the photosynthetic pigment concentrations were determined colorimetrically at their respective wavelengths following the methodology of Wellburn [
24]. Pigment concentrations were expressed as mg·g
−1·FW.
2.7. Plant´s Harvesting Lamp PAR Model
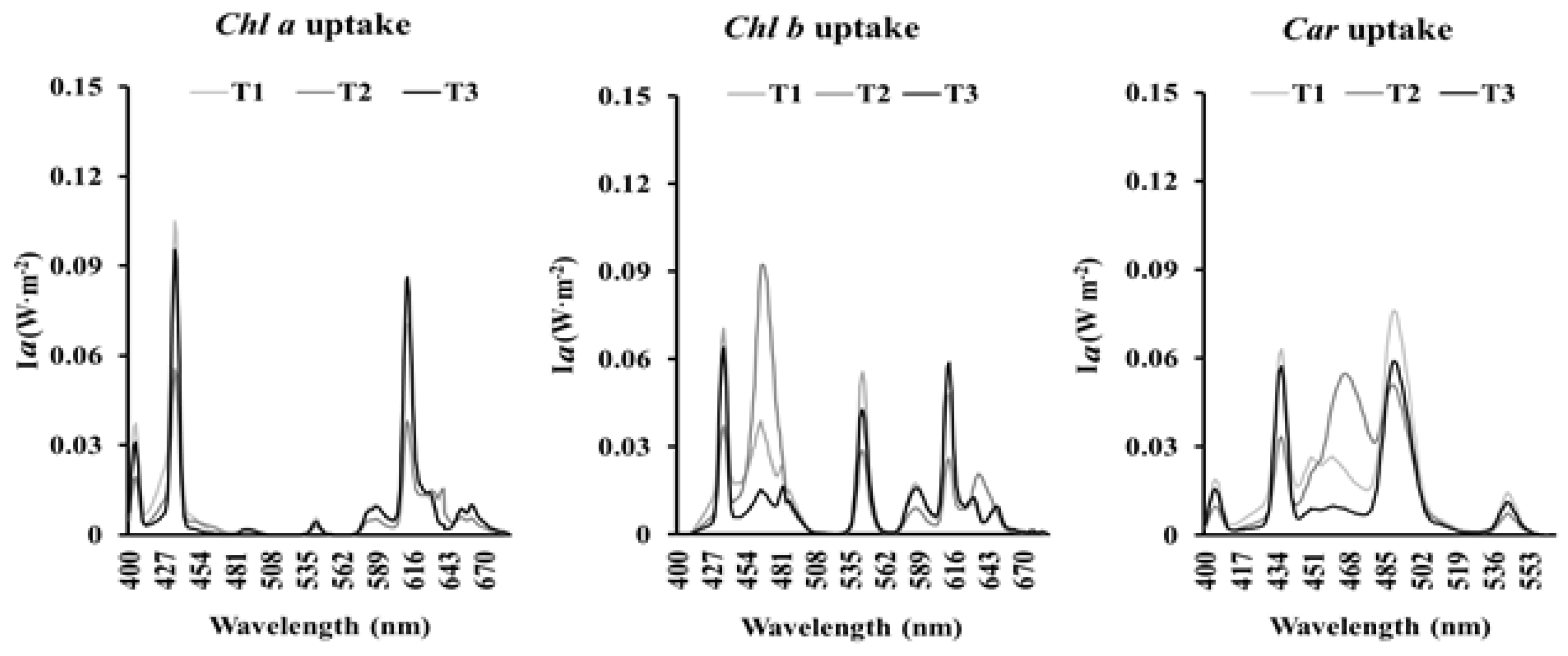
To study the energy collected by a plant which could to be transformed into biomass, we considered the concepts of theoretical irradiance energy absorbed by pigments (Ia), the theoretical use of spectral energy by each pigment of the plant (USEpigment), and the theoretical integrated use of spectral energy (IUSE) by a crop as the capacity of harvesting radiation energy by a plant.
The theoretical irradiance energy absorbed by pigments (Ia) (chlorophyll a and b and carotenoids) was calculated using the following equation:
where:
Ia (car, chl a, chl b): total irradiance energy absorbed by each pigment expressed in W·m−2;
Ka (car, chl a, chl b): percentage of energy absorbed by pigment according to each wavelength following the recommendations established by Whitmarsh and Govindjee [
25]; and
Ie: irradiance emitted by each lamp per wavelength expressed in W·m−2.
