
No Remdesivir Resistance Observed in the Phase 3 Severe and Moderate COVID-19 SIMPLE Trials
 ,
,  , , , , , , , , , , , ,
, , , , , , , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. SIMPLE Study Designs
2.2. Virology Data Collection
2.3. Sequencing Methodologies
2.4. Sequencing Data Processing
2.5. SARS-CoV-2 Lineage Determination
2.6. SARS-CoV-2 Sequence Analysis
2.7. SARS-CoV-2 Phenotypic Analysis
3. Results
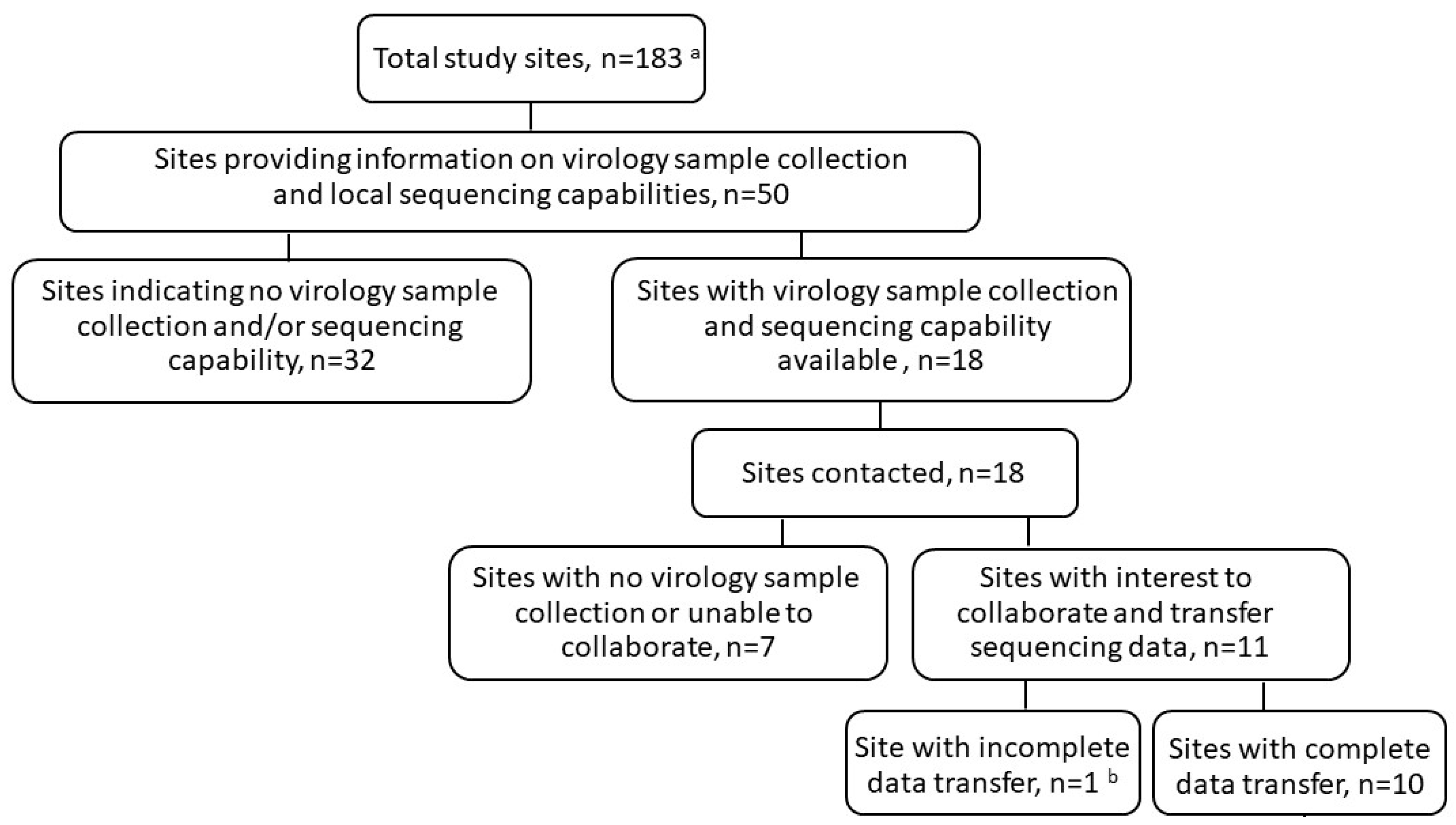
3.1. Sequencing Data Collection
3.2. Lineage and Baseline Nsp12 Substitutions
3.3. Post-Baseline Amino Acid Substitutions in SARS-CoV-2 Nsp12
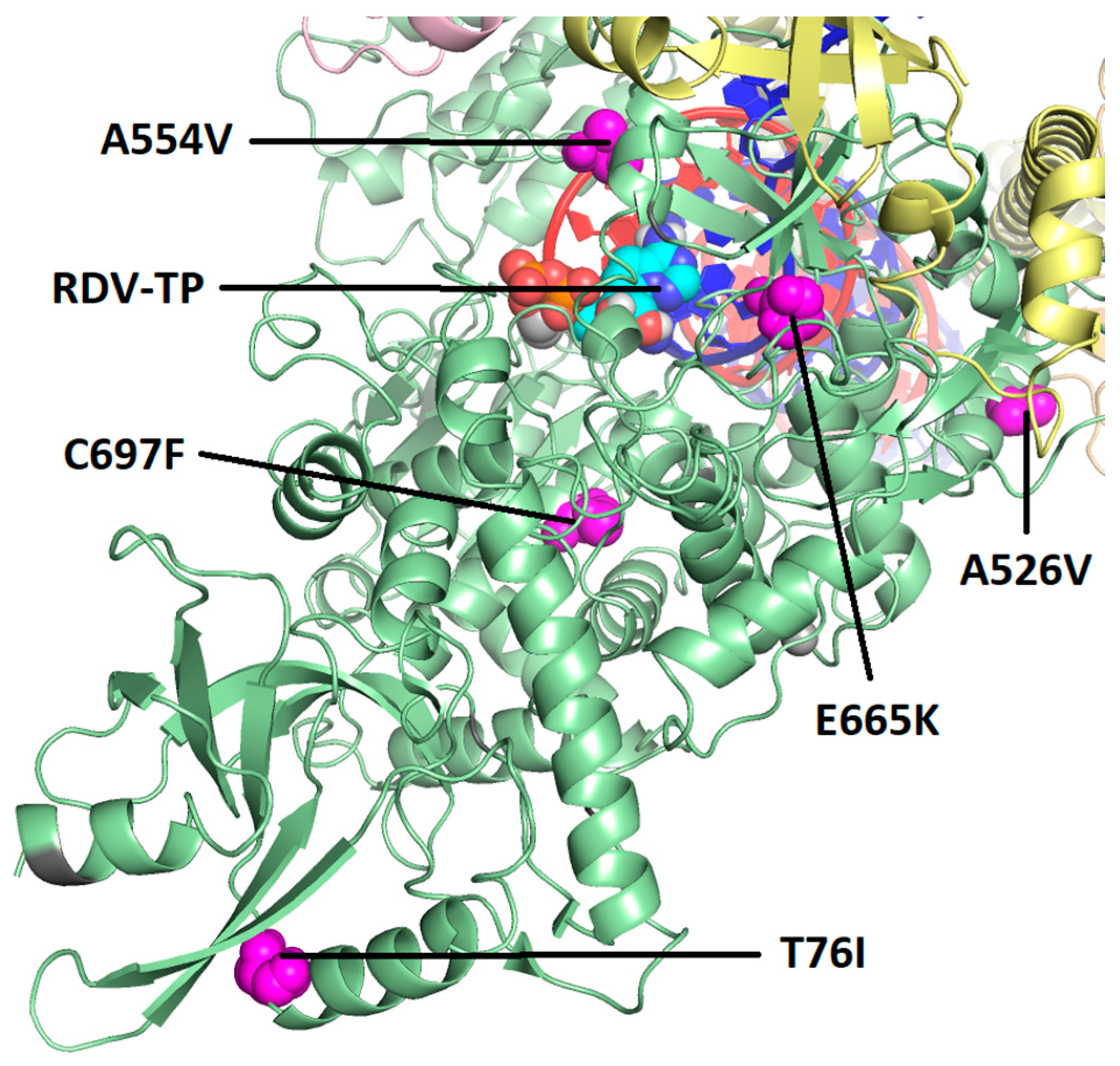
3.4. Phenotypic Analysis of Emergent Nsp12 Substitutions
3.5. Post-Baseline Amino Acid Substitutions Emerging in Other Proteins of the Polymerase Complex
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kokic, G.; Hillen, H.S.; Tegunov, D.; Dienemann, C.; Seitz, F.; Schmitzova, J.; Farnung, L.; Siewert, A.; Höbartner, C.; Cramer, P. Mechanism of SARS-CoV-2 polymerase stalling by remdesivir. Nat. Commun. 2021, 12, 279. [Google Scholar] [CrossRef]
- Bravo, J.P.K.; Dangerfield, T.L.; Taylor, D.W.; Johnson, K.A. Remdesivir is a delayed translocation inhibitor of SARS-CoV-2 replication. Mol. Cell 2021, 81, 1548–1552. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.J.; Tchesnokov, E.P.; Woolner, E.; Perry, J.K.; Feng, J.Y.; Porter, D.P.; Götte, M. Remdesivir is a direct-acting antiviral that inhibits RNA-dependent RNA polymerase from severe acute respiratory syndrome coronavirus 2 with high potency. J. Biol. Chem. 2020, 295, 6785–6797. [Google Scholar] [CrossRef] [PubMed]
- Lo, M.K.; Jordan, R.; Arvey, A.; Sudhamsu, J.; Shrivastava-Ranjan, P.; Hotard, A.L.; Flint, M.; McMullan, L.K.; Siegel, D.; Clarke, M.O.; et al. GS-5734 and its parent nucleoside analog inhibit Filo-, Pneumo-, and Paramyxoviruses. Sci. Rep. 2017, 7, 43395. [Google Scholar] [CrossRef]
- Sheahan, T.P.; Sims, A.C.; Graham, R.L.; Menachery, V.D.; Gralinski, L.E.; Case, J.B.; Leist, S.R.; Pyrc, K.; Feng, J.Y.; Trantcheva, I.; et al. Broad-spectrum antiviral GS-5734 inhibits both epidemic and zoonotic coronaviruses. Sci. Transl. Med. 2017, 9, eaal3653. [Google Scholar] [CrossRef] [PubMed]
