Community-Level Physiological Profiles of Microorganisms from Different Types of Soil That Are Characteristic to Poland—A Long-Term Microplot Experiment



Abstract
:1. Introduction
2. Materials and Methods
2.1. Microplot Experiment
2.2. Soil Sampling
2.3. pH Measurements
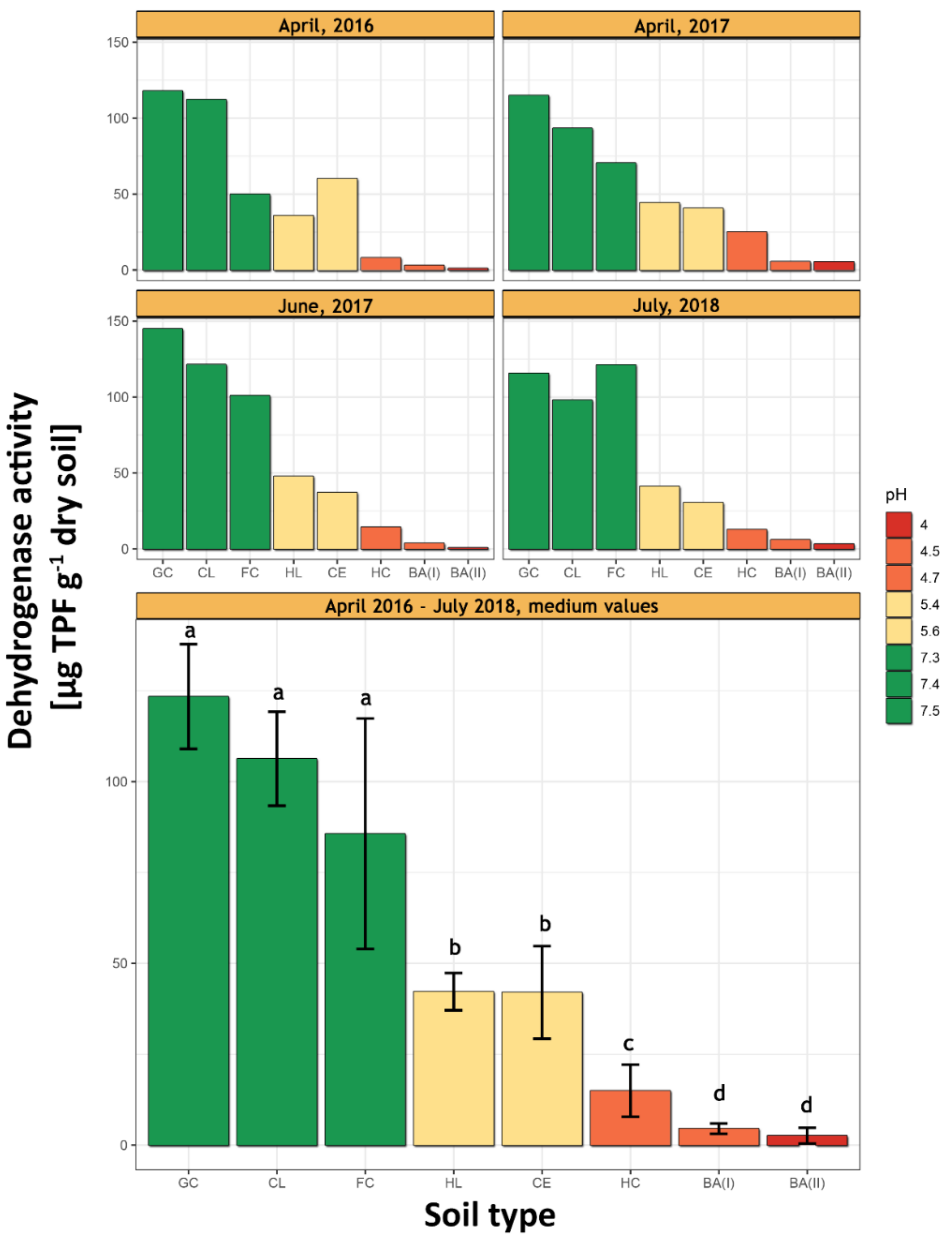
2.4. Dehydrogenases Activity Measurements
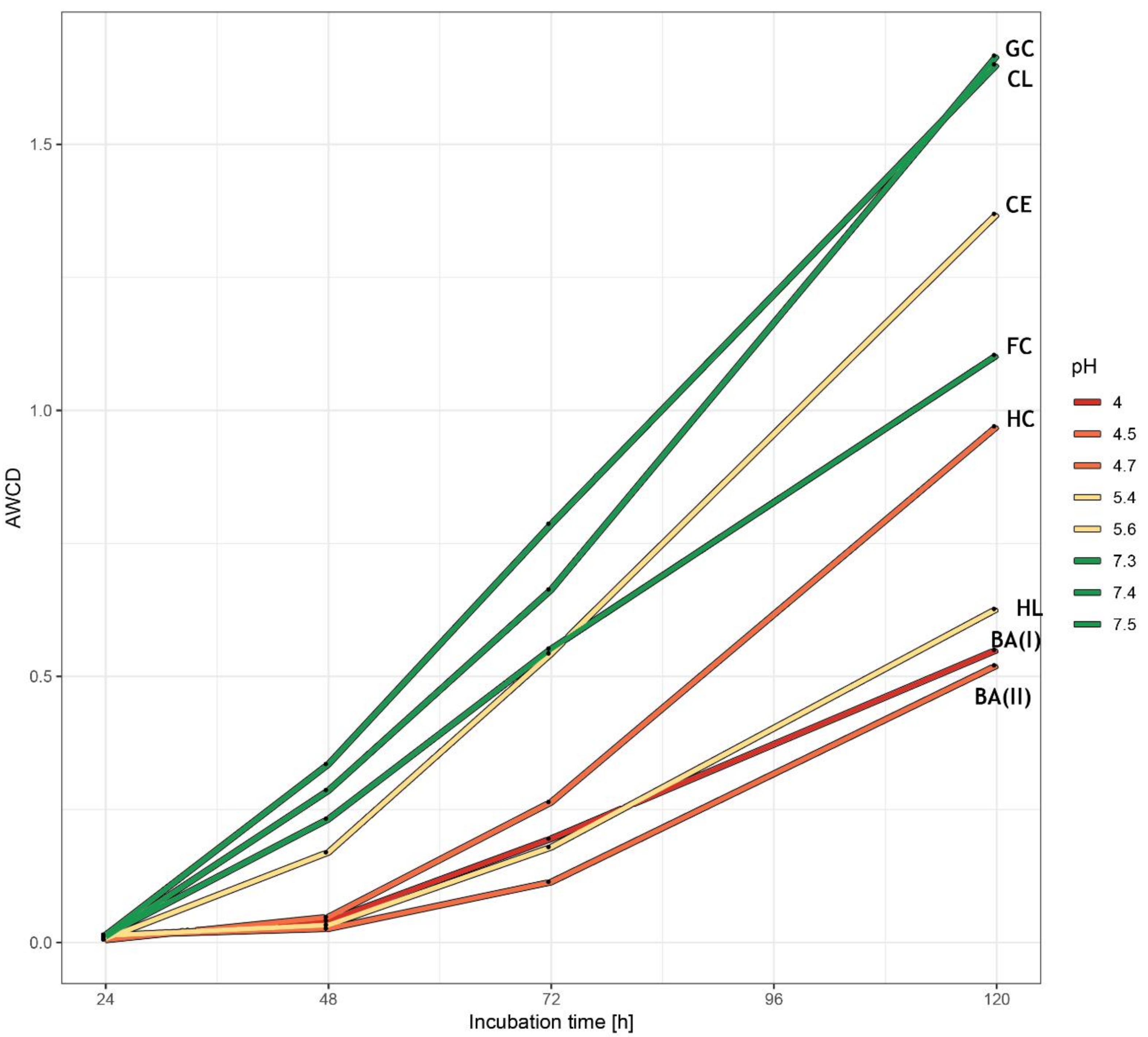
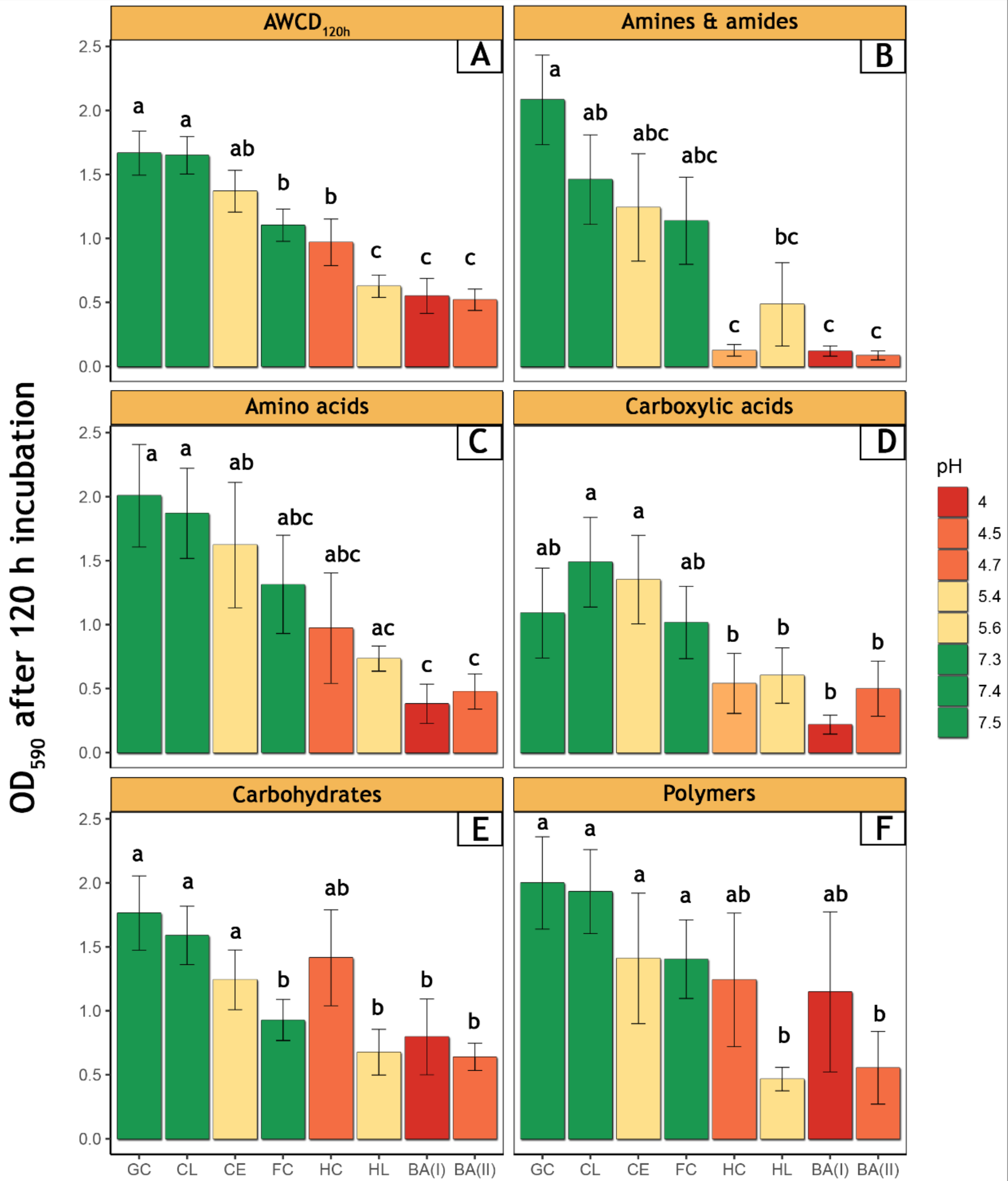
2.5. Community-Level Physiological Profiles of Microorganisms—EcoPlate™
2.6. Statistical Analyses
3. Results
3.1. pH and the Dehydrogenases Activity
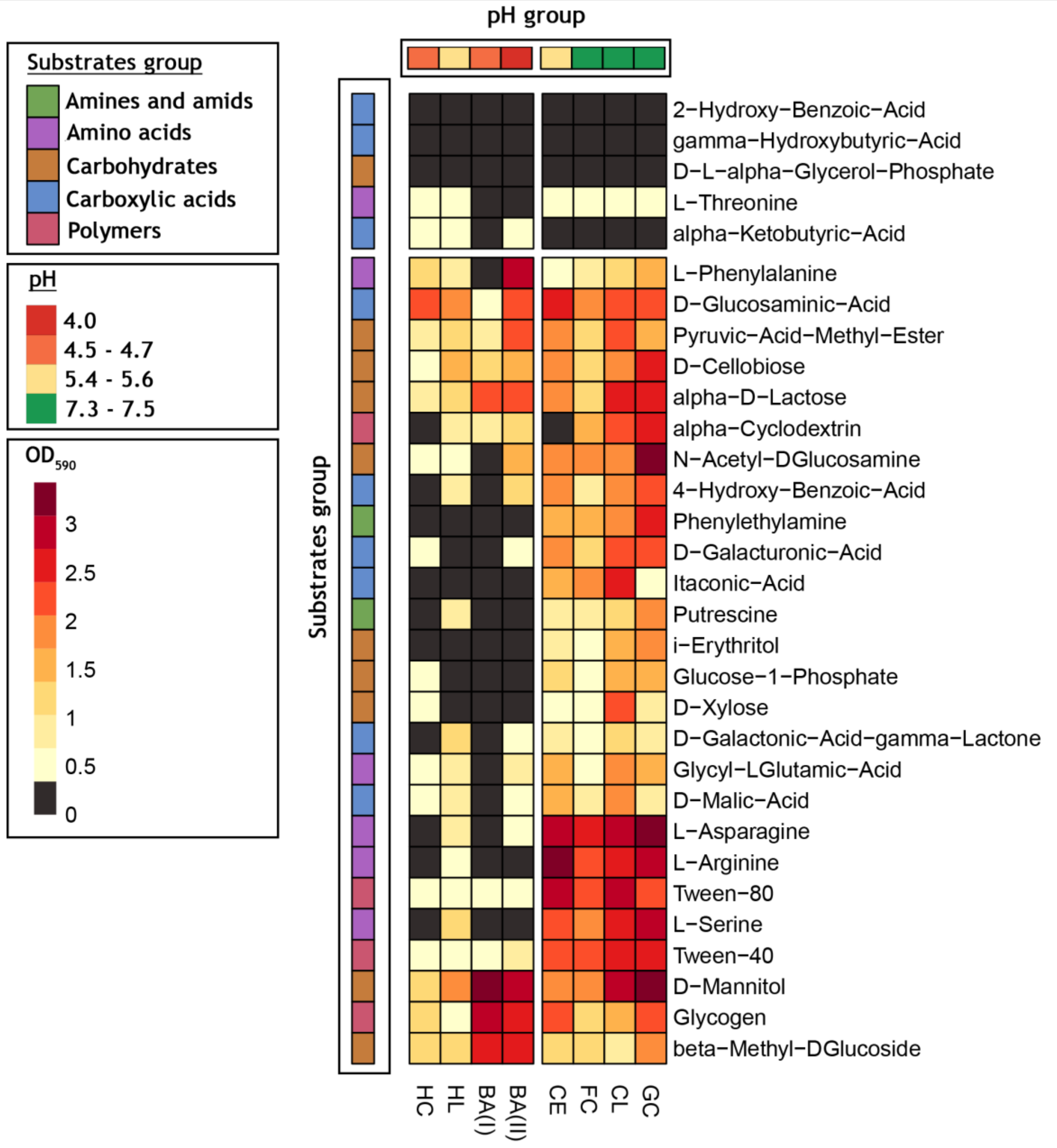
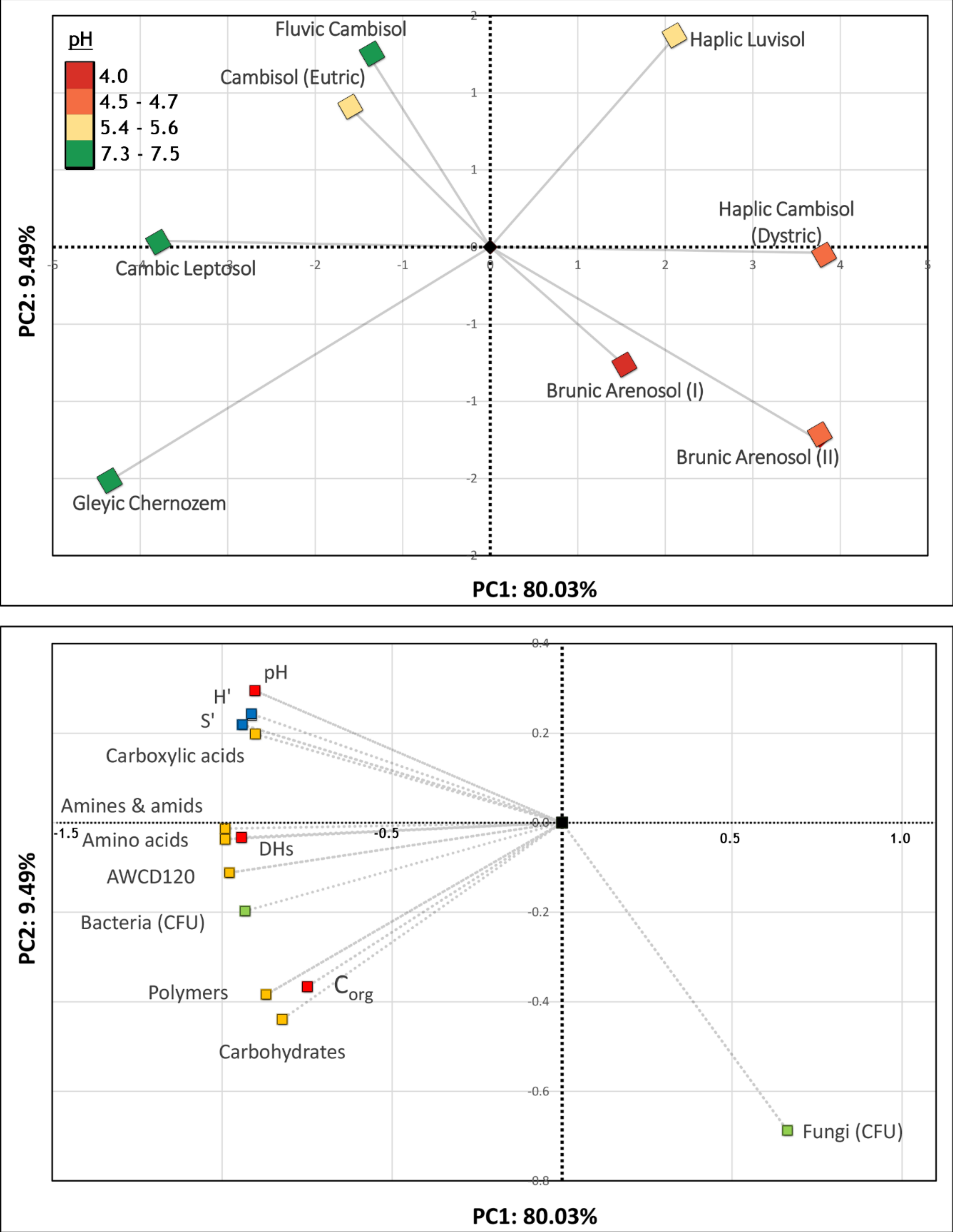
3.2. Bacterial Community-Level Physiological Profiles—EcoPlate™
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EcoPlate™ Substrates | Well Color Development (OD590) | σ2 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| BA(II) | CL | FC | HL | GC | BA(I) | CE | HC | ||
| β- methyl- D- glucoside | 2.71 (±0.28) | 1.00 (±0.37) | 1.26 (±0.67) | 1.23 (±0.37) | 1.93 (±0.66) | 2.68 (±0.26) | 1.30 (±1.23) | 1.10 (±0.78) | 0.49 |
| D- galactonic acid γ- lactone | 0.56 (±0.50) | 1.28 (±0.21) | 0.61 (±0.18) | 1.11 (±0.32) | 0.83 (±0.20) | 0.31 (±0.50) | 0.98 (±0.33) | 0.05 (±0.01) | 0.17 |
| L- arginine | 0.25 (±0.46) | 2.59 (±0.88) | 2.23 (±0.89) | 0.36 (±0.41) | 2.80 (±1.29) | 0.15 (±0.08) | 3.12 (±0.97) | 0.23 (±0.20) | 1.77 |
| Pyruvic acid methyl ester | 2.36 (±0.50) | 2.17 (±0.80) | 1.11 (±0.34) | 1.35 (±0.30) | 1.51 (±0.58) | 0.79 (±0.76) | 1.95 (±0.61) | 0.96 (±0.42) | 0.34 |
| D- xylose | 0.23 (±0.09) | 2.28 (±0.54) | 0.65 (±0.09) | 0.22 (±0.08) | 0.84 (±0.45) | 0.29 (±0.03) | 0.60 (±0.62) | 0.64 (±0.53) | 0.45 |
| D- galacturonic acid | 0.40 (±0.09) | 2.23 (±0.13) | 1.03 (±0.27) | 0.32 (±0.17) | 2.37 (±0.75) | 0.21 (±0.05) | 1.90 (±0.95) | 0.44 (±0.28) | 0.84 |
| L- asparagine | 0.43 (±0.13) | 3.04 (±0.03) | 2.74 (±0.64) | 0.86 (±0.83) | 3.43 (±0.25) | 0.21 (±0.14) | 3.06 (±0.18) | 0.27 (±0.08) | 2.04 |
| Tween 40 | 0.93 (±0.79) | 2.62 (±0.53) | 2.09 (±0.67) | 0.51 (±0.18) | 2.43 (±0.39) | 0.51 (±0.21) | 2.33 (±0.94) | 0.41 (±0.04) | 0.95 |
| i- erythritol | 0.29 (±0.10) | 1.52 (±0.64) | 0.59 (±0.10) | 0.30 (±0.08) | 1.72 (±0.25) | 0.27 (±0.09) | 0.83 (±0.19) | 0.19 (±0.12) | 0.36 |
| 2- hydroxy benzoic acid | 0.07 (±0.04) | 0.25 (±0.19) | 0.03 (±0.01) | 0.15 (±0.04) | 0.04 (±0.02) | 0.07 (±0.02) | 0.17 (±0.07) | 0.11 (±0.03) | 0.01 |
| L- phenylalanine | 3.03 (±0.37) | 1.36 (±0.61) | 0.72 (±0.62) | 0.70 (±0.14) | 1.50 (±0.45) | 0.19 (±0.19) | 0.55 (±0.33) | 1.14 (±1.70) | 0.77 |