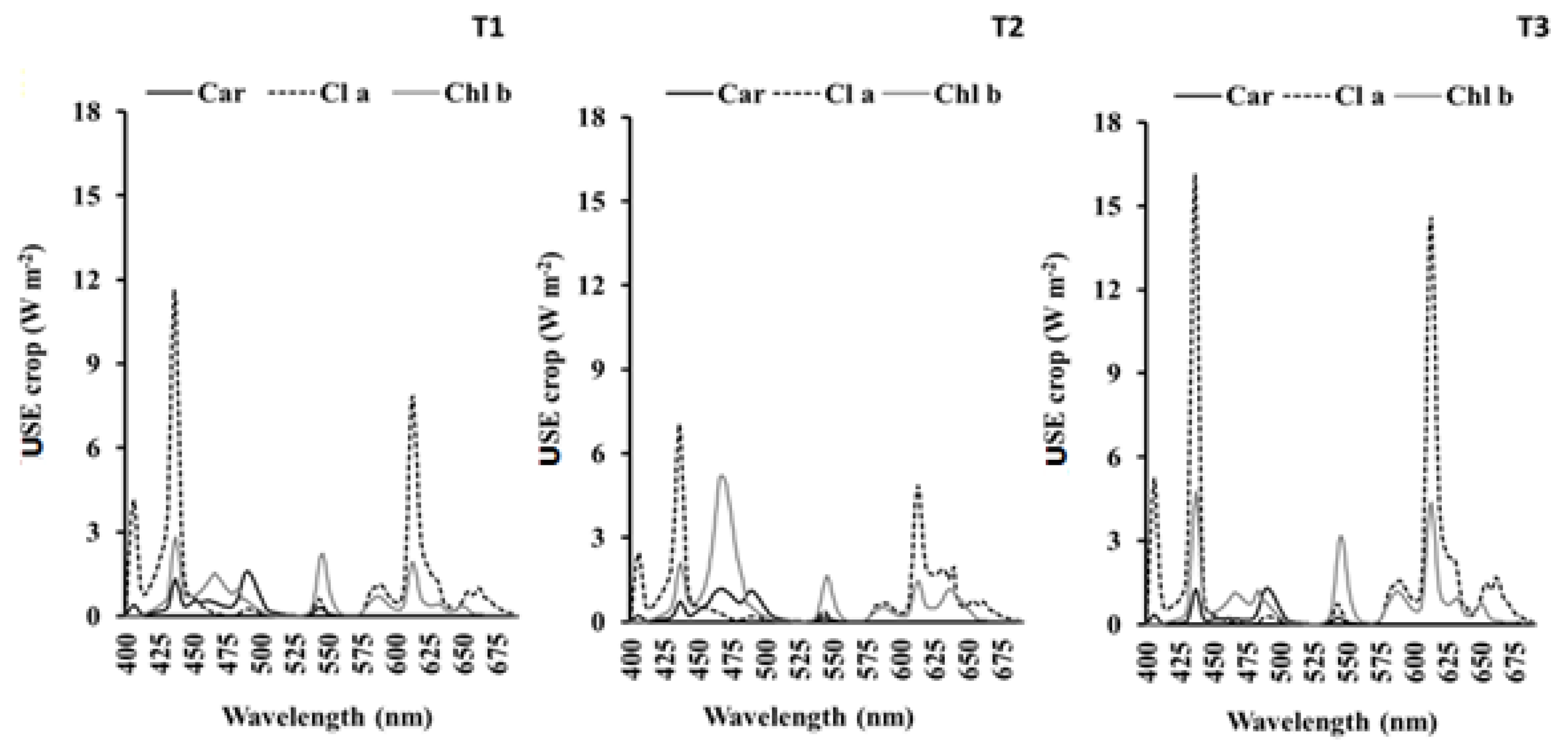
Considering that all leaves have received the same irradiance, the theoretical USEpigment for each light treatment was calculated using the following equation:
where:
USEpigment (car, chl a, chl b): use of spectral energy absorbed by each pigment per plants expressed in W·m−2;
LW: leaf fresh weight per plant expressed in g plant−1; and
c: pigment concentration in leaves, expressed in mg·g−1·FW.
