
Effects of Strain Differences, Humidity Changes, and Saliva Contamination on the Inactivation of SARS-CoV-2 by Ion Irradiation
 and
and 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Cultures
2.2. Viruses
2.3. Virus Titration
2.4. Saliva
2.5. Treatment of Viruses with Ions
2.6. Humidity Conditions
2.7. Statistical Analysis
3. Results
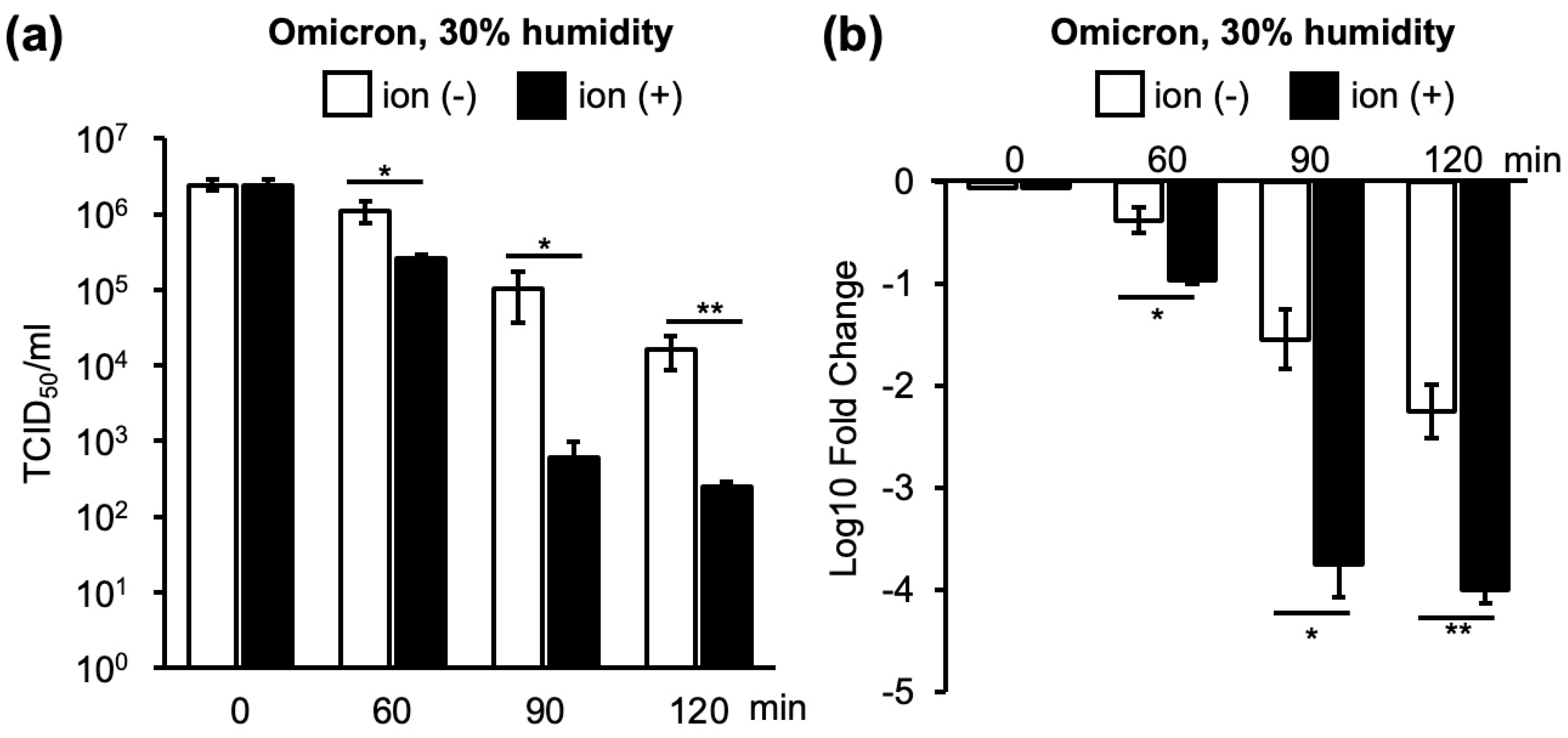