| Tween 80 | 0.40 (±1.40) | 3.00 (±0.19) | 2.17 (±0.30) | 0.44 (±0.31) | 2.40 (±0.19) | 0.51 (±1.47) | 2.81 (±0.71) | 0.49 (±0.45) | 1.36 |
| D- mannitol | 2.78 (±0.09) | 2.92 (±0.19) | 2.05 (±0.38) | 1.81 (±0.89) | 3.12 (±0.38) | 3.19 (±0.41) | 1.88 (±0.19) | 1.10 (±0.71) | 0.57 |
| 4- hydroxy benzoic acid | 1.10 (±1.60) | 1.99 (±0.49) | 0.99 (±0.65) | 0.80 (±1.05) | 2.19 (±0.89) | 0.11 (±0.05) | 1.73 (±0.95) | 0.03 (±0.03) | 0.66 |
| L- serine | 0.34 (±0.08) | 2.70 (±0.62) | 1.77 (±0.45) | 1.05 (±0.92) | 2.77 (±1.45) | 0.17 (±0.09) | 2.34 (±0.25) | 0.33 (±0.20) | 1.21 |
| α- cyclodextrin | 1.04 (±1.28) | 2.28 (±0.29) | 1.65 (±0.66) | 0.69 (±1.18) | 2.51 (±0.46) | 0.95 (±1.59) | 0.16 (±0.27) | 0.03 (±0.02) | 0.84 |
| N- acetyl-D- glucosamine | 1.60 (±1.37) | 1.84 (±0.12) | 1.84 (±0.41) | 0.39 (±0.35) | 3.22 (±0.06) | 0.27 (±0.10) | 2.01 (±0.29) | 0.37 (±0.22) | 1.06 |
| γ- hydroxybutyric acid | 0.03 (±0.04) | 0.08 (±0.07) | 0.10 (±0.12) | 0.05 (±0.12) | 0.12 (±0.16) | 0.04 (±0.03) | 0.04 (±0.07) | 0.07 (±0.08) | 0.00 |
| L- threonine | 0.29 (±0.15) | 0.62 (±0.08) | 0.51 (±0.39) | 0.63 (±0.35) | 0.68 (±0.41) | 0.30 (±0.33) | 0.40 (±0.03) | 0.48 (±0.29) | 0.02 |
| Glycogen | 2.72 (±0.27) | 1.70 (±0.70) | 1.22 (±0.38) | 0.42 (±0.47) | 2.11 (±0.66) | 2.96 (±0.46) | 2.12 (±0.71) | 1.36 (±1.51) | 0.69 |
| D- glucosaminic acid | 2.32 (±0.95) | 2.09 (±0.45) | 1.92 (±0.41) | 2.04 (±0.73) | 2.23 (±0.74) | 0.60 (±0.79) | 2.57 (±0.49) | 2.11 (±1.17) | 0.35 |
| Itaconic acid | 0.04 (±1.73) | 2.41 (±0.58) | 1.91 (±0.63) | 0.03 (±0.55) | 0.43 (±1.17) | 0.01 (±1.41) | 1.68 (±1.22) | 0.05 (±0.16) | 1.02 |
| Glycyl-L- glutamic acid | 0.71 (±0.90) | 1.76 (±0.27) | 0.61 (±0.21) | 0.78 (±0.62) | 1.62 (±0.67) | 0.30 (±0.10) | 1.44 (±0.61) | 0.42 (±0.08) | 0.32 |
| D- cellobiose | 1.56 (±0.86) | 1.75 (±0.59) | 1.17 (±0.27) | 1.42 (±1.00) | 2.65 (±0.10) | 1.31 (±0.59) | 1.98 (±0.10) | 0.51 (±0.45) | 0.39 |
| Glucose-1- phosphate | 0.02 (±0.03) | 1.38 (±0.05) | 0.49 (±0.21) | 0.00 (±0.00) | 1.71 (±0.40) | 0.02 (±0.01) | 1.14 (±0.16) | 0.57 (±0.99) | 0.45 |
| α- ketobutyric acid | 0.56 (±0.24) | 0.34 (±0.08) | 0.28 (±0.07) | 0.36 (±0.08) | 0.14 (±0.07) | 0.02 (±0.00) | 0.32 (±0.01) | 0.46 (±0.12) | 0.03 |
| Phenylethylamine | 0.17 (±0.23) | 1.81 (±0.34) | 1.48 (±0.65) | 0.16 (±0.10) | 2.43 (±1.66) | 0.08 (±0.05) | 1.66 (±1.02) | 0.05 (±0.05) | 0.93 |
| α-D- lactose | 2.32 (±0.61) | 2.57 (±0.27) | 1.35 (±0.05) | 1.15 (±0.37) | 2.54 (±0.94) | 2.11 (±0.27) | 2.05 (±0.72) | 0.70 (±0.28) | 0.48 |
