
Wide Real-Life Data Support Reduced Sensitivity of Antigen Tests for Omicron SARS-CoV-2 Infections
 , , , , , , , , , ,
, , , , , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Ethics
2.3. SARS-CoV-2 Antigen Diagnostic Tests
- Group 1 ADT: Lateral Flow Immunochromatography rapid assay
- Group 2 ADT: Microfluidic-based rapid assay
- Group 3 ADT: Chemiluminescence-based assay
2.4. SARS-CoV-2 RT-PCR Analysis
2.5. SARS-CoV-2 Genome Sequencing
2.6. Bioinformatic Analysis of Genome Sequences
2.7. Nucleocapsid (N) Protein Mutation Analysis
2.8. Statistical Analysis
3. Results
3.1. Evaluation of ADT Performance in Delta versus Omicron VOCs Period
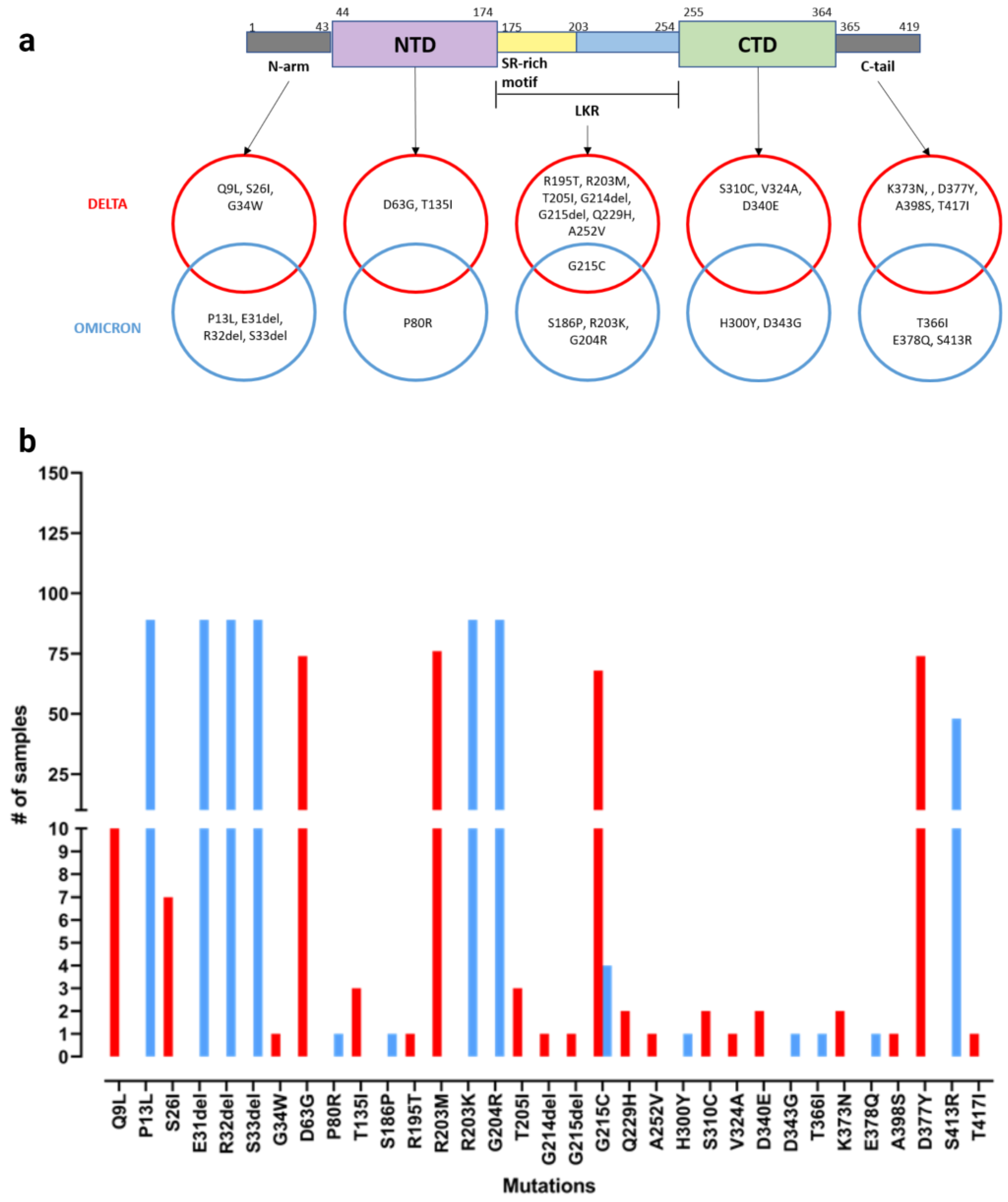
3.2. Evaluation of Nucleocapsid Protein Mutations in Delta and Omicron Variants
3.3. Mouth versus Nose Viral Load in Omicron-Infected Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Oragnization. WHO Coronavirus (COVID-19) Dashboard. 2023. Available online: https://data.who.int/dashboards/covid19/ (accessed on 16 April 2024).
