
Recessive Resistance Derived from Tomato cv. Tyking-Limits Drastically the Spread of Tomato Yellow Leaf Curl Virus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tomato Plants, Virus Isolate, and Whitefly Population
2.2. Virus Inoculation
2.3. Primary and Secondary TYLCV-IL Spread Experiments
2.4. B. tabaci Preference
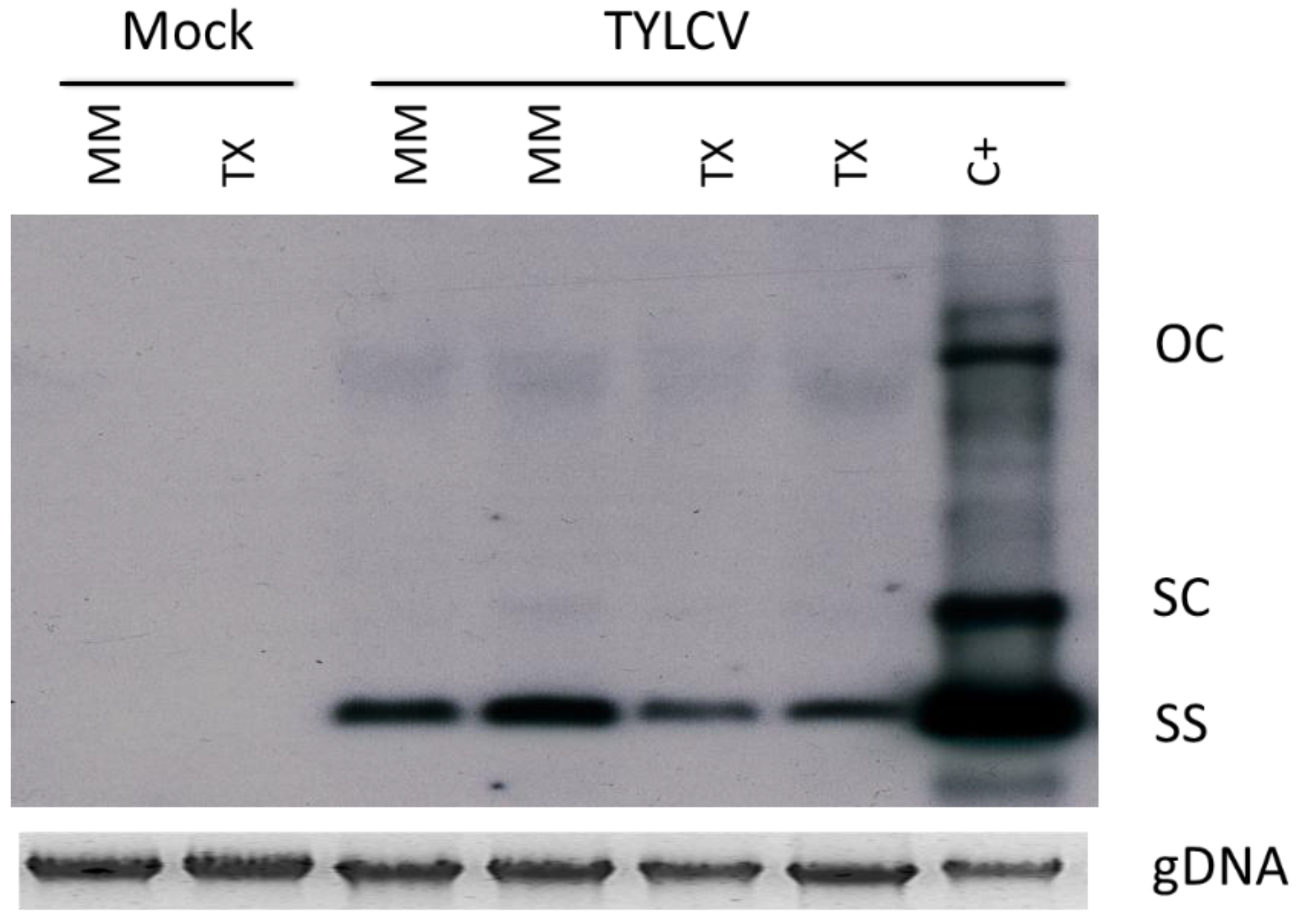
2.5. Virus Detection and Determination of TYLCV-IL Accumulation Levels in Infected Tomato Plants
2.6. Data Analysis
3. Results
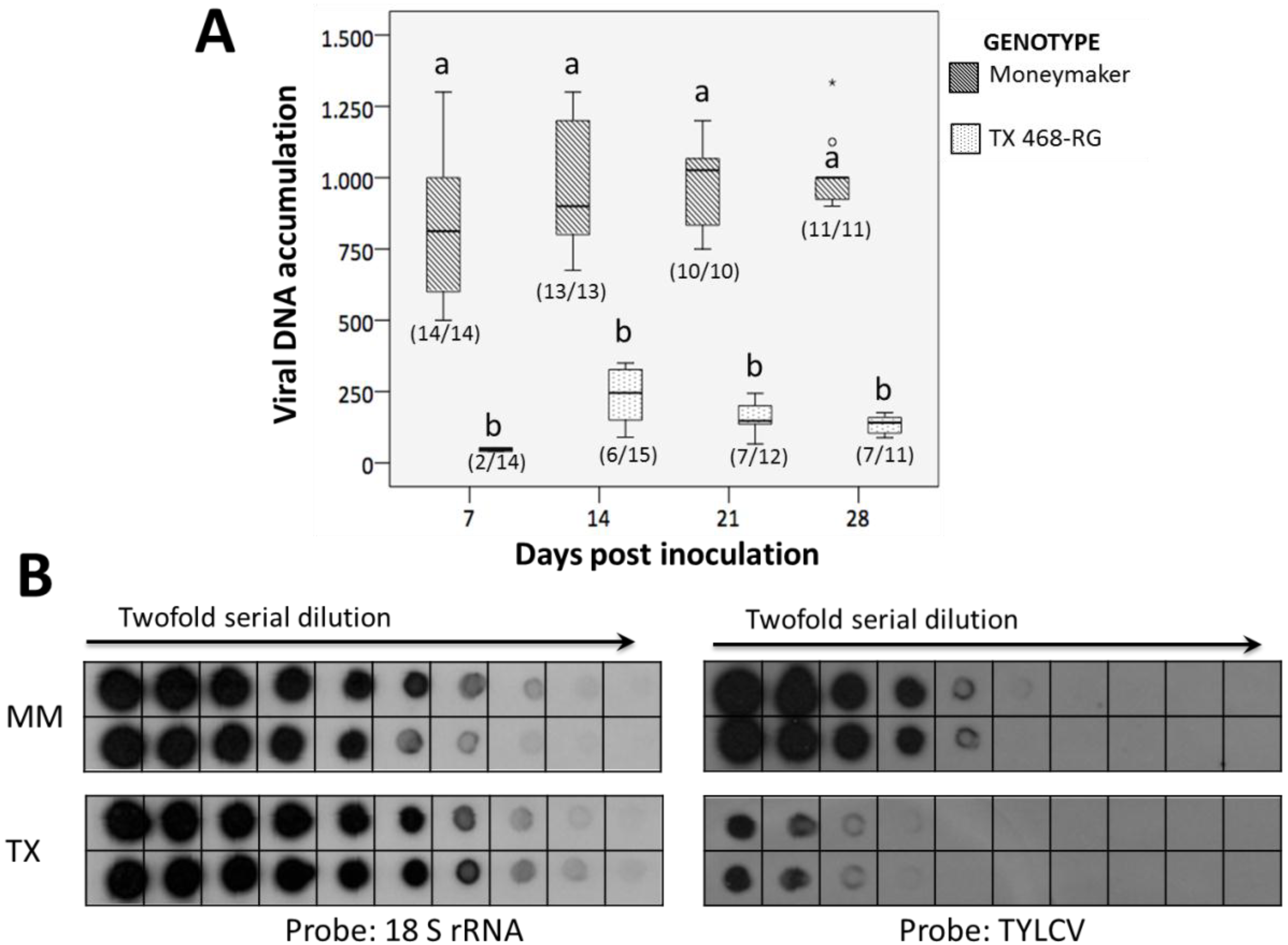
3.1. TYLCV-IL Accumulation is Highly Restricted in TX 468-RG
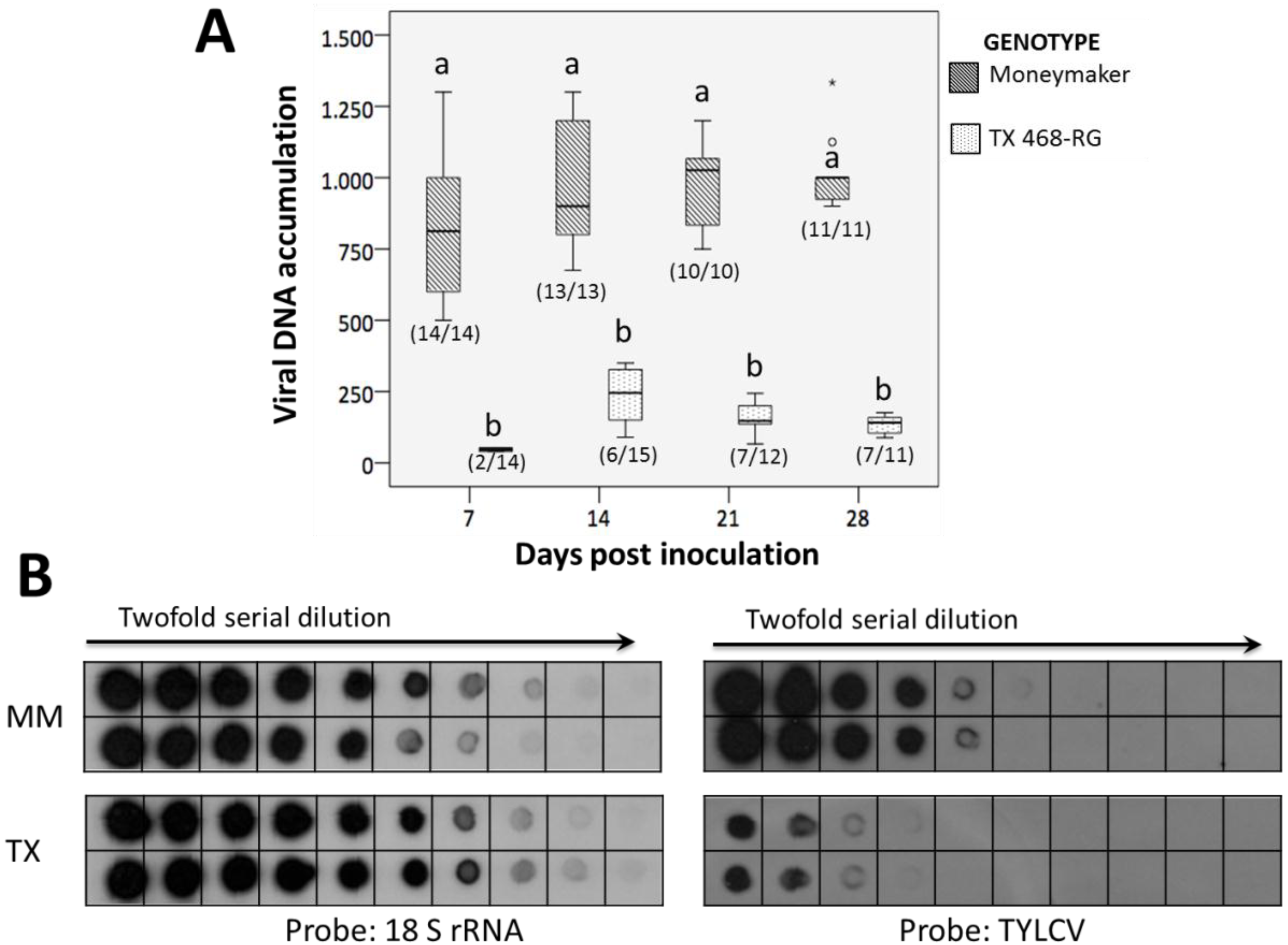
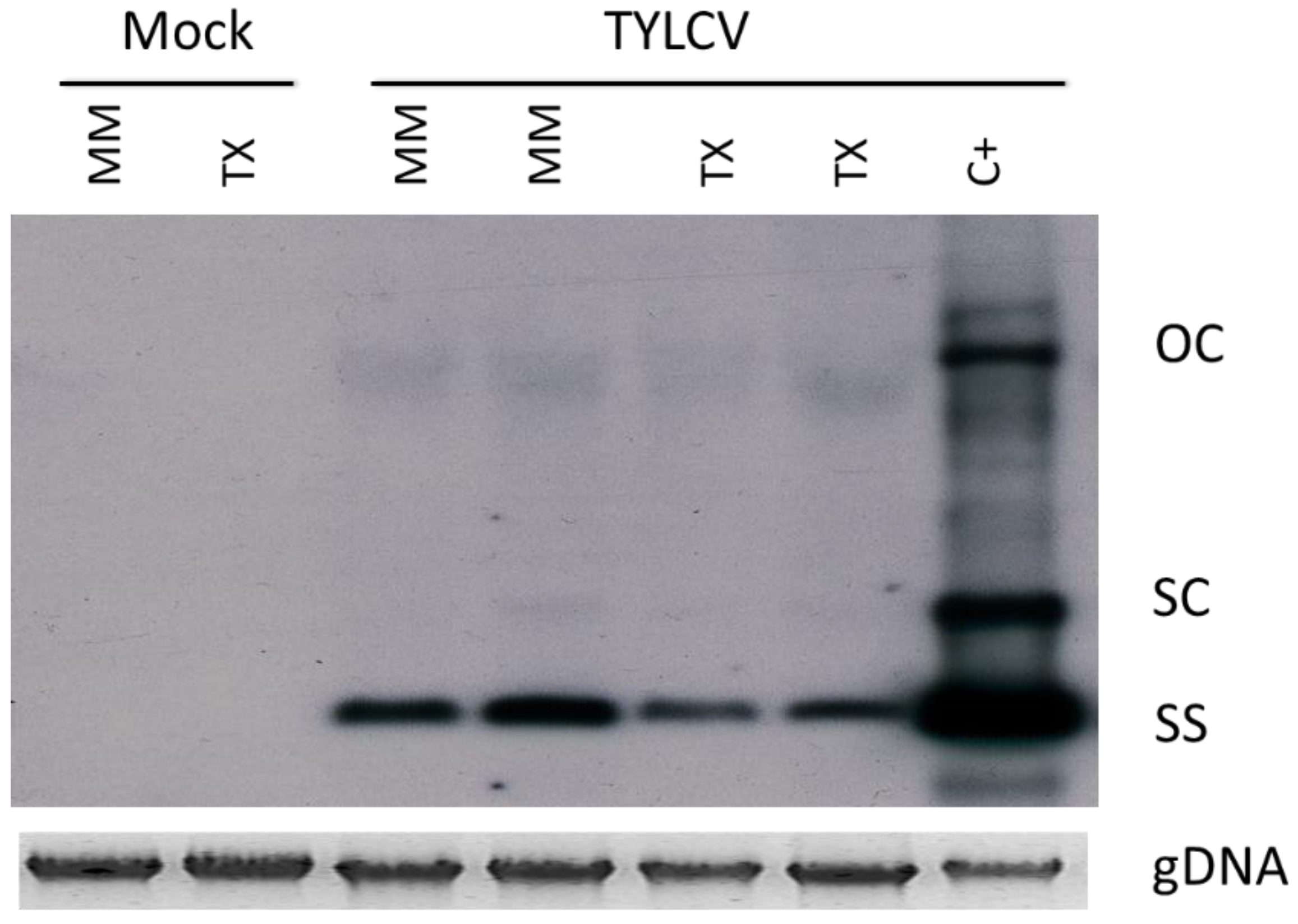

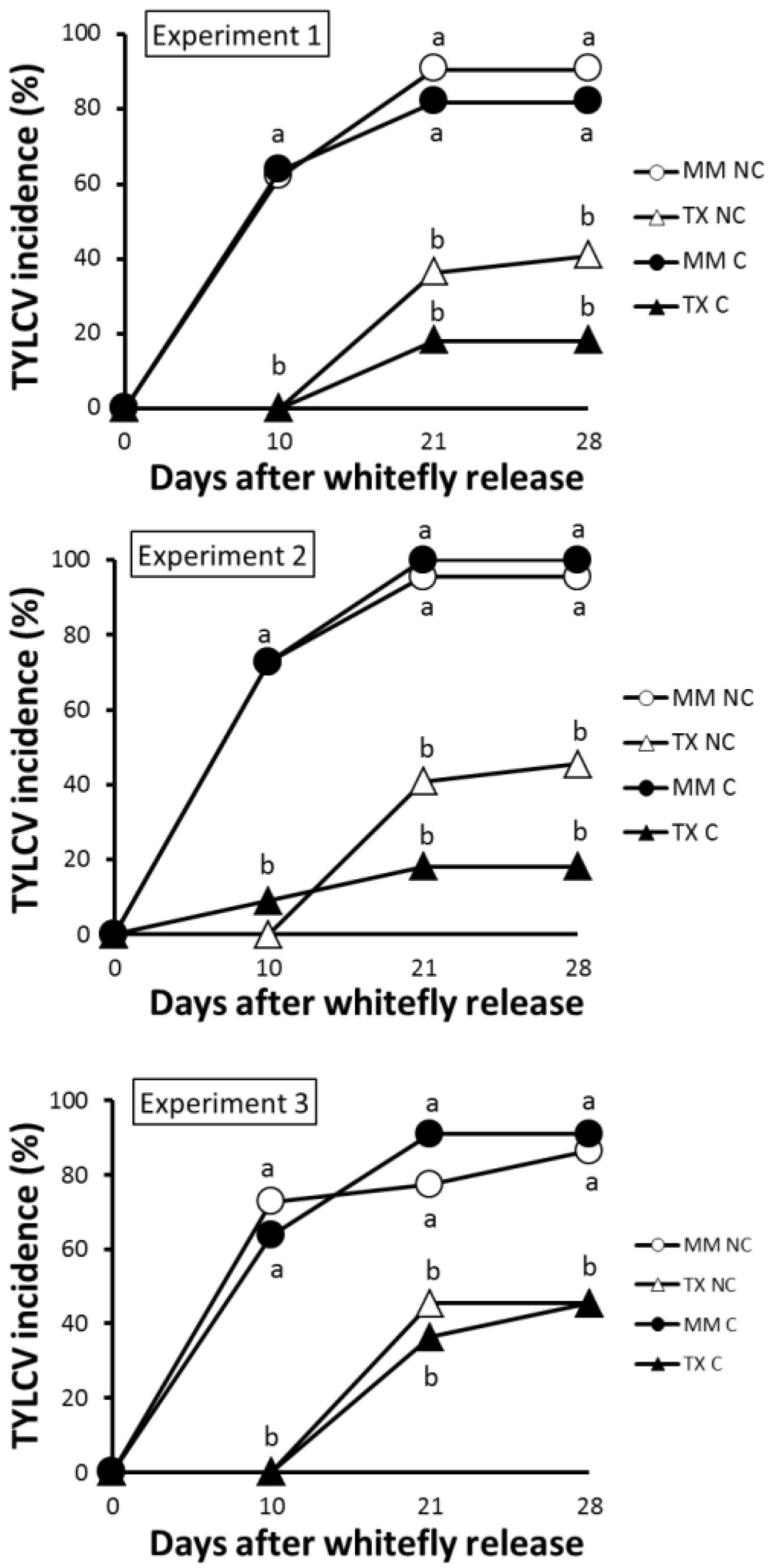
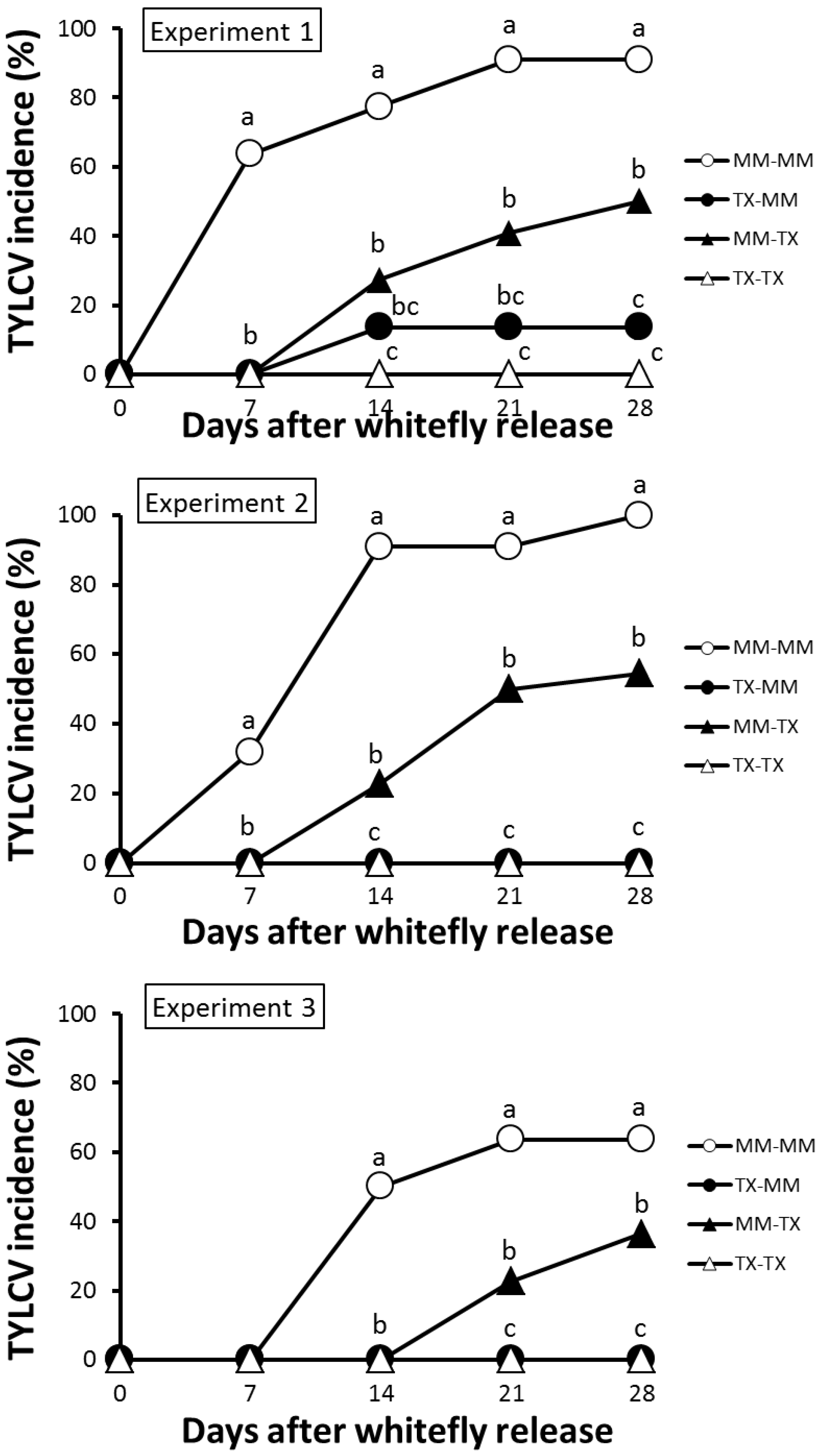
3.2. TX 468-RG Results in a Reduced Primary Spread of TYLCV-IL
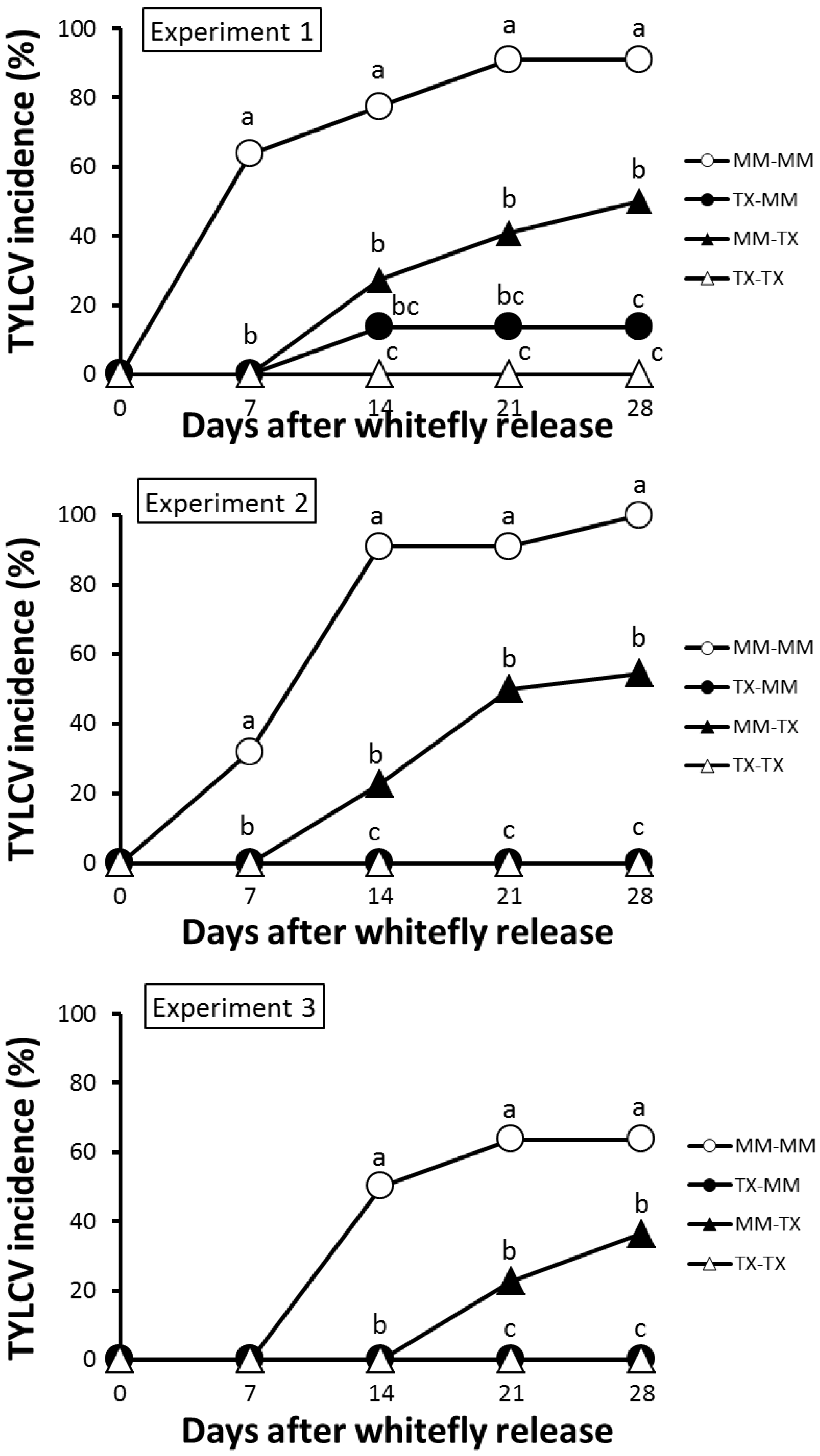
3.3. TX 468-RG Limits Secondary Spread of TYLCV-IL from Infected Source Plants to either TX 468-RG or MM
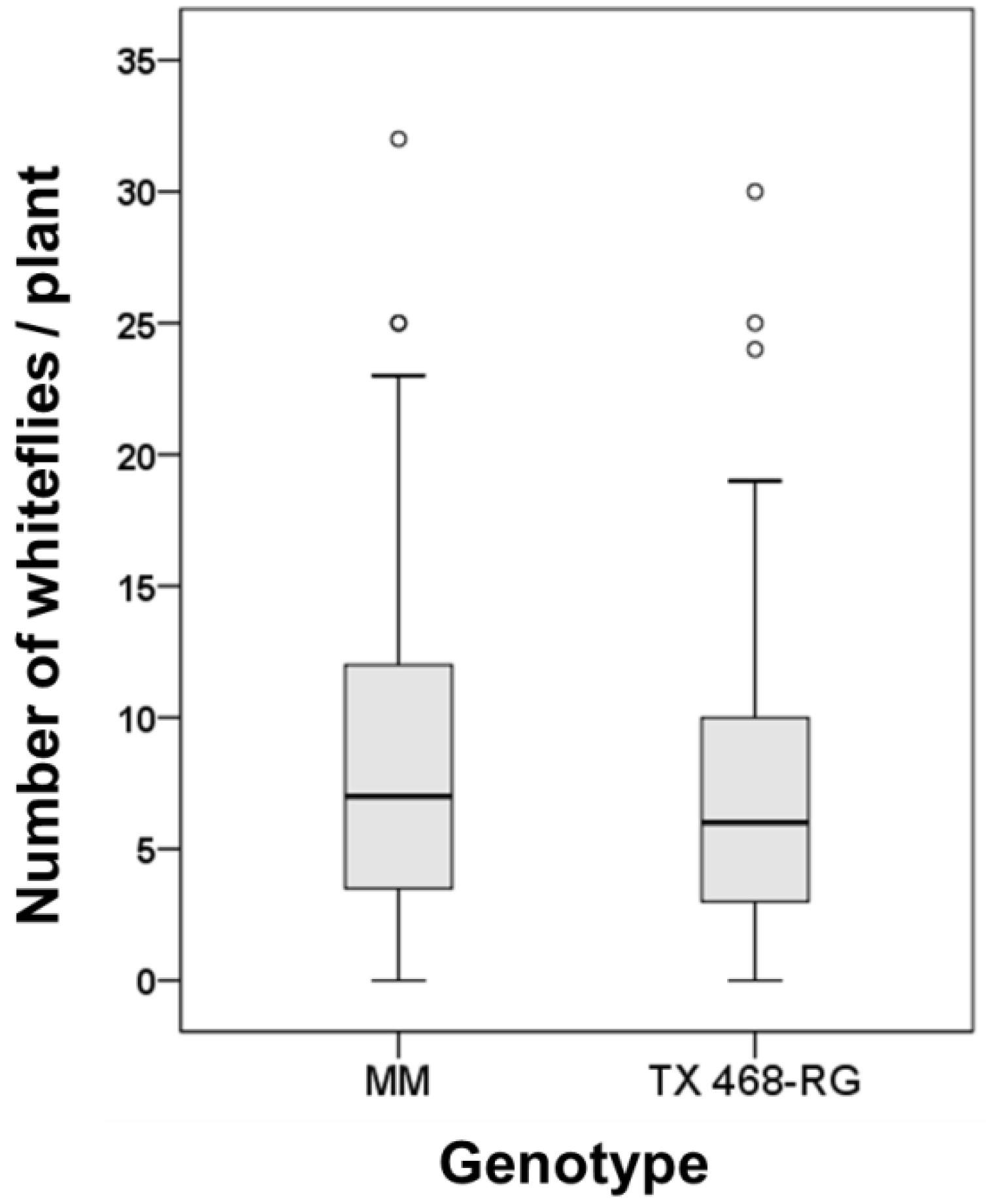
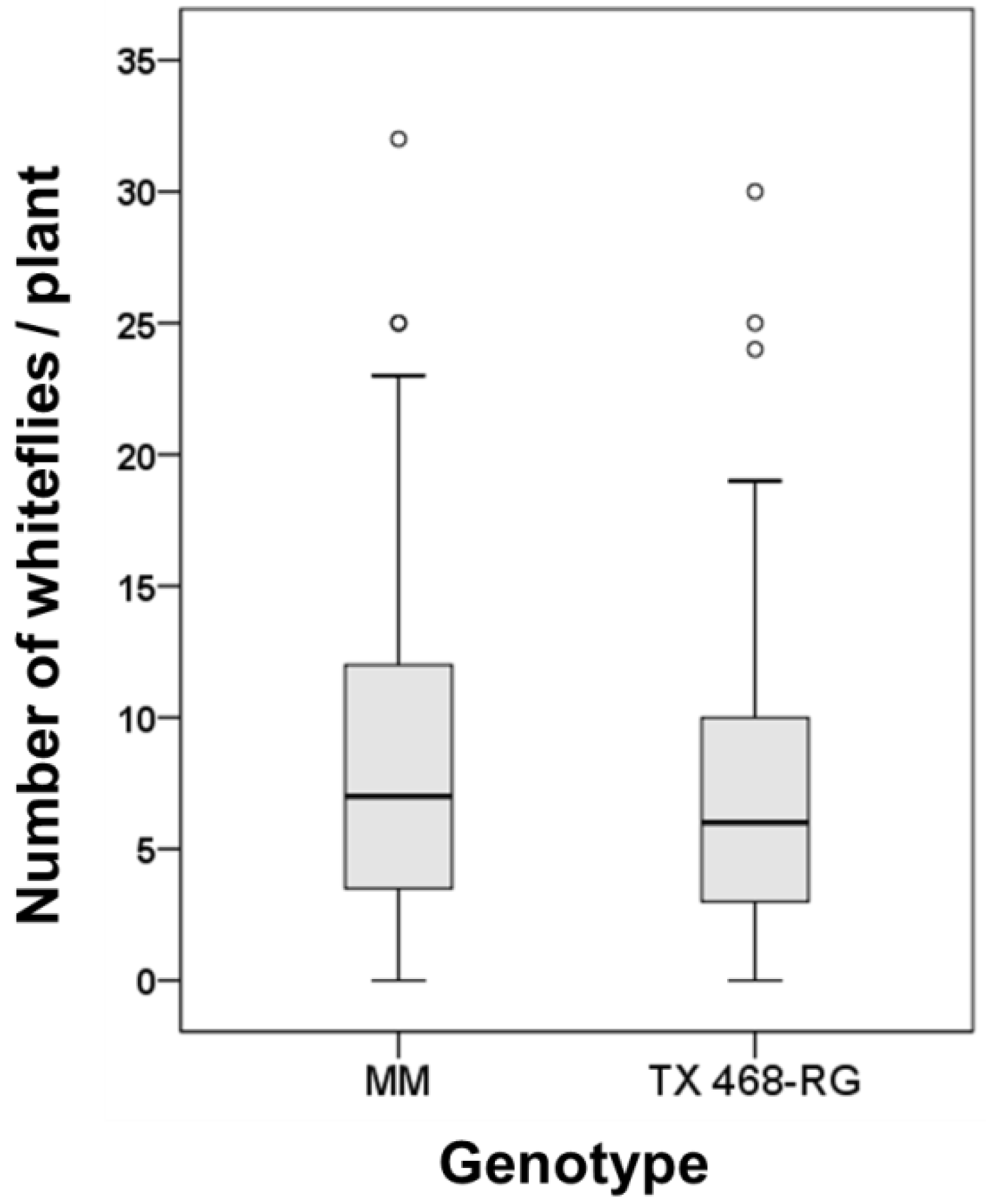
3.4. B. tabaci Exhibited no Preference for either MM or TX 468-RG
4. Discussion

Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hanssen, I.M.; Lapidot, M.; Thomma, B.P.H.J. Emerging viral diseases of tomato crops. Mol. Plant Microbe Interact. 2010, 23, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Navas-Castillo, J.; Fiallo-Olivé, E.; Sánchez-Campos, S. Emerging virus diseases transmitted by whiteflies. Annu. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar] [CrossRef] [PubMed]