- Warren, T.K.; Jordan, R.; Lo, M.K.; Ray, A.S.; Mackman, R.L.; Soloveva, V.; Siegel, D.; Perron, M.; Bannister, R.; Hui, H.C.; et al. Therapeutic efficacy of the small molecule GS-5734 against Ebola virus in rhesus monkeys. Nature 2016, 531, 381–385. [Google Scholar] [CrossRef]
- Beigel, J.H.; Tomashek, K.M.; Dodd, L.E.; Mehta, A.K.; Zingman, B.S.; Kalil, A.C.; Hohmann, E.; Chu, H.Y.; Luetkemeyer, A.; Kline, S.; et al. Remdesivir for the treatment of COVID-19–Final report. N. Engl. J. Med. 2020, 383, 1813–1826. [Google Scholar] [CrossRef]
- Goldman, J.D.; Lye, D.C.B.; Hui, D.S.; Marks, K.M.; Bruno, R.; Montejano, R.; Spinner, C.D.; Galli, M.; Ahn, M.Y.; Nahass, R.G.; et al. Remdesivir for 5 or 10 days in patients with severe COVID-19. N. Engl. J. Med. 2020, 383, 1827–1837. [Google Scholar] [CrossRef]
- Spinner, C.D.; Gottlieb, R.L.; Criner, G.J.; Arribas López, J.R.; Cattelan, A.M.; Soriano Viladomiu, A.; Ogbuagu, O.; Malhotra, P.; Mullane, K.M.; Castagna, A.; et al. Effect of remdesivir vs standard care on clinical status at 11 days in patients with moderate COVID-19: A randomized clinical trial. JAMA 2020, 324, 1048–1057. [Google Scholar] [CrossRef]
- Gottlieb, R.L.; Vaca, C.E.; Paredes, R.; Mera, J.; Webb, B.J.; Perez, G.; Oguchi, G.; Ryan, P.; Nielsen, B.U.; Brown, M.; et al. Early remdesivir to prevent progression to severe COVID-19 in outpatients. N. Engl. J. Med. 2022, 386, 305–315. [Google Scholar] [CrossRef]
- Stevens, L.J.; Pruijssers, A.J.; Lee, H.W.; Gordon, C.J.; Tchesnokov, E.P.; Gribble, J.; George, A.S.; Hughes, T.M.; Lu, X.; Li, J.; et al. Mutations in the SARS-CoV-2 RNA dependent RNA polymerase confer resistance to remdesivir by distinct mechanisms. Sci. Transl. Med. 2022, eabo0718. [Google Scholar] [CrossRef]
- Checkmahomed, L.; Carbonneau, J.; Du Pont, V.; Riola, N.C.; Perry, J.K.; Li, J.; Paré, B.; Simpson, S.M.; Smith, M.A.; Porter, D.P.; et al. In vitro selection of remdesivir-resistant SARS-CoV-2 demonstrates high barrier to resistance. Antimicrob. Agents Chemother. 2022, 66, e0019822. [Google Scholar] [CrossRef] [PubMed]
- Szemiel, A.M.; Merits, A.; Orton, R.J.; MacLean, O.A.; Pinto, R.M.; Wickenhagen, A.; Lieber, G.; Turnbull, M.L.; Wang, S.; Furnon, W.; et al. In vitro selection of remdesivir resistance suggests evolutionary predictability of SARS-CoV-2. PLoS Pathog. 2021, 17, e1009929. [Google Scholar] [CrossRef] [PubMed]