- Tromberg, B.J.; Schwetz, T.A.; Pérez-Stable, E.J.; Hodes, R.J.; Woychik, R.P.; Bright, R.A.; Fleurence, R.L.; Collins, F.S. Rapid Scaling Up of Covid-19 Diagnostic Testing in the United States—The NIH RADx Initiative. N. Engl. J. Med. 2020, 383, 1071–1077. [Google Scholar] [CrossRef] [PubMed]
- Fourati, S.; Langendorf, C.; Audureau, E.; Challine, D.; Michel, J.; Soulier, A.; Ahnou, N.; Désveaux, I.; Picard, O.; Ortonne, V.; et al. Performance of Six Rapid Diagnostic Tests for SARS-CoV-2 Antigen Detection and Implications for Practical Use. J. Clin. Virol. 2021, 142, 104930. [Google Scholar] [CrossRef]
- Bai, Z.; Cao, Y.; Liu, W.; Li, J. The SARS-CoV-2 Nucleocapsid Protein and Its Role in Viral Structure, Biological Functions, and a Potential Target for Drug or Vaccine Mitigation. Viruses 2021, 13, 1115. [Google Scholar] [CrossRef] [PubMed]
- Diao, B.; Wen, K.; Zhang, J.; Chen, J.; Han, C.; Chen, Y.; Wang, S.; Deng, G.; Zhou, H.; Wu, Y. Accuracy of a Nucleocapsid Protein Antigen Rapid Test in the Diagnosis of SARS-CoV-2 Infection. Clin. Microbiol. Infect. 2021, 27, 289.e1–289.e4. [Google Scholar] [CrossRef]
- Mak, G.C.; Cheng, P.K.; Lau, S.S.; Wong, K.K.; Lau, C.; Lam, E.T.; Chan, R.C.; Tsang, D.N. Evaluation of Rapid Antigen Test for Detection of SARS-CoV-2 Virus. J. Clin. Virol. 2020, 129, 104500. [Google Scholar] [CrossRef]
- Porte, L.; Legarraga, P.; Vollrath, V.; Aguilera, X.; Munita, J.M.; Araos, R.; Pizarro, G.; Vial, P.; Iruretagoyena, M.; Dittrich, S.; et al. Evaluation of a Novel Antigen-Based Rapid Detection Test for the Diagnosis of SARS-CoV-2 in Respiratory Samples. Int. J. Infect. Dis. 2020, 99, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Fang, Z.; Ma, F.; Li, J.; Huang, Z.; Zhang, Y.; Li, J.; Chen, K. The Role of N Protein in Diagnosis and Vaccination in the Context of Variants: Present Status and Prospects. Front. Microbiol. 2023, 14, 1217567. [Google Scholar] [CrossRef]
- Viana, R.; Moyo, S.; Amoako, D.G.; Tegally, H.; Scheepers, C.; Althaus, C.L.; Anyaneji, U.J.; Bester, P.A.; Boni, M.F.; Chand, M.; et al. Rapid Epidemic Expansion of the SARS-CoV-2 Omicron Variant in Southern Africa. Nature 2022, 603, 679–686. [Google Scholar] [CrossRef]
- Assessment of the Further Spread and Potential Impact of the SARS-CoV-2 Omicron Variant of Concern in the EU/EEA, 19th Update. Available online: https://www.ecdc.europa.eu/en/publications-data/covid-19-omicron-risk-assessment-further-emergence-and-potential-impact (accessed on 31 March 2024).