| D,L- α-glycerol phosphate | 0.02 (±0.01) | 0.04 (±0.06) | 0.10 (±0.14) | 0.02 (±0.03) | 0.02 (±0.00) | 0.03 (±0.01) | 0.02 (±0.03) | 0.24 (±0.39) | 0.01 |
| D- malic acid | 0.35 (±0.06) | 1.91 (±0.24) | 0.99 (±0.80) | 0.83 (±0.56) | 1.02 (±0.65) | 0.18 (±0.10) | 1.70 (±0.83) | 0.56 (±0.71) | 0.37 |
| Putrescine | 0.08 (±0.03) | 1.11 (±0.14) | 0.80 (±0.16) | 0.81 (±0.85) | 1.73 (±0.18) | 0.16 (±0.01) | 0.82 (±0.46) | 0.12 (±0.04) | 0.33 |
References
- Doran, J.W.; Zeiss, M.R. Soil health and sustainability: managing the biotic component of soil quality. Appl. Soil Ecol. 2000, 15, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef] [Green Version]
- Lehman, R.; Cambardella, C.; Stott, D.; Acosta-Martinez, V.; Manter, D.; Buyer, J.; Maul, J.; Smith, J.; Collins, H.; Halvorson, J.; et al. Understanding and Enhancing Soil Biological Health: The Solution for Reversing Soil Degradation. Sustainability 2015, 7, 988–1027. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, A.C.; Smith, K.L. Soil microbial diversity and the sustainability of agricultural soils. Plant Soil 1995, 170, 75–86. [Google Scholar] [CrossRef]
- Sessitsch, A.; Weilharter, A.; Gerzabek, M.H.; Kirchmann, H.; Kandeler, E. Microbial population structures in soil particle size fractions of a long-term fertilizer field experiment. Appl. Environ. Microbiol. 2001, 67, 4215–4224. [Google Scholar] [CrossRef] [PubMed]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Latour, X.; Philippot, L.; Corberand, T.; Lemanceau, P. The establishment of an introduced community of fluorescent pseudomonads in the soil and in the rhizosphere is affected by the soil type. FEMS Microbiol. Ecol. 1999, 30, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Chau, J.F.; Bagtzoglou, A.C.; Willig, M.R. The Effect of Soil Texture on Richness and Diversity of Bacterial Communities. Environ. Forensics 2011, 12, 333–341. [Google Scholar] [CrossRef]
- Schreiter, S.; Ding, G.C.; Heuer, H.; Neumann, G.; Sandmann, M.; Grosch, R.; Kropf, S.; Smalla, K. Effect of the soil type on the microbiome in the rhizosphere of field-grown lettuce. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Girvan, M.S.; Bullimore, J.; Pretty, J.N.; Osborn, A.M.; Ball, A.S. Soil Type Is the Primary Determinant of the Composition of the Total and Active Bacterial Communities in Arable Soils. Appl. Environ. Microbiol. 2003, 69, 1800–1809. [Google Scholar] [CrossRef] [Green Version]