- Czosnek, H.; Ghanim, M.; Ghanim, M. The circulative pathway of begomoviruses in the whitefly vector Bemisia tabaci—Insights from studies with Tomato yellow leaf curl virus. Ann. Appl. Biol. 2002, 14, 215–231. [Google Scholar] [CrossRef]
- Lefeuvre, P.; Martin, D.P.; Harkins, G.; Lemey, P.; Gray, A.J.; Meredith, S.; Lakay, F.; Monjane, A.; Lett, J.M.; Varsani, A.; et al. The spread of Tomato yellow leaf curl virus from the Middle East to the world. PLoS Pathog. 2010, 6, e1001164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordano, L.B.; Fonseca, M.E.N.; Silva, J.B.C.; Inoue-Nagata, A.K.; Boiteux, L.S. Efeito da infecção precoce por begomovirus com genoma bipartido em características de frutos de tomate industrial. Hort. Bras. 2005, 23, 815–818. [Google Scholar] [CrossRef]
- Cahill, M.; Gorman, K.; Day, S.; Denholm, I.; Elbert, A.; Nauen, R. Baseline determination and detection of resistance to imidacloprid in Bemisia tabaci (Homoptera: Aleyrodidae). Bull. Entomol. Res. 1996, 86, 343–349. [Google Scholar] [CrossRef]
- Cahill, M.; Jarvis, W.; Gorman, K.; Denholm, I. Resolution of baseline responses and documentation of resistance to buprofezin in Bemisia tabaci (Homoptera: Aleyrodidae). Bull. Entomol. Res. 1996, 86, 117–122. [Google Scholar] [CrossRef]