- Carbo, E.C.; Mourik, K.; Boers, S.A.; Munnink, B.O.; Nieuwenhuijse, D.; Jonges, M.; Welkers, M.R.A.; Matamoros, S.; van Harinxma Thoe Slooten, J.; Kraakman, M.E.M.; et al. A comparison of five Illumina, Ion Torrent, and nanopore sequencing technology-based approaches for whole genome sequencing of SARS-CoV-2. Eur. J. Clin. Microbiol. Infect. Dis. 2023, 42, 701–713. [Google Scholar] [CrossRef] [PubMed]
- Quick, J. nCoV-2019 Sequencing Protocol v3 (LoCost) V.3. Protocols.io. 2020. Available online: https://www.protocols.io/view/ncov-2019-sequencing-protocol-v3-locost-bp2l6n26rgqe/v3 (accessed on 1 May 2022).
- Tyson, J.R.; James, P.; Stoddart, D.; Sparks, N.; Wickenhagen, A.; Hall, G.; Choi, J.H.; Lapointe, H.; Kamelian, K.; Smith, A.D.; et al. Improvements to the ARTIC multiplex PCR method for SARS-CoV-2 genome sequencing using nanopore. bioRxiv 2020. bioRxiv: 2020.09.04.283077. [Google Scholar] [CrossRef]
- Gonzalez-Reiche, A.S.; Hernandez, M.M.; Sullivan, M.J.; Ciferri, B.; Alshammary, H.; Obla, A.; Fabre, S.; Kleiner, G.; Polanco, J.; Khan, Z.; et al. Introductions and early spread of SARS-CoV-2 in the New York City area. Science 2020, 369, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.P.; Stromberg, M.P.; Ward, A.; Stewart, C.; Garrison, E.P.; Marth, G.T. MOSAIK: A hash-based algorithm for accurate next-generation sequencing short-read mapping. PLoS ONE 2014, 9, e90581. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Muruato, A.E.; Zhang, X.; Lokugamage, K.G.; Fontes-Garfias, C.R.; Zou, J.; Liu, J.; Ren, P.; Balakrishnan, M.; Cihlar, T.; et al. A nanoluciferase SARS-CoV-2 for rapid neutralization testing and screening of anti-infective drugs for COVID-19. Nat. Commun. 2020, 11, 5214. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Song, W.; Chen, S.; Yuan, Z.; Yi, Z. A bacterial artificial chromosome (BAC)-vectored noninfectious replicon of SARS-CoV-2. Antiviral Res. 2021, 185, 104974. [Google Scholar] [CrossRef]
- Pitts, J.; Li, J.; Perry, J.K.; Du Pont, V.; Riola, N.; Rodriquez, L.; Lu, X.; Kurhade, C.; Xie, X.; Camus, G.; et al. Remdesivir and GS-441524 retain antiviral activity against Delta, Omicron, and other emergent SARS-CoV-2 variants. Antimicrob. Agents Chemother. 2022, 66, e0022222. [Google Scholar] [CrossRef]