Considering the three photosynthetic pigments, the theoretical IUSE by crop was calculated as the summation of the USE of each pigment:
2.8. Statistical Analysis
The experiment had a completely randomized block design, and the values obtained for each plant and each variable were considered as independent replicates. The data were analyzed through one-way analysis of variance (ANOVA) and least significant difference (LSD) tests (p < 0.05) in order to assess the differences between treatments. To relate the IUSE and biomass (TDW), a simple regression was done, and this regression was compared with the classical simple regression between PAR and biomass (TDW). All the statistical analyses were carried out with Statgraphic Plus for Windows (version 5.1.).
4. Discussion
The light spectrum greatly influences plant development, morphology, and physiology [
26]. In our experiment, growth under different types of lighting had different effects on the biomass and the biochemical parameters of
D. maculata plants. The highest value of root dry weight in plants grown under TLD lamps and BR-LEDs (T2) may be ascribed to the lower value of red light of this type of lamp. Different results were observed by Randall and Lopez [
27], who noted an increase of root dry weight in
Celosia and
Impatiens plants under increasing red light treatment. After 8 weeks of exposure to different light treatments, the highest value of stem dry weight in plants grown under TLD lamps (T1) may be ascribed to this treatment having the highest value of blue light. Similar results were reported by Terfa and co-workers [
28,
29], who noted an increase of stem dry weight in rose plants under blue light treatment. Concerning leaf dry weight, plants grown under high-efficiency fluorescent TL5 showed the highest value, and this could be due to the high value of red light emitted by these lamps. These results are at odds with the data obtained by Zheng and Van Labeke [
30], who reported that the leaf dry weight in
Ficus benjamina and
Sinningia speciosa plants was significantly lower under red light.
The highest value of total plant dry weight under TLD lamps (T1) and high-efficiency fluorescent TL5 (T3) may be ascribed to the different effects induced by the radiation emitted by these lamps. In plants grown under TLD lamps (T1), the increase in total plant dry weight could be related to the highest values of PAR, whereas in plants grown under high-efficiency fluorescent TL5 (T3), it could be attributed to the highest values of red light. The increase in total plant dry weight under high levels of photosynthetically active radiation was in line with the results obtained by Currey and Lopez [
31], who also noted an increase in total plant dry weight in ornamental species such as
Impatiens and
Petunia. With respect to the effects of red light on total plant dry weight in ornamental species, Wollaeger and Runkle [
32] reported that the plant dry weight was greatest for
Salvia and
Petunia under these light conditions. These results could be explained by the fact that increasing red light enhanced leaf area and consequently resulted in a greater photosynthetic capacity, which led to an increase in total plant dry weight, although the values of PAR in high-efficiency TL5 fluorescents (T3) was lower compared to TLD lamps (T1). The different trends in biomass partitioning in plants grown under three different light treatments suggested preferential biomass accumulation between different sink tissues with respect to the light characterization. Moreover, in our experiment, water content in all the organs studied was not affected by the light treatments. These results suggest that there was no relationship between total radiation and water content, as can be expected, although it is well known that an increase in total radiation involves a high transpiration and, consequently, a decrease in the water content in TLD lamps (T1) and high-efficiency fluorescent TL5 (T3)) [
33]. In addition, it is necessary to highlight that the highest B/R ratio in TLD lamps and BR-LEDs (T2) may induce a high stomatal aperture according to Inoue and Kinoshita [
34], promoting the same rate of transpiration as in the other light treatments.
Mills and Jones [
35] proposed the following as a leaf nutrient reference in
D. maculata: N (25–40 mg·g
−1·DW), P (2.5–8.5 mg·g
−1·DW), and K (17–65 mg·g
−1·DW). The leaf nutrient reference for the variety “Camille” proposed by Segura et al. [
36] was the following: N (30 mg·g
−1·DW), P (12 mg·g
−1·DW), and K (50 mg·g
−1·DW). In our experiment, the N concentration was lower than the references cited, which can be related with the specific characteristics of the variety. Phosphorus concentration was inside the range proposed by Mills and Jones [
35] and lower than that of Segura et al. [
36]. In the case of K concentration, this was inside the ranges proposed by both references.