- National Institute for Communicable Diseases. The Daily Covid-19 Effective Reproductive Number (R) in South Africa. 2021. Available online: https://www.nicd.ac.za/wp-content/uploads/2021/12/COVID-19-Effective-Reproductive-Number-in-South-Africa-week-51.pdf (accessed on 4 March 2024).
- Pulliam, J.R.C.; Van Schalkwyk, C.; Govender, N.; Von Gottberg, A.; Cohen, C.; Groome, M.J.; Dushoff, J.; Mlisana, K.; Moultrie, H. Increased Risk of SARS-CoV-2 Reinfection Associated with Emergence of Omicron in South Africa. Science 2022, 376, eabn4947. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cheng, G. Sequence Analysis of the Emerging SARS-CoV-2 Variant Omicron in South Africa. J. Med. Virol. 2022, 94, 1728–1733. [Google Scholar] [CrossRef]
- Yang, Q.; Syed, A.A.S.; Fahira, A.; Shi, Y. Structural Analysis of the SARS-CoV-2 Omicron Variant Proteins. Research 2021, 2021, 9769586. [Google Scholar] [CrossRef] [PubMed]
- Boehm, E.; Kronig, I.; Neher, R.A.; Eckerle, I.; Vetter, P.; Kaiser, L. Novel SARS-CoV-2 Variants: The Pandemics within the Pandemic. Clin. Microbiol. Infect. 2021, 27, 1109–1117. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Yin, W.; Jiang, Y.; Xu, H.E. Structure Genomics of SARS-CoV-2 and Its Omicron Variant: Drug Design Templates for COVID-19. Acta Pharmacol. Sin. 2022, 43, 3021–3033. [Google Scholar] [CrossRef] [PubMed]
- Ferré, V.M.; Peiffer-Smadja, N.; Visseaux, B.; Descamps, D.; Ghosn, J.; Charpentier, C. Omicron SARS-CoV-2 Variant: What We Know and What We Don’t. Anaesth. Crit. Care Pain Med. 2022, 41, 100998. [Google Scholar] [CrossRef] [PubMed]
- Deerain, J.; Druce, J.; Tran, T.; Batty, M.; Yoga, Y.; Fennell, M.; Dwyer, D.E.; Kok, J.; Williamson, D.A. Assessment of the Analytical Sensitivity of 10 Lateral Flow Devices against the SARS-CoV-2 Omicron Variant. J. Clin. Microbiol. 2022, 60, e02479-e21. [Google Scholar] [CrossRef]
- Stanley, S.; Hamel, D.J.; Wolf, I.D.; Riedel, S.; Dutta, S.; Cheng, A.; Kirby, J.E.; Kanki, P.J. Limit of Detection for Rapid Antigen Testing of the SARS-CoV-2 Omicron and Delta Variants of Concern Using Live-Virus Culture. J. Clin. Microbiol. 2022, 60, e00140-22. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.; Westbrook, A.; Bassit, L.; Parsons, R.; Fitts, E.; Greenleaf, M.; McLendon, K.; Sullivan, J.A.; O’Sick, W.; Baugh, T.; et al. Sensitivity of rapid antigen tests against SARS-CoV-2 Omicron and Delta variants. J. Clin. Microbiol. 2023, 61, e0013823. [Google Scholar] [CrossRef]
- Widyasari, K.; Kim, S. Efficacy of novel SARS-CoV-2 rapid antigen tests in the era of omicron outbreak. PLoS ONE 2023, 18, e0289990. [Google Scholar] [CrossRef]