- Malone, B.; Perry, J.K.; Olinares, P.D.B.; Lee, H.W.; Chen, J.; Appleby, T.C.; Feng, J.Y.; Bilello, J.P.; Ng, H.; Sotiris, J.; et al. Structural basis for substrate selection by the SARS-CoV-2 replicase. Nature 2023, 614, 781–787. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Li, J.; Parvangada, A.; Perry, J.; Cihlar, T.; Mo, H.; Porter, D.; Svarovskaia, E. Genetic conservation of SARS-CoV-2 RNA replication complex in globally circulating isolates and recently emerged variants from humans and minks suggests minimal pre-existing resistance to remdesivir. Antiviral Res. 2021, 188, 105033. [Google Scholar] [CrossRef] [PubMed]
- Fontanet, A.; Autran, B.; Lina, B.; Kieny, M.P.; Karim, S.S.A.; Sridhar, D. SARS-CoV-2 variants and ending the COVID-19 pandemic. Lancet 2021, 397, 952–954. [Google Scholar] [CrossRef] [PubMed]
- Mahase, E. COVID-19: What new variants are emerging and how are they being investigated? BMJ 2021, 372, n158. [Google Scholar] [CrossRef] [PubMed]
- van Oosterhout, C.; Hall, N.; Ly, H.; Tyler, K.M. COVID-19 evolution during the pandemic-Implications of new SARS-CoV-2 variants on disease control and public health policies. Virulence 2021, 12, 507–508. [Google Scholar] [CrossRef] [PubMed]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; COVID-19 Genomics UK (COG-UK) Consortium; et al. SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef] [PubMed]
- Bojkova, D.; Widera, M.; Ciesek, S.; Wass, M.N.; Michaelis, M.; Cinatl, J., Jr. Reduced interferon antagonism but similar drug sensitivity in Omicron variant compared to Delta variant SARS-CoV-2 isolates. Cell Res. 2022, 32, 319–321. [Google Scholar] [CrossRef] [PubMed]
- Vangeel, L.; De Jonghe, S.; Maes, P.; Slechten, B.; Raymenants, J.; André, E.; Leyssen, P.; Neyts, J.; Jochmans, D. Remdesivir, molnupiravir and nirmatrelvir remain active against SARS-CoV-2 Omicron and other variants of concern. Antiviral Res. 2022, 198, 105252. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.M.; Katz, M.J.; Ginde, A.A.; Juneja, K.; Ramchandani, M.; Schiffer, J.T.; Vaca, C.; Gottlieb, R.L.; Tian, Y.; Elboudwarej, E.; et al. Consistent effects of early remdesivir on symptoms and disease progression across at-risk outpatient subgroups: Treatment effect heterogeneity in PINETREE study. Infect. Dis. Ther. 2023, 12, 1189–1203. [Google Scholar] [CrossRef]
