1. Introduction
Chronic bee paralysis virus (CBPV) causes chronic paralysis, an infectious disease of adult honey bees. The symptoms of this disease include severe trembling of the wings and bodies, and diseased bees often crawl on the ground. Some individuals become hairless and, thus, darker in appearance. Diseased bees die within a few days [
1,
2]. CBPV can persist as an unapparent infection but may multiply to high levels in honey bee colonies [
3], causing significant losses [
4]. Although CBPV shares several characteristics with the
Nodaviridae and
Tombusviridae virus families, CBPV is considered a new family of positive single-stranded RNA viruses. The sizes of the two major CBPV RNAs were determined to be 3674 bases for RNA 1 and 2305 bases for RNA 2 [
5].
We previously conducted the first nationwide epidemiological survey in Japan of honey bee parasites and pathogens, to measure their impact on colonies of the non-native European honey bee
Apis mellifera and of the native Japanese honey bee
Apis cerana japonica [
6]. We found that microsporidia,
Nosema ceranae, but not
Nosema apis, commonly infects
A. mellifera colonies (64% of colonies tested). Honey bee viruses, such as deformed wing virus (DWV), black queen cell virus (BQCV), and Israel acute paralysis virus (IAPV), are also prevalent (>66% of colonies tested) in
A. mellifera colonies. The prevalence of
Nosema and viruses in
A. c. japonica colonies is lower than that in
A. mellifera colonies. Our survey also revealed a tracheal mite (
Acarapis woodi) infestation in both honey bee species for the first time in Japan. These results demonstrate the infestation of native honey bees by parasite and pathogens of non-native honey bees that are traded globally [
6]. Although CBPV shows a worldwide distribution [
7], our previous study of seven viruses (ABPV, BQCV, CBPV, DWV, IAPV, KBV, and SBV) infecting healthy honey bee (
A. mellifera and
A. c. japonica) colonies in Japan did not detect CBPV in any of the samples analyzed. However, we later found CBPV by RNAseq analysis (high-throughput sequencing of mRNA) in dying workers from two colonies (Japan-2 and -5) used for pollination of strawberries in greenhouses. This observation led us to test for CBPV infection in both healthy and collapsing
A. mellifera and
A. c. japonica colonies.
2. Results
Healthy 60
A. mellifera and 55
A. c. japonica colonies without significant loss of workers were analyzed for CBPV infection by collecting and pooling 50 workers from a single colony. In addition, approximately 50 dying workers (crawling on the ground in front of the hive entrance) from a single colony were sampled and pooled for 29 collapsing
A. mellifera colonies with significant loss of workers. This investigation has identified 3 more CBPV-infected
A. mellifera colonies (Japan-1, -3, and -4) among the collapsing colonies. CBPV-positive
A. c. japonica colonies have not been found.
Table 1 summarizes the presence of pathogens and parasites in five CBPV-infected
A. mellifera colonies with severe loss of workers. CBPV was not found in healthy colonies. Infection by multiple viruses was detected in four colonies (Japan-1, -2, -3, and -4), and
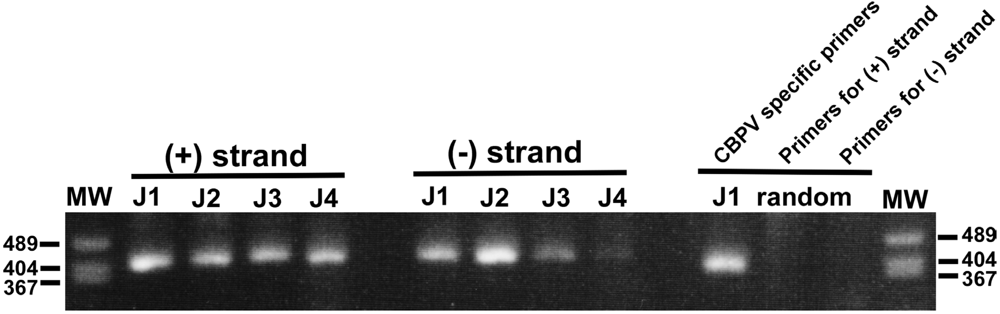
N. ceranae is present in all colonies. Furthermore, the Japan-3 colony is also infested with tracheal mites. To examine a possible role of CBPV infection in the severe loss of workers, the level of viral replication was analyzed by detecting the (−) strand RNA of the viral genome. The (−) strand viral RNA is synthesized by RNA-dependent RNA polymerase (RdRP) using the (+) strand viral RNA genome as a template. This is then used as a template to synthesize the (+) strand viral RNA genome by RdRP. A similar level of both (+) and (−) strand RNAs were found in the dying workers from the Japan-1, -2, and -3 colonies, suggesting that CBPV proliferates in the workers themselves (
Figure 1). To test for the specificity of detecting the (+) and (−) strands of CBPV RNA2 by strand-specific RT-PCR, we performed an additional control experiment, in which we analyzed CBPV cDNAs obtained with random hexamer primers. Although the CBPV-specific primers generated a positive band, the two sets of primers used for the strand-specific RT-PCR failed to give positive signals (
Figure 1). The relative level of (−) strand RNA was low in dying workers from the Japan-4 colony, and neither (+) nor (−) strand RNAs were detected in dying workers from the Japan-5 colony with this method (data not shown). Thus, CBPV infection and proliferation may contribute to the loss of workers from the Japan-1, -2, and -3 but not from the Japan-4 and -5 colonies.