- Osterman, A.; Badell, I.; Basara, E.; Stern, M.; Kriesel, F.; Eletreby, M.; Öztan, G.N.; Huber, M.; Autenrieth, H.; Knabe, R.; et al. Impaired Detection of Omicron by SARS-CoV-2 Rapid Antigen Tests. Med. Microbiol. Immunol. 2022, 211, 105–117. [Google Scholar] [CrossRef]
- Schuit, E.; Veldhuijzen, I.K.; Venekamp, R.P.; Van Den Bijllaardt, W.; Pas, S.D.; Lodder, E.B.; Molenkamp, R.; GeurtsvanKessel, C.H.; Velzing, J.; Huisman, R.C.; et al. Diagnostic Accuracy of Rapid Antigen Tests in Asymptomatic and Presymptomatic Close Contacts of Individuals with Confirmed SARS-CoV-2 Infection: Cross Sectional Study. BMJ 2021, 374, n1676. [Google Scholar] [CrossRef]
- Marais, G.; Hsiao, N.; Iranzadeh, A.; Doolabh, D.; Joseph, R.; Enoch, A.; Chu, C.; Williamson, C.; Brink, A.; Hardie, D. Improved Oral Detection Is a Characteristic of Omicron Infection and Has Implications for Clinical Sampling and Tissue Tropism. J. Clin. Virol. 2022, 152, 105170. [Google Scholar] [CrossRef] [PubMed]
- Hui, K.P.Y.; Ng, K.-C.; Ho, J.C.W.; Yeung, H.-W.; Ching, R.H.H.; Gu, H.; Chung, J.C.K.; Chow, V.L.Y.; Sit, K.-Y.; Hsin, M.K.Y.; et al. Replication of SARS-CoV-2 Omicron BA.2 Variant in Ex Vivo Cultures of the Human Upper and Lower Respiratory Tract. eBioMedicine 2022, 83, 104232. [Google Scholar] [CrossRef]
- Lin, J.; Frediani, J.K.; Damhorst, G.L.; Sullivan, J.A.; Westbrook, A.; McLendon, K.; Baugh, T.J.; O’Sick, W.H.; Roback, J.D.; Piantadosi, A.L.; et al. Where is Omicron? Comparison of SARS-CoV-2 RT-PCR and Antigen Test Sensitivity at Commonly Sampled Anatomic Sites over the Course of Disease. Medrxiv 2022. [Google Scholar] [CrossRef]
- Bossuyt, P.M.; Reitsma, J.B.; Bruns, D.E.; Gatsonis, C.A.; Glasziou, P.P.; Irwig, L.; Lijmer, J.G.; Moher, D.; Rennie, D.; de Vet, H.C.W.; et al. STARD 2015: An Updated List of Essential Items for Reporting Diagnostic Accuracy Studies. BMJ 2015, 351, h5527. [Google Scholar] [CrossRef]
- Schrödinger, L.; DeLano, W. PyMol. 2020. Available online: http://www.pymol.org/pymol (accessed on 30 December 2023).
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.H.; Pires, D.E.; Ascher, D.B. DynaMut: Predicting the Impact of Mutations on Protein Conformation, Flexibility and Stability. Nucleic Acids Res. 2018, 46, W350–W355. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Lu, H.; Zhang, N.; Zhu, Z.; Wang, S.; Li, M. PremPS: Predicting the Impact of Missense Mutations on Protein Stability. PLoS Comput. Biol. 2020, 16, e1008543. [Google Scholar] [CrossRef]
- R Foundation for Statistical Computing, Vienna, Austria. R: A Language and Environment for Statistical Computing. 2022. Available online: https://www.R-project.org/ (accessed on 16 April 2024).
- EpiCentro Sorveglianza Integrata COVID-19: I Principali Dati Nazionali. Available online: https://www.epicentro.iss.it/coronavirus/sars-cov-2-sorveglianza-dati (accessed on 31 August 2023).