- Gómez, P.; Rodríguez-Hernandez, A.M.; Moury, B.; Aranda, M.A. Genetic resistance for the sustainable control of plant virus diseases: Breeding, mechanisms and durability. Eur. J. Plant Pathol. 2009, 125, 1–22. [Google Scholar] [CrossRef]
- Lapidot, M.; Friedmann, M. Breeding for resistance to whitefly-transmitted geminiviruses. Ann. Appl. Biol. 2002, 140, 109–127. [Google Scholar] [CrossRef]
- Zamir, D.; Eksteinmichelson, I.; Zakay, Y.; Navot, N.; Zeidan, M.; Sarfatti, M.; Eshed, Y.; Harel, E.; Pleban, T.; Vanoss, H.; et al. Mapping and introgression of a Tomato yellow leaf curl virus tolerance gene, Ty-1. Theor. Appl. Gen. 1994, 88, 141–146. [Google Scholar] [CrossRef]
- Barbieri, M.; Acciarri, N.; Sabatini, E.; Sardo, L.; Accotto, G.P.; Pecchioni, N. Introgression of resistance to two Mediterranean virus species causing tomato yellow leaf curl into a valuable traditional tomato variety. J. Plant Pathol. 2010, 92, 485–493. [Google Scholar]
- García-Andrés, S.; Tomás, D.M.; Sánchez-Campos, S.; Navas-Castillo, J.; Moriones, E. Frequent occurrence of recombinants in mixed infections of tomato yellow leaf curl disease-associated begomoviruses. Virology 2007, 365, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Michelson, I.; Zamir, D.; Czosnek, H. Accumulation and translocation of Tomato yellow leaf curl virus (TYLCV) in a Lycopersicon esculentum breeding line containing the L. chilense TYLCV tolerance gene Ty-1. Phytopathology 1994, 34, 928–933. [Google Scholar]