Table 1.
Summary of pathogens and parasite present in CBPV-infected A. mellifera colonies.
Table 1.
Summary of pathogens and parasite present in CBPV-infected A. mellifera colonies.
| Sample | Virus | Microsporidia | Protist | Tracheal mite | Severe loss of workers | Use for pollination |
|---|
| Japan-1 | CBPV, BQCV, DWV | N. ceranae | − | − | + | − |
| Japan-2 | CBPV, BQCV, DWV | N. ceranae | − | − | + | + |
| Japan-3 | CBPV, BQCV, DWV, IAPV, SBV | N. ceranae | − | + | + | − |
| Japan-4 | CBPV, BQCV, DWV, IAPV | N. ceranae | − | − | + | − |
| Japan-5 | CBPV | N. ceranae | − | − | + | + |
The presence of microsporidia (N. apis and N. ceranae), protists (Crithidia mellificae and Apicystis bombi), and tracheal mite in five CBPV-infected A. mellifera colonies (Japan-1 to -5) was examined by PCR. A severe loss of workers was found with all colonies, and Japan-2 and -5 colonies were used for pollination in strawberry greenhouses.
Figure 1.
Molecular detection of the (+) and (−) strands of CBPV RNA2 in dying workers.(+) and (−) strands of CBPV RNA2 in dying workers from the A. mellifera Japan-1, -2, -3, and -4 colonies (J1 to J4) have been detected separately by strand-specific RT-PCR. Both (+) and (−) strand RNAs are present in the dying workers; however, the relative level of (−) strand RNA in those from the Japan-4 (J4) colony is low. Control PCR reactions were performed using CBPV cDNAs (Japan-1, J1) generated with random hexamer primers (“random”) and the CBPV-specific primers or using the two sets of primers used for detecting the (+) and (−) strands of CBPV RNA2. The sizes of three bands of the molecular weight marker (MW) are shown on both sides of the gel.
Figure 1.
Molecular detection of the (+) and (−) strands of CBPV RNA2 in dying workers.(+) and (−) strands of CBPV RNA2 in dying workers from the A. mellifera Japan-1, -2, -3, and -4 colonies (J1 to J4) have been detected separately by strand-specific RT-PCR. Both (+) and (−) strand RNAs are present in the dying workers; however, the relative level of (−) strand RNA in those from the Japan-4 (J4) colony is low. Control PCR reactions were performed using CBPV cDNAs (Japan-1, J1) generated with random hexamer primers (“random”) and the CBPV-specific primers or using the two sets of primers used for detecting the (+) and (−) strands of CBPV RNA2. The sizes of three bands of the molecular weight marker (MW) are shown on both sides of the gel.
Figure 2.
Phylogeny of Japanese, European, Uruguayan, and mainland US CBPV isolates. The condensed phylogenetic tree (bootstrap values <50%) based on alignment of the putative RdRP gene sequences of Japanese, European, Uruguayan, and mainland US CBPV isolates was constructed using the maximum likelihood method. The number of each node represents the bootstrap value resulting from 1000replicates. Each isolate is indicated by its accession number.
Figure 2.