- Langille, M.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.; Clemente, J.; Burkepile, D.; Vega Thurber, R.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garland, J.L.; Mills, A.L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [PubMed]
- Kuźniar, A.; Banach, A.; Stępniewska, Z.; Frąc, M.; Oszust, K.; Gryta, A.; Kłos, M.; Wolińska, A. Community-level physiological profiles of microorganisms inhabiting soil contaminated with heavy metals. Int. Agrophysics 2018, 32, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Lv, T.; Zhang, Y.; Carvalho, P.N.; Zhang, L.; Button, M.; Arias, C.A.; Weber, K.P.; Brix, H. Microbial community metabolic function in constructed wetland mesocosms treating the pesticides imazalil and tebuconazole. Ecol. Eng. 2017, 98, 378–387. [Google Scholar] [CrossRef]
- Chou, Y.M.; Shen, F.T.; Chiang, S.C.; Chang, C.M. Functional diversity and dominant populations of bacteria in banana plantation soils as influenced by long-term organic and conventional farming. Appl. Soil Ecol. 2017, 110, 21–33. [Google Scholar] [CrossRef]
- Furtak, K.; Gawryjołek, G.; Gajda, A.M.; Gałązka, A. Effects of maize and winter wheat grown under different cultivation techniques on biological activity of soil. Plant Soil Environ. 2017, 63, 449–454. [Google Scholar] [Green Version]
- Li, G.; Kim, S.; Park, M.; Son, Y. Short-Term Effects of Experimental Warming and Precipitation Manipulation on Soil Microbial Biomass C and N, Community Substrate Utilization Patterns and Community Composition. Pedosphere 2017, 27, 714–724. [Google Scholar] [CrossRef]
- Ros, M.; Goberna, M.; Pascual, J.A.; Klammer, S.; Insam, H. 16S rDNA analysis reveals low microbial diversity in community level physiological profile assays. J. Microbiol. Methods 2008, 72, 221–226. [Google Scholar] [CrossRef]
- Lladó, S.; Baldrian, P. Community-level physiological profiling analyses show potential to identify the copiotrophic bacteria present in soil environments. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Huang, X.P.; Mo, C.H.; Yu, J.; Zhao, H.M.; Meng, C.; Li, Y.W.; Li, H.; Cai, Q.Y.; Wong, M.H. Variations in microbial community and ciprofloxacin removal in rhizospheric soils between two cultivars of Brassica parachinensis L. Sci. Total Environ. 2017, 603–604, 66–76. [Google Scholar] [CrossRef]
- Jin, X.; Ma, Y.; Kong, Z.; Kou, W.; Wu, L. The Variation of Sediment Bacterial Community in Response to Anthropogenic Disturbances of Poyang Lake, China. Wetlands 2017, 1–11. [Google Scholar] [CrossRef]