- Boiteux, L.S.; Oliveira, V.R.; Silva, C.H.; Makishima, N.; Inoue-Nagata, A.K.; Fonseca, M.E.N.; Giordano, L.B. Reaction of tomato hybrids carrying the Ty-1 locus to Brazilian bipartite Begomovirus species. Hort. Bras. 2007, 25, 20–23. [Google Scholar] [CrossRef]
- Davino, S.; Napoli, C.; Dellacroce, C.; Miozzi, L.; Noris, E.; Davino, M.; Accotto, G.P. Two new natural begomovirus recombinants associated with the tomato yellow leaf curl disease co-exist with parental viruses in tomato epidemics in Italy. Virus Res. 2009, 143, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Monci, F.; Sánchez-Campos, S.; Navas-Castillo, J.; Moriones, E. A natural recombinant between the geminiviruses Tomato yellow leaf curl Sardinia virus and Tomato yellow leaf curl virus exhibits a novel pathogenic phenotype and is becoming prevalent in Spanish populations. Virology 2002, 303, 317–326. [Google Scholar] [CrossRef] [PubMed]
- García-Cano, E.; Resende, R.O.; Boiteux, L.S.; Giordano, L.B.; Fernández-Muñoz, R.; Moriones, E. Phenotypic expression, stability, and inheritance of a recessive resistance to monopartite begomoviruses associated with tomato yellow leaf curl disease in tomato. Phytopathology 2008, 98, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Giordano, L.B.; Silva-Lobo, V.L.; Santana, F.M.; Fonseca, M.E.N.; Boiteux, L.S. Inheritance of resistance to the bipartite Tomato chlorotic mottle begomovirus derived from Lycopersicon esculentum cv. “Tyking”. Euphytica 2005, 143, 27–33. [Google Scholar] [CrossRef]