Phylogeny of Japanese, European, Uruguayan, and mainland US CBPV isolates. The condensed phylogenetic tree (bootstrap values <50%) based on alignment of the putative RdRP gene sequences of Japanese, European, Uruguayan, and mainland US CBPV isolates was constructed using the maximum likelihood method. The number of each node represents the bootstrap value resulting from 1000replicates. Each isolate is indicated by its accession number.
The phylogeny of Japanese, European, Uruguayan, and mainland US CBPV isolates was constructed using the maximum likelihood method and the putative RdRP coding sequences (
Figure 2). This gene was selected for the analysis since many sequences of the isolates from different countries are available in a GenBank. As previously reported [
8,
9], the European isolates are grouped into two major clades, except two isolates: one French (EU122231) and one Swiss (FJ345324). Uruguayan isolates also cluster with one exception (FJ345326). Ten mainland US isolates form the independent clade, and four Japanese isolates (Japan-1, -3, -4, and -5) also independently cluster. However, one Japanese isolate (Japan-2) is basal to the mainland US and Japanese isolates (
Figure 2). Although there are several exceptions, European, mainland US, Uruguayan, and Japanese isolates show the trends of geographic separation.
3. Discussion
This reports a molecular identification of CBPV in Japan, and is consistent with its worldwide distribution [
7], which may result from global trade of
A. mellifera queens among many countries. Although our current investigation has revealed a lower prevalence of CBPV in Japan, relative to other viruses, including DWV, BQCV, IAPV, and SBV [
6], this prevalence could be underestimated because one primer set we used for primary screening may fail to detect more divergent strains of CBPV infecting both
A. mellifera and
A. c. japonica. We have not found CBPV-positive
A. c. japonica colonies in the current study, suggesting that CBPV may have a species barrier for infection, as demonstrated with SBV [
6,
10]. Alternatively, the low prevalence in
A. mellifera colonies may, itself, explain the absence of CBPV in
A. c. japonica colonies. A survey of honey bee viruses in China indicated that the prevalence of CBPV was 11.8% in 34 apiaries with
A. mellifera colonies [
11], and appears to be higher than that in Japan (5.6%). CBPV infection is not thought to induce massive colony losses, but it could contribute to a severe loss of workers in an infected colony, as we observed with the Japan-1, -2, and -3 colonies. However, these colonies were also infected with other viruses and
N. ceranae (
Table 1), suggesting that CBPV may not be the single cause for the loss of workers. The relative level of (−) strand RNA was high in dying workers from the Japan-2 colony used for pollination in a strawberry green house. It demonstrates that CBPV actively proliferates in these workers (
Figure 1), and this is likely to result from the immune suppression of honey bees used for long-term pollination in greenhouses [
12].
The geographic separation of Japanese CBPV isolates from those of other countries is quite similar to the separation we previously observed with DWV [
6]. The lack of major honey bee trade between Europe/mainland US and Japan in the last 26 years (1985–2010) may have resulted in the geographic separation of Japanese isolates. Twenty-seven and 100 queen bees were imported from Italy and Switzerland to Japan in 1987 and 1997, respectively. Two hundred and five queen bees were imported from Yugoslavia during 1989–1991, and 2138 queen bees were imported from Slovenia during 1998–2010. Similarly, 77 queen bees were imported from mainland US during 1985–1992. This phylogenetic relationship did not change even when the tree was rooted by including an outgroup virus, for example, BQCV (data not shown). We also constructed a phylogenetic tree based on a 527-nucleotide sequence encoding the putative structural proteins of CBPV isolates from Japan and other countries. The putative structural protein gene was previously identified in RNA2 [
5]. The Japanese isolates also cluster, but form an independent clade along with two Uruguayan isolates and one Swiss isolate (data not shown). However, as described by [
8], the amino acid sequences of these structural proteins show a very low degree of divergence, suggesting that they may not be suitable for inferring the phylogeny. Since Japan has imported most of its honey bee queens from Hawaii (12486 queen bees), New Zealand (51761 queen bees), and Australia (87698 queen bees) in the last 26 years, it would be interesting to determine the phylogeny of CBPV isolates from Japan and from these areas. Such a phylogeny may reveal the role of honey bee trade for viral transmission between countries.



{kind=link}
{kind=link}
{kind=link}
{kind=link}