- Narayanamurthy, V.; Sweetnam, J.M.; Denner, D.R.; Chen, L.W.; Naureckas, E.T.; Laxman, B.; White, S.R. The metabolic footprint of the airway bacterial community in cystic fibrosis. Microbiome 2017, 5, 67. [Google Scholar] [CrossRef] [PubMed]
- Pino-Otín, M.R.; Muñiz, S.; Val, J.; Navarro, E. Effects of 18 pharmaceuticals on the physiological diversity of edaphic microorganisms. Sci. Total Environ. 2017, 595, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Cardozo, F.M., Jr.; Carneiro, R.F.V.; Rocha, S.M.B.; Nunes, L.A.P.L.; dos Santos, V.M.; de Lima Feitoza, L.; de Araújo, A.S.F. The Impact of Pasture Systems on Soil Microbial Biomass and Community-level Physiological Profiles. L. Degrad. Dev. 2018, 29, 284–291. [Google Scholar] [CrossRef]
- Gałązka, A.; Gawryjołek, K.; Grządziel, J.; Frąc, M.; Księżak, J. Microbial community diversity and the interaction of soil under maize growth in different cultivation techniques. Plant Soil Environ. 2017, 63, 264–270. [Google Scholar] [Green Version]
- Siebielec, G.; Siebielec, S.; Podolska, G. Comparison of microbial and chemical characteristics of soil types after over 100 years of cereal production. Polish J. Agron. 2015, 23, 88–100. [Google Scholar]
- Wallace, R.H.; Lochhead, A.G. Qualitative Studies of Soil Microorganisms: IX. Amino acid requirements of rhisosphere bacteria. Can. J. Res. 1950, 28c, 1–6. [Google Scholar] [CrossRef]
- Martin, J.P. Use of acid, rose bengal, and streptomycin in the plate method for estimating soil fungi. Soil Sci. 1950. [Google Scholar] [CrossRef]
- Casida, L.E., Jr.; Klein, D.A.; Santoro, T. Soil dehydrogenase activity. Soil Sci. 1964, 98, 371–376. [Google Scholar] [CrossRef]
- Insam, H. A New Set of Substrates Proposed for Community Characterization in Environmental Samples. In Microbial Communities; Springer: Berlin/Heidelberg, Germany, 1997; pp. 259–260. [Google Scholar]
- Weber, K.P.; Legge, R.L. Community-Level Physiological Profiling. In Bioremediation: Methods and Protocols; Cummings, S.P., Ed.; Humana Press: Totowa, NJ, USA, 2010; ISBN 978-1-60761-439-5. [Google Scholar]
- Preston-Mafham, J.; Boddy, L.; Randerson, P.F. Analysis of microbial community functional diversity using sole-carbon-source utilisation profiles—A critique. FEMS Microbiol. Ecol. 2002, 42, 1–4. [Google Scholar]
- Garland, J.L. Analysis and interpretation of community-level physiological profiles in microbial ecology. FEMS Microbiol. Ecol. 1997, 24, 289–300. [Google Scholar] [CrossRef]