There are a limited number of studies on mineral nutrient uptake by crops as affected by different light treatments. The results obtained in this experiment showed that P concentrations in the different organs of the plants under the three light treatments were similar. Similarly, Evans et al. [
37] reported no differences in P concentrations in
Betula cordifolia and
Abies balsamea under different light conditions. Nevertheless, Kopsell and Sams [
38] reported an increase in P in shoot tissue in sprouting broccoli microgreens grown under blue light exposure. The N and K concentrations in roots and leaves, respectively, were also similar. However, N concentration in stems and leaves, and K concentration in roots and leaves of the plants, displayed different patterns under different light treatments, being the highest under TLD lamps (T1). The highest N concentration in stems and leaves in plants grown under TLD lamps may be ascribed to the highest values of total radiation as reported by Sakuraba and Yanagisawa [
39]. Moreover, the activity of nitrate reductase is influenced by the total radiation, as reported by Fallovo et al. [
40], which involves a decrease in nitrate concentration due to its reduction, which leads to a high level of nitrogen uptake and the consequent high level in leaves. Almansa et al. [
41] also reported an increase in N concentration in tomato seedlings grown under lamps with high blue spectrum values, which could be explained by the high activity of nitrate reductase in plants grown under blue light conditions, as proposed by Azuara and Aparicio [
42]. The highest K concentration in roots and leaves in plants grown under TLD lamps can be also related with the high values of blue light radiated by the lamps. Similarly, Babourina et al. [
43] reported that Arabidopsis seedlings grown under blue light conditions showed an increase in K concentration due to the changes in guard cell membrane transport activity.
Although the effects of light quality on proline biosynthesis are not well documented, it is well known that proline is a compound that ameliorates the damage caused by Reactive Oxygen Species (ROS) formation under different light conditions [
44]. In our experiment, the highest concentration of proline in leaves was observed in plants grown under high levels of blue light. Similar results were reported by Zheng and Van Labeke [
45] for
Chrysanthemum plants grown under blue compared with the other light qualities. Auxin synthesis is modified by intensity and quality spectrum light [
46]. In our experiment, the highest IAA concentration in plants grown under TLD lamps may be due to the high values of blue light discharged by the lamps. These results agree with the findings reported by Fukuda et al. [
47], who noted an increase in IAA concentration in
Petunia hybrida plants grown under blue light conditions. However, cowpea and sunflower plants showed an increase in the endogenous level of IAA under FR light treatment and reduced by a high R:FR ratio [
48,
49]. The results obtained by these researchers can be ascribed to the process of photoconversion of phytochrome (phyB) into the inactive (Pr) form and the low phyA levels due to the R-enriched light environment before the shift to low R/FR-ratio light since phyA and phyB are both repressors of auxin-regulated gene transcription [
50].
Light quality participates in carbohydrate metabolism and consequently in the growth of plants [
51]. In this experiment, total soluble sugars and starch concentrations in leaves were higher in plants grown under high-efficiency TL5 fluorescents, which can be related to the high values of red in the lamps. These results were in line with earlier investigations which showed that the increase in sucrose and starch content under R treatment is dependent on the reduction in the translocation rate of photosynthetic products out of the leaves [
52,
53]. Moreover, it is necessary to point out that the starch concentration was higher than the total soluble sugar concentration, which could be due to the effects of red light, as it may retard the translocation of photosynthetic products and thereby increase the accumulation of starch [
11].
Changes in irradiance wavelengths can cause changes in chlorophyll biosynthesis [
54]. In our experiment, the lowest chlorophyll concentrations were found in plants grown under TLD lamps. Contradictory results were reported by Randall and Lopez [
55], who noted an increase in chlorophyll concentration as the amount of blue light increased for several bedding ornamental species, such as
Antirrhinum majus,
Pelargonium hortorum, and
Salvia splendens. Similar results were obtained for roses grown under blue light treatments by Bergstrand et al. [
56]. No variations in carotenoid concentrations under different light treatments were reported in this experiment. These results suggest that carotenoid concentrations did not appear to be affected by the wavelengths in the light environment, coinciding with the results reported by Zheng and Labeke [
30], who noted that carotenoid concentrations in
Cordyline australis and
F. benjamina did not vary under different light treatments.
The pigment concentration plays a crucial role in the potential photosynthesis rate as well as plant growth [
57]. The majority of harvesting light energy is carried out through chlorophylls, which are mainly absorbed in the blue–red part of the spectrum [
58]. Harvesting of light energy by carotenoids is lower, since they are less efficient at excitation energy transfer than chlorophylls [
59].