- Ye, Q.; Lu, S.; Corbett, K.D. Structural Basis for SARS-CoV-2 Nucleocapsid Protein Recognition by Single-Domain Antibodies. Front. Immunol. 2021, 12, 719037. [Google Scholar] [CrossRef]
- Wu, C.; Qavi, A.J.; Hachim, A.; Kavian, N.; Cole, A.R.; Moyle, A.B.; Wagner, N.D.; Sweeney-Gibbons, J.; Rohrs, H.W.; Gross, M.L.; et al. Characterization of SARS-CoV-2 Nucleocapsid Protein Reveals Multiple Functional Consequences of the C-Terminal Domain. iScience 2021, 24, 102681. [Google Scholar] [CrossRef]
- Cubuk, J.; Alston, J.J.; Incicco, J.J.; Singh, S.; Stuchell-Brereton, M.D.; Ward, M.D.; Zimmerman, M.I.; Vithani, N.; Griffith, D.; Wagoner, J.A.; et al. The SARS-CoV-2 Nucleocapsid Protein is Dynamic, Disordered, and Phase Separates with RNA. Nat. Commun. 2021, 12, 1936. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhi, H.; Teng, Y. The Outbreak of SARS-CoV-2 Omicron Lineages, Immune Escape, and Vaccine Effectivity. J. Med. Virol. 2023, 95, e28138. [Google Scholar] [CrossRef] [PubMed]
- Schuit, E.; Venekamp, R.P.; Hooft, L.; Veldhuijzen, I.K.; Van Den Bijllaardt, W.; Pas, S.D.; Zwart, V.F.; Lodder, E.B.; Hellwich, M.; Koppelman, M.; et al. Diagnostic Accuracy of Covid-19 Rapid Antigen Tests with Unsupervised Self-Sampling in People with Symptoms in the Omicron Period: Cross Sectional Study. BMJ 2022, 378, e071215. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Balda, S.; Apreja, M.; Kataria, K.; Capalash, N.; Sharma, P. COVID-19 Diagnosis: Current and Future Techniques. Int. J. Biol. Macromol. 2021, 193, 1835–1844. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, A.; Amarilla, A.A.; Aguado, J.; Modhiran, N.; Albornoz, E.A.; Baradar, A.A.; McMillan, C.L.D.; Choo, J.J.Y.; Idris, A.; Supramaniam, A.; et al. Nucleocapsid Specific Diagnostics for the Detection of Divergent SARS-CoV-2 Variants. Front. Immunol. 2022, 13, 926262. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.Y.; Marais, G.; Opperman, C.; Doolabh, D.; Iranzadeh, A.; Marais, C.; Cox, H.; Williamson, C.; Hardie, D.; Brink, A. Performance of Saliva and Mid-Turbinate Swabs for Detection of the Beta Variant in South Africa. Lancet Infect. Dis. 2021, 21, 1354. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.-P.; Jennings, R.; Hart, B.; Cangelosi, G.A.; Wood, R.C.; Wehber, K.; Verma, P.; Vojta, D.; Berke, E.M. Swabs Collected by Patients or Health Care Workers for SARS-CoV-2 Testing. N. Engl. J. Med. 2020, 383, 494–496. [Google Scholar] [CrossRef]
- Venekamp, R.P.; Schuit, E.; Hooft, L.; Veldhuijzen, I.K.; van den Bijllaardt, W.; Pas, S.D.; Zwart, V.F.; Lodder, E.B.; Hellwich, M.; Koppelman, M.; et al. Diagnostic accuracy of SARS-CoV-2 rapid antigen self-tests in asymptomatic individuals in the omicron period: A cross-sectional study. Clin. Microbiol. Infect. 2023, 29, 391.e1–391.e7. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ADT Group | Commercial Name | Manufacture | Assay Type | Target Protein | Sampling Site | Sensitivity | Specificity |
|---|---|---|---|---|---|---|---|
| 1 | STANDARD Q COVID-19 Ag Test 2.0 | SD Biosensor, Inc. Korea | Lateral Flow, Immuno-chromatography Rapid ADT | N | Nasal swab | 94.94% | 100% |