- Hill, T.C.J.; Walsh, K.A.; Harris, J.A.; Moffett, B.F. Using ecological diversity measures with bacterial communities. FEMS Microbiol. Ecol. 2003, 43, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zak, J.; Willig, M.; Moorhead, D.; Wildman, H. Functional diversity of microbial communities: A quantitative approach. Soil Biol. Biochem. 1994, 26, 1101–1108. [Google Scholar] [CrossRef]
- Jalowiecki, L.; Chojniak, J.M.; Dorgeloh, E.; Hegedusova, B.; Ejhed, H.; Magner, J.; Plaza, G.A. Microbial Community Profiles in Wastewaters from Onsite Wastewater Treatment Systems Technology. PLoS ONE 2016, 11, e0147725. [Google Scholar] [CrossRef] [PubMed]
- Grządziel, J. Functional redundancy of soil microbiota – Does more always mean better? Polish J. Soil Sci. 2017, 50. [Google Scholar] [CrossRef]
- Bastida, F.; Kandeler, E.; Moreno, J.L.; Ros, M.; Garcı, C. Application of fresh and composted organic wastes modifies structure, size and activity of soil microbial community under semiarid climate. Appl. Soil Ecol. 2008, 40, 318–329. [Google Scholar] [CrossRef]
- Gałązka, A.; Gawryjołek, K.; Grządziel, J.; Księżak, J. Effect of different agricultural management practices on soil biological parameters including glomalin fraction. Plant Soil Environ. 2017, 63, 300–306. [Google Scholar] [Green Version]
- Gruba, P.; Błońska, E.; Lasota, J.; Gruba, P. Effect of temperate forest tree species on soil dehydrogenase and urease activities in relation to other properties of soil derived from loess and glaciofluvial sand. Ecol. Res. 2016, 31, 655–664. [Google Scholar] [Green Version]
- Wolińska, A.; Stępniewska, Z. Dehydrogenase Activity in the Soil Environment. In Dehydrogenases; Canuto, R.A., Ed.; IntechOpen: Rijeka, Croatia, 2012. [Google Scholar] [Green Version]
- Januszek, K.; Błońska, E.; Długa, J.; Socha, J. Dehydrogenase activity of forest soils depends on the assay used. Int. Agrophysics 2015, 29, 47–59. [Google Scholar] [CrossRef] [Green Version]
- Czaban, J.; Wróblewska, B.; Niedźwiecki, J.; Sułek, A. Relationships between numbers of microbial communities in Polish agricultural soils and properties of these soils, paying special attention to xerophilic/xerotolerant fungi. Polish J. Environ. Stud. 2010, 19, 1171–1183. [Google Scholar]
- Aciego Pietri, J.C.; Brookes, P.C. Relationships between soil pH and microbial properties in a UK arable soil. Soil Biol. Biochem. 2008, 40, 1856–1861. [Google Scholar] [CrossRef]
- Bååth, E.; Arnebrant, K. Growth rate and response of bacterial communities to pH in limed and ash treated forest soils. Soil Biol. Biochem. 1994, 26, 995–1001. [Google Scholar] [CrossRef]