- Morilla, G.; Janssen, D.; García-Andrés, S.; Moriones, E.; Cuadrado, I.M.; Bejarano, E.R. Pepper (Capsicum annuum) is a dead-end host for Tomato yellow leaf curl virus. Phytopathology 2005, 95, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Monci, F.; García-Andrés, S.; Maldonado, J.A.; Moriones, E. Resistance to monopartite begomoviruses associated with the bean leaf crumple disease in Phaseolus vulgaris controlled by a single dominant gene. Phytopathology 2005, 95, 819–826. [Google Scholar] [CrossRef] [PubMed]
- Tomás, D.M.; Cañizares, C.; Abad, J.; Fernández-Muñoz, R.; Moriones, E. Resistance to tomato yellow leaf curl virus accumulation in the tomato wild relative Solanum habrochaites associated with the C4 viral protein. Mol. Plant Microbe Interact. 2011, 24, 849–861. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Lopez, M.J.; Garzo, E.; Bonani, J.P.; Fereres, A.; Fernández-Muñoz, R.; Moriones, E. Whitefly resistance traits derived from the wild tomato Solanum pimpinellifolium affect the preference and feeding behavior of Bemisia tabaci and reduce the spread of Tomato yellow leaf curl virus. Phytopathology 2011, 101, 1191–1201. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.L.; Madden, L.V. Introduction to Plant Disease Epidemiology; John Wiley and Sons: New York, NY, USA, 1990. [Google Scholar]