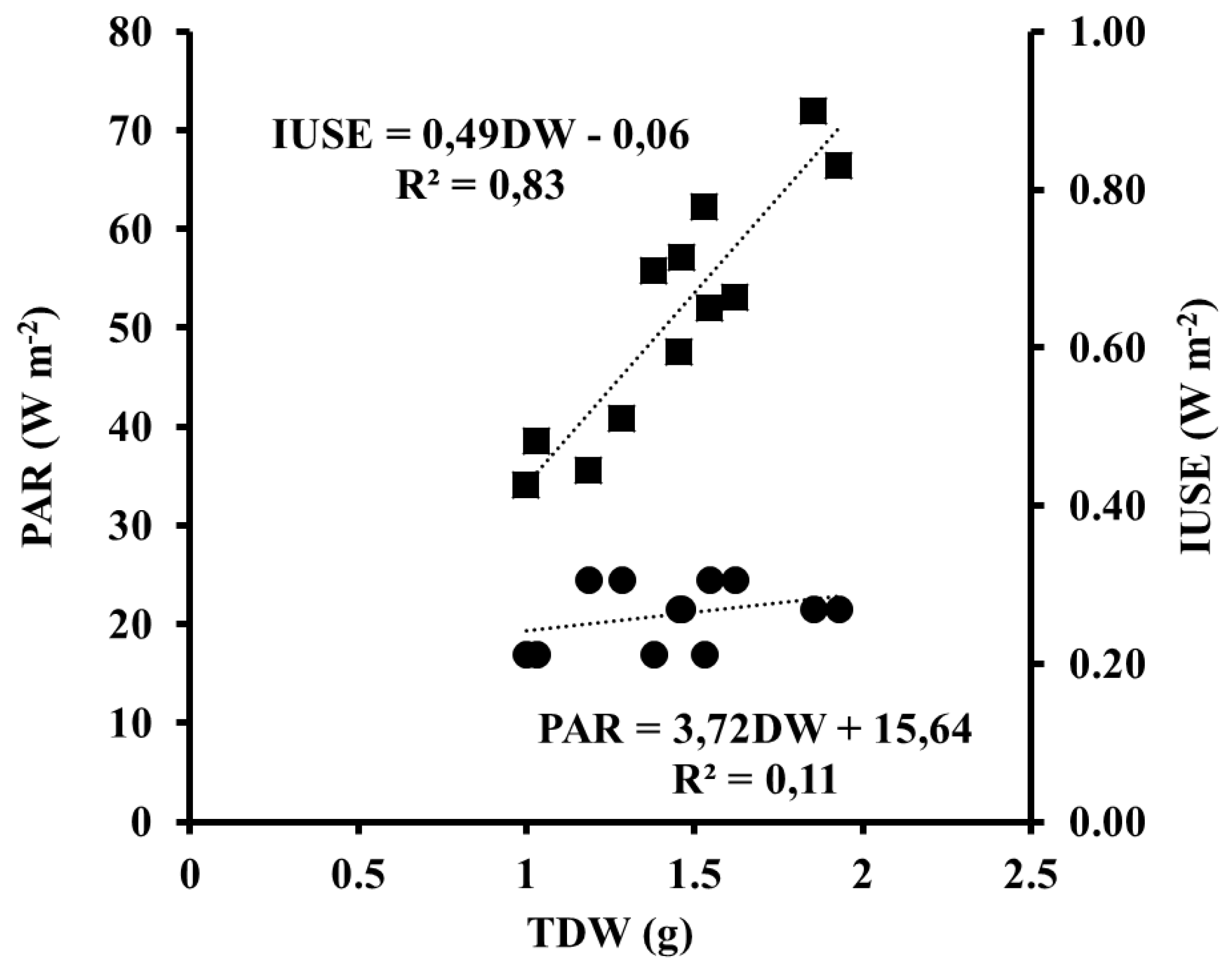
In our experiment we have assessed the IUSE by the spectral energy emitted by the lamp, radiation absorption by the pigments, and the photosynthetic pigment concentration in plants in order to evaluate the energy savings by the plant. The results obtained reporting a higher correlation between IUSE and DW. Surveying preceding works, there are no references to this methodology. In our approach, we have considered that the main function of chlorophyll in plants is the absorbance of light and transference through plants during photosynthesis, as well as the joint pigment capacity for harvesting radiation [
60].

 and
and 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}