- Bååth, E. Growth rates of bacterial communities in soils at varying pH: A comparison of the thymidine and leucine incorporation techniques. Microb. Ecol. 1998, 36, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Rousk, J.; Brookes, P.C.; Bååth, E. Contrasting Soil pH Effects on Fungal and Bacterial Growth Suggest Functional Redundancy in Carbon Mineralization. Appl. Environ. Microbiol. 2009, 75, 1589–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, T.; Brye, K.; Savin, M.; Lee, J.; Gbur, E. Microbial Carbon Substrate Utilization Differences among High- and Average-Yield Soybean Areas. Agriculture 2017, 7, 48. [Google Scholar] [CrossRef]
- Tautges, N.E.; Sullivan, T.S.; Reardon, C.L.; Burke, I.C. Soil microbial diversity and activity linked to crop yield and quality in a dryland organic wheat production system. Appl. Soil Ecol. 2016, 108, 258–268. [Google Scholar] [CrossRef]
- Soman, C.; Li, D.; Wander, M.M.; Kent, A.D. Long-term fertilizer and crop-rotation treatments differentially affect soil bacterial community structure. Plant Soil 2017, 413, 145–159. [Google Scholar] [CrossRef]
- Wolinska, A.; Frąc, M.; Oszust, K.; Szafranek-Nakonieczna, A.; Zielenkiewicz, U.; Stępniewska, Z. Microbial biodiversity of meadows under different modes of land use: catabolic and genetic fingerprinting. World J. Microbiol. Biotechnol. 2017, 33. [Google Scholar] [CrossRef] [Green Version]
- Jesus, E.d.S.; Liang, C.; Quensen, J.F.; Susilawati, E.; Jackson, R.D.; Balser, T.C.; Tiedje, J.M. Influence of corn, switchgrass, and prairie cropping systems on soil microbial communities in the upper Midwest of the United States. GCB Bioenergy 2016, 8, 481–494. [Google Scholar] [CrossRef]
- Marschner, P.; Yang, C.-H.; Lieberei, R.; Crowley, D. Soil and plant specific effects on bacterial community composition in the rhizosphere. Soil Biol. Biochem. 2001, 33, 1437–1445. [Google Scholar] [CrossRef]





| Soil Type | Soil Texture Group 1 | pH (in H2O) | Corg. (g kg−1 soil) | Bacteria 109 CFU g−1 soil [27] | Fungi 105 CFU g−1 soil [28] |
|---|---|---|---|---|---|
| GC | SL | 7.4 | 21.3 | 0.38 ± 0.06 b | 1.16 ± 0.05 bcd |
| FC | SL | 7.5 | 8.20 | 0.28 ± 0.20 a | 0.45 ± 0.29 d |
| CL | SL | 7.3 | 11.00 | 0.36 ± 0.02 bc | 0.70 ± 0.00 cd |
| CE | SL | 5.6 | 7.67 | 0.22 ± 0.01 bcd | 0.46 ± 0.29 d |
| HL | LS | 5.4 | 6.87 | 0.04 ± 0.00 d | 0.88 ± 0.26 bcd |
| BA(I) | LS | 4.7 | 5.27 | 0.23 ± 0.01 bcd | 1.54 ± 0.12 bc |
| HC | S | 4.5 | 7.53 | 0.06 ± 0.01 cd | 1.67 ± 0.42 b |
| BA(II) | LS | 4.0 | 4.57 | 0.06 ± 0.01 cd | 3.04 ± 0.21 a |
| Temperature (°C) | Precipitation (mm) | |||