- Tukey, J.W. Box-and-Whisker Plots. In Exploratory Data Analysis; Addison-Wesley Publishing: Reading, MA, USA, 1977; pp. 39–43. [Google Scholar]
- Celix, A.; Lopez-Sesé, A.I.; Almarza, N.; Gómez-Guillamón, M.L.; Rodríguez-Cerezo, E. Characterization of Cucurbit yellow stunting disorder virus, a Bemisia tabaci-transmitted closterovirus. Phytopathology 1996, 86, 1370–1376. [Google Scholar]
- García-Andrés, S.; Tomás, D.M.; Navas-Castillo, J.; Moriones, E. Resistance-driven selection of begomoviruses associated with the tomato yellow leaf curl disease. Virus Res. 2009, 146, 66–72. [Google Scholar] [CrossRef] [PubMed]
- García-Cano, E.; Resende, R.O.; Fernández-Munoz, R.; Moriones, E. Synergistic interaction between Tomato chlorosis virus and Tomato spotted wilt virus results in breakdown of resistance in tomato. Phytopathology 2006, 96, 1263–1269. [Google Scholar] [CrossRef] [PubMed]
- Fraser, R.S.S. The genetics of resistance to plant-viruses. Annu. Rev. Phytopathol. 1990, 28, 179–200. [Google Scholar] [CrossRef]
- Hull, R. Matthew’s Plant Virology, 4th ed.; Academic Press: San Diego, CA, USA, 2002. [Google Scholar]
- Ueki, S.; Citovsky, V. Spread throughout the plant: Systemic transport of viruses. In Viral Transport in Plants; Waigmann, E., Heinlein, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 85–118. [Google Scholar]
- Chellapan, P.; Vanitharani, R.; Fauquet, C.M. Short interfering RNA accumulation correlates with host recovery in DNA virus-infected hosts, and gene silencing targets specific viral sequences. J. Virol. 2004, 78, 7465–7477. [Google Scholar] [CrossRef] [PubMed]