- Mackman, R.L.; Kalla, R.V.; Babusis, D.; Pitts, J.; Barrett, K.T.; Chun, K.; Du Pont, V.; Rodriguez, L.; Moshiri, J.; Xu, Y.; et al. Discovery of GS-5245 (obeldesivir), an oral prodrug of nucleoside GS-441524 that exhibits antiviral efficacy in SARS-CoV-2-infected African green monkeys. J. Med. Chem. 2023, 66, 11701–11717. [Google Scholar] [CrossRef]
- Focosi, D.; Maggi, F.; McConnell, S.; Casadevall, A. Very low levels of remdesivir resistance in SARS-CoV-2 genomes after 18 months of massive usage during the COVID19 pandemic: A GISAID exploratory analysis. Antiviral Res. 2022, 198, 105247. [Google Scholar] [CrossRef] [PubMed]
- Hedskog, C.; Rodriguez, L.; Roychoudhury, P.; Huang, M.L.; Jerome, K.R.; Hao, L.; Ireton, R.C.; Li, J.; Perry, J.K.; Han, D.; et al. Viral resistance analyses from the remdesivir Phase 3 Adaptive COVID-19 Treatment Trial-1 (ACTT-1). J. Infect. Dis. 2023, 228, 1263–1273. [Google Scholar] [CrossRef] [PubMed]
- Owusu, D.; Pomeroy, M.A.; Lewis, N.M.; Wadhwa, A.; Yousaf, A.R.; Whitaker, B.; Dietrich, E.; Hall, A.J.; Chu, V.; Thornburg, N.; et al. Persistent SARS-CoV-2 RNA shedding without evidence of infectiousness: A cohort study of individuals with COVID-19. J. Infect. Dis. 2021, 224, 1362–1371. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhang, T.; Ren, H.; Sun, S.; Yu, X.; Sheng, J.; Shi, Y.; Zhao, H. Impact of age on duration of viral RNA shedding in patients with COVID-19. Aging 2020, 12, 22399–22404. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Chen, Y.; Yuan, J.; Yi, P.; Ding, C.; Wu, W.; Li, Y.; Ni, Q.; Zou, R.; Li, X.; et al. Factors associated with prolonged viral RNA shedding in patients with coronavirus disease 2019 (COVID-19). Clin. Infect. Dis. 2020, 71, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Showers, W.M.; Leach, S.M.; Kechris, K.; Strong, M. Longitudinal analysis of SARS-CoV-2 spike and RNA-dependent RNA polymerase protein sequences reveals the emergence and geographic distribution of diverse mutations. Infect. Genet. Evol. 2022, 97, 105153. [Google Scholar] [CrossRef] [PubMed]
- Nesterenko, P.A.; McLaughlin, J.; Tsai, B.L.; Burton Sojo, G.; Cheng, D.; Zhao, D.; Mao, Z.; Bangayan, N.J.; Obusan, M.B.; Su, Y.; et al. HLA-A∗02:01 restricted T cell receptors against the highly conserved SARS-CoV-2 polymerase cross-react with human coronaviruses. Cell Rep. 2021, 37, 110167. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, S.; Klein, J.; Robertson, A.; Peña-Hernández, M.A.; Lin, M.J.; Roychoudhury, P.; Lu, P.; Fournier, J.; Ferguson, D.; Mohamed Bakhash, S.A.; et al. De novo emergence of a remdesivir resistance mutation during treatment of persistent SARS-CoV-2 infection in an immunocompromised patient: A case report. Nat. Commun. 2021, 13, 1547. [Google Scholar] [CrossRef]