|---|---|---|---|---|
| Month | 2015 | 2016 | 2015 | 2016 |
| I | 1.3 | –3.3 | 55.3 | 33.0 |
| II | 1.0 | 3.7 | 9.9 | 64.5 |
| III | 5.1 | 4.3 | 45.8 | 53.0 |
| IV | 8.6 | 9.6 | 28.5 | 38.4 |
| V | 13.5 | 15.6 | 111.6 | 72.2 |
| VI | 17.9 | 19.8 | 32.0 | 27.9 |
| VII | 20.4 | 20.1 | 55.4 | 86.6 |
| VIII | 22.6 | 18.7 | 3.6 | 41.9 |
| IX | 15.3 | 15.6 | 126.0 | 20.6 |
| X | 7.0 | 7.7 | 30.0 | 100.2 |
| XI | 5.2 | 3.2 | 47.4 | 44.9 |
| XII | 4.0 | 0.8 | 25.2 | 64.6 |
| Year | 10.2 | 9.7 | 570.7 | 647.8 |
| Index | Formula | Definition | Reference |
|---|---|---|---|
| AWCD | C—reading of the well OD; R—reading of the control well OD; n—the number of substrates on an EcoPlate™ (31) | [12] | |
| Shannon diversity (H′) | pi—proportional color development of the well over the total color development of all wells of a plate; N—the number of substrates on an EcoPlate™ (31) | [34] | |
| Shannon evenness (E) | E = H′/lnS | S—the number of wells with color development (substrate utilization richness) | [35] |
| Richness (S) | Numbers of utilized substrates with OD590 ≥ 0.25 | – | [36] |
| Sample | H′ | E | S |
|---|---|---|---|
| CL | 3.266 (±0.010) b | 0.977 (±0.006) a | 28.333 (±0.577) c |
| GC | 3.198 (±0.019) ab | 0.967 (±0.012) a | 27.333 (±0.577) ac |
| FC | 3.188 (±0.027) ab | 0.964 (±0.015) a | 27.333 (±1.155) ac |
| CE | 3.155 (±0.014) ab | 0.957 (±0.004) a | 27.000 (±0.000) ac |
| HL | 2.971 (±0.072) ad | 0.983 (±0.020) a | 20.667 (±2.887) ab |
| HC | 2.814 (±0.014) cd | 0.935 (±0.027) a | 20.333 (±1.528) ab |
| BA(II) | 2.786 (±0.209) cd | 0.995 (±0.032) a | 17.000 (±5.196) b |
| BA(I) | 2.565 (±0.152) c | 0.992 (±0.056) a | 13.667 (±3.215) b |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grządziel, J.; Furtak, K.; Gałązka, A. Community-Level Physiological Profiles of Microorganisms from Different Types of Soil That Are Characteristic to Poland—A Long-Term Microplot Experiment. Sustainability 2019, 11, 56. https://doi.org/10.3390/su11010056
Grządziel J, Furtak K, Gałązka A. Community-Level Physiological Profiles of Microorganisms from Different Types of Soil That Are Characteristic to Poland—A Long-Term Microplot Experiment. Sustainability. 2019; 11(1):56. https://doi.org/10.3390/su11010056
Chicago/Turabian StyleGrządziel, Jarosław, Karolina Furtak, and Anna Gałązka. 2019. "Community-Level Physiological Profiles of Microorganisms from Different Types of Soil That Are Characteristic to Poland—A Long-Term Microplot Experiment" Sustainability 11, no. 1: 56. https://doi.org/10.3390/su11010056