- Bian, X.Y.; Thomas, M.R.; Rasheed, M.S.; Saeed, M.; Hanson, P.; de Barro, P.J.; Rezaian, M.A. A recessive allele (tgr-1) conditioning tomato resistance to geminivirus infection is associated with impaired viral movement. Phytopathology 2007, 97, 930–937. [Google Scholar] [CrossRef] [PubMed]
- Fargette, D.; Leslie, M.; Harrison, B.D. Serological studies on the accumulation and localisation of three tomato leaf curl geminiviruses in resistant and susceptible Lycopersicon species and tomato culivars. Ann. Appl. Biol. 1996, 128, 317–328. [Google Scholar] [CrossRef]
- Friedmann, M.; Lapidot, M.; Cohen, S.; Pilowsky, M. A novel source of resistance to tomato yellow leaf curl virus exhibiting a symptomless reaction to viral infection. J. Am. Soc. Hort. Sci. 1998, 123, 1004–1007. [Google Scholar]
- Lapidot, M.; Friedmann, M.; Pilowsky, M.; Ben Joseph, R.; Cohen, S. Effect of host plant resistance to Tomato yellow leaf curl virus (TYLCV) on virus acquisition and transmission by its whitefly vector. Phytopathology 2001, 91, 1209–1213. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.; Riley, D.; Diffie, S.; Sparks, A.; Adkins, S. Whitefly population dynamics and evaluation of whitefly-transmitted Tomato yellow leaf curl virus (TYLCV)-resistant tomato genotypes as whitefly and TYLCV reservoirs. J. Econ. Entomol. 2012, 105, 1447–1456. [Google Scholar] [CrossRef] [PubMed]