- Hogan, J.I.; Duerr, R.; Dimartino, D.; Marier, C.; Hochman, S.; Mehta, S.; Wang, G.; Heguy, A. Remdesivir resistance in transplant recipients with persistent COVID-19. Clin. Infect. Dis. 2023, 76, 342–345. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Study Part | Treatment Group | Participants | Participants with Sequencing Data | |||
|---|---|---|---|---|---|---|
| Baseline | Post-Baseline | Both Baseline and Post-Baseline | ||||
| Severe COVID-19 study a | A | RDV for up to 5 days | 200 | 2 | 1 | 0 |
| RDV for up to 10 days | 197 | 1 | 1 | 1 | ||
| B | Non-randomized expanded access, RDV for up to 10 days | 3597 | 51 | 16 | 9 | |
| Invasive mechanical ventilation, RDV for up to 10 days | 844 | 19 | 14 | 9 | ||
| Total | 4838 | 73 | 32 | 19 | ||
| Moderate COVID-19 study b | A | RDV for up to 5 days | 191 | 3 | 0 | 0 |
| RDV for up to 10 days | 193 | 0 | 1 | 0 | ||
| SOC | 200 | 4 | 2 | 2 | ||
| B | Non-randomized expanded access, RDV for up to 10 days | 503 | 7 | 3 | 2 | |
| Total | 1087 | 14 | 6 | 4 | ||
| Number of Participants with Amino Acid Substitutions Occurring in ≥2 Participants at Baseline, n (%) a | ||
|---|---|---|
| Severe COVID-19 Study | Moderate COVID-19 Study | |
| P323L | 50/53 (94%) | 8/9 (88%) |
| A554V | 2/64 (3.1%) | 0/13 (0%) |
| Number of Participants with Emergent Amino Acid Substitutions at Post-Baseline, n (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Severe COVID-19 Study | Moderate COVID-19 Study | |||||||||
| Part A | Part B | Part A | Part B | |||||||
| RDV for up to 5 Days (n = 0) | RDV for up to 10 Days (n = 1) | Non-randomized Expanded Access (RDV for up to 10 Days) (n = 9) | Invasive Mechanical Ventilation (RDV for up to 10 Days) (n = 9) | Total (n = 19) | RDV for up to 5 Days (n = 0) | RDV for up to 10 Days (n = 0) | SOC (n = 2) | Non-Randomized Expanded Access (RDV for up to 10 Days) (n = 2) | Total in RDV Arms (n = 2) | |
| T76T/I | NA | 0 | 1/9 (11%) | 0 | 1/19 (5%) | NA | NA | 0 | 0 | 0 |