| 1 | Panbio™ COVID-19 Ag Rapid Test Device | Abbott Rapid Diagnostics Jena GmbH, Germany | Lateral Flow, Immuno-chromatography Rapid ADT | N | Nasal swab | 98.1% | 99.8% |
| 1 | Green Spring SARS-CoV-2 Antigen Rapid Test Kit | Shenzhen Lvshiyuan Biotechnology Co. Ltd., China | Lateral Flow, Immuno-chromatography Rapid ADT | N | Nasal swab | 96.77% | 100% |
| 2 | FREND™ COVID-19 Ag | NanoEntek, South Korea | Microfluidic-based rapid ADT | N | Nasophar. Swab | 94.12% | 94.12% |
| 3 | MAGLUMI® SARS-CoV-2 Ag | Shenzhen New Industries Biomedical Engineering Co., Ltd. China | Laboratory-chemiluminescence-based ADT | N | Nasophar. Swab | 97.7% | 99.6% |
| 3 | LIAISON® SARS-CoV-2 Ag | DiaSorin, Inc | Laboratory chemiluminescence-based ADT | N | Nasal swab | 99.0% | 98.0% |
| Delta Wave | Omicron Wave | ||||
|---|---|---|---|---|---|
| Demographics | Count (n) | Value (%) | Count (n) | Value (%) | |
| Population | 2726 | 2449 | |||
| Gender | |||||
| Female | 1319 | 48.38 | 1216 | 49.65 | |
| Male | 1407 | 51.61 | 1233 | 50.34 | |
| Age (years) | Female | Male | Female | Male | |
| Lower 95% CI | 46.71 | 46.81 | 39.01 | 41.20 | |
| Upper 95% CI | 49.68 | 49.71 | 42.02 | 44.33 | |
| Median | 48.00 | 52.00 | 37.00 | 41.00 | |
| Delta | Omicron | Delta | Omicron | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TP/(TP + FN) | SE (95% CI) | TP/(TP + FN) | SE (95% CI) | p | TN/(TN + FP) | SP (95% CI) | TN/(TN + FP) | SP (95% CI) | p | |
| Overall | 122/192 | 63.5 (56.3, 74.0) | 65/195 | 33.3 (26.8, 44.0) | <0.001 | 2512/2534 | 99.1 (98.7, 99.5) | 2253/2254 | 99.9 (99.8, 100) | <0.001 |
| Group 1 | 27/54 | 50 (36.1, 63.9) | 32/96 | 33.3 (24.0, 43.7) | 0.045 | 549/550 | 99.8 (99.0, 100) | 1496/1497 | 99.9 (99.6, 100) | 0.460 |
| Group 2 | 73/112 | 65.2 (55.6, 73.9) | 22/62 | 35.5 (23.7, 48.7) | <0.001 | 1822/1838 | 99.1 (98.6, 99.5) | 614/614 | 100 (99.4, 100) | 0.020 |
| Group 3 | 22/26 | 84.6 (65.1, 95.6) | 11/37 | 29.7 (15.9, 47.0) | <0.001 | 141/146 | 96.6 (92.2, 98.9) | 143/143 | 100 (97.5, 100) | 0.026 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piubelli, C.; Treggiari, D.; Lavezzari, D.; Deiana, M.; Dishnica, K.; Tosato, E.M.S.; Mazzi, C.; Cattaneo, P.; Mori, A.; Pomari, E.; et al. Wide Real-Life Data Support Reduced Sensitivity of Antigen Tests for Omicron SARS-CoV-2 Infections. Viruses 2024, 16, 657. https://doi.org/10.3390/v16050657
Piubelli C, Treggiari D, Lavezzari D, Deiana M, Dishnica K, Tosato EMS, Mazzi C, Cattaneo P, Mori A, Pomari E, et al. Wide Real-Life Data Support Reduced Sensitivity of Antigen Tests for Omicron SARS-CoV-2 Infections. Viruses. 2024; 16(5):657. https://doi.org/10.3390/v16050657
Chicago/Turabian StylePiubelli, Chiara, Davide Treggiari, Denise Lavezzari, Michela Deiana, Klevia Dishnica, Emma Maria Sole Tosato, Cristina Mazzi, Paolo Cattaneo, Antonio Mori, Elena Pomari, and et al. 2024. "Wide Real-Life Data Support Reduced Sensitivity of Antigen Tests for Omicron SARS-CoV-2 Infections" Viruses 16, no. 5: 657. https://doi.org/10.3390/v16050657