- Escriu, F.; Perry, K.L.; García-Arenal, F. Transmissibility of Cucumber mosaic virus by Aphis gossypii correlates with viral accumulation and is affected by the presence of its satellite RNA. Phytopathology 2000, 90, 1068–1072. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Antignus, Y. Tomato yellow leaf curl virus (TYLCV), a whitefly-borne geminivirus of tomato. Adv. Dis. Vector Res. 1994, 10, 259–288. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira-Carvalho, R.C.; Díaz-Pendón, J.A.; Fonseca, M.E.N.; Boiteux, L.S.; Fernández-Muñoz, R.; Moriones, E.; Resende, R.O. Recessive Resistance Derived from Tomato cv. Tyking-Limits Drastically the Spread of Tomato Yellow Leaf Curl Virus. Viruses 2015, 7, 2518-2533. https://doi.org/10.3390/v7052518
Pereira-Carvalho RC, Díaz-Pendón JA, Fonseca MEN, Boiteux LS, Fernández-Muñoz R, Moriones E, Resende RO. Recessive Resistance Derived from Tomato cv. Tyking-Limits Drastically the Spread of Tomato Yellow Leaf Curl Virus. Viruses. 2015; 7(5):2518-2533. https://doi.org/10.3390/v7052518
Chicago/Turabian StylePereira-Carvalho, Rita C., Juan A. Díaz-Pendón, Maria Esther N. Fonseca, Leonardo S. Boiteux, Rafael Fernández-Muñoz, Enrique Moriones, and Renato O. Resende. 2015. "Recessive Resistance Derived from Tomato cv. Tyking-Limits Drastically the Spread of Tomato Yellow Leaf Curl Virus" Viruses 7, no. 5: 2518-2533. https://doi.org/10.3390/v7052518