| A526V | NA | 0 | 0 | 1/9 (11%) | 1/19 (5%) | NA | NA | 0 | 0 | 0 |
| A554V a | NA | 0 | 0 | 1/9 (11%) a | 1/19 (5%) | NA | NA | 0 | 0 | 0 |
| E665K a | NA | 0 | 0 | 1/9 (11%) a | 1/19 (5%) | NA | NA | 0 | 0 | 0 |
| C697C/F | NA | 0 | 1/9 (11%) | 0 | 1/19 (5%) | NA | NA | 0 | 0 | 0 |
| Substitution in Nsp12 | RDV EC50 (nM) | EC50 Fold Change ± SD from WA1 | ||||
|---|---|---|---|---|---|---|
| 1st Replicate | 2nd Replicate | 3rd Replicate | 4th Replicate | Mean ± SD | ||
| Wildtype (SH01) | 8.83 | 8.44 | 9.51 | 13.03 | 9.95 ± 2.10 | 1.00 |
| T76I | - | - | 7.16 | 9.48 | 8.32 ± 1.63 | 0.84 |
| A526V | 10.69 | 13.08 | - | - | 11.89 ± 1.69 | 1.19 |
| A554V | 14.67 | 14.67 | - | - | 14.45 ± 0.31 | 1.45 |
| E665K | No replication | NA | NA | |||
| C697F | 10.33 | 9.43 | - | - | 9.88 ± 0.64 | 0.99 |
| Change from Baseline | Number of Participants with Baseline and Post-Baseline Sequencing Data | ||||
|---|---|---|---|---|---|
| A | B | Total (n = 12) | |||
| RDV for up to 5 Days (n = 0) | RDV for up to 10 Days (n = 0) | Non-Randomized Expanded Access (RDV for up to 10 Days) (n = 6) a | Invasive Mechanical Ventilation (RDV for up to 10 Days) (n = 6) | ||
| Nsp8 A89T | NA | NA | 0 | 1/6 (17%) | 1/9 (11%) |
| Nsp13 F581L | NA | NA | 1/4 (25%) | 0 | 1/10 (10%) |
| Nsp14 T21R | NA | NA | 0 | 1/6 (17%) | 1/11 (9%) |
| Nsp14 I101V + A287V | NA | NA | 1/5 (20%) | 0 | 1/11 (9%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hedskog, C.; Spinner, C.D.; Protzer, U.; Hoffmann, D.; Ko, C.; Gottlieb, R.L.; Askar, M.; Roestenberg, M.; de Vries, J.J.C.; Carbo, E.C.; et al. No Remdesivir Resistance Observed in the Phase 3 Severe and Moderate COVID-19 SIMPLE Trials. Viruses 2024, 16, 546. https://doi.org/10.3390/v16040546
Hedskog C, Spinner CD, Protzer U, Hoffmann D, Ko C, Gottlieb RL, Askar M, Roestenberg M, de Vries JJC, Carbo EC, et al. No Remdesivir Resistance Observed in the Phase 3 Severe and Moderate COVID-19 SIMPLE Trials. Viruses. 2024; 16(4):546. https://doi.org/10.3390/v16040546
Chicago/Turabian StyleHedskog, Charlotte, Christoph D. Spinner, Ulrike Protzer, Dieter Hoffmann, Chunkyu Ko, Robert L. Gottlieb, Medhat Askar, Meta Roestenberg, Jutte J. C. de Vries, Ellen C. Carbo, and et al. 2024. "No Remdesivir Resistance Observed in the Phase 3 Severe and Moderate COVID-19 SIMPLE Trials" Viruses 16, no. 4: 546. https://doi.org/10.3390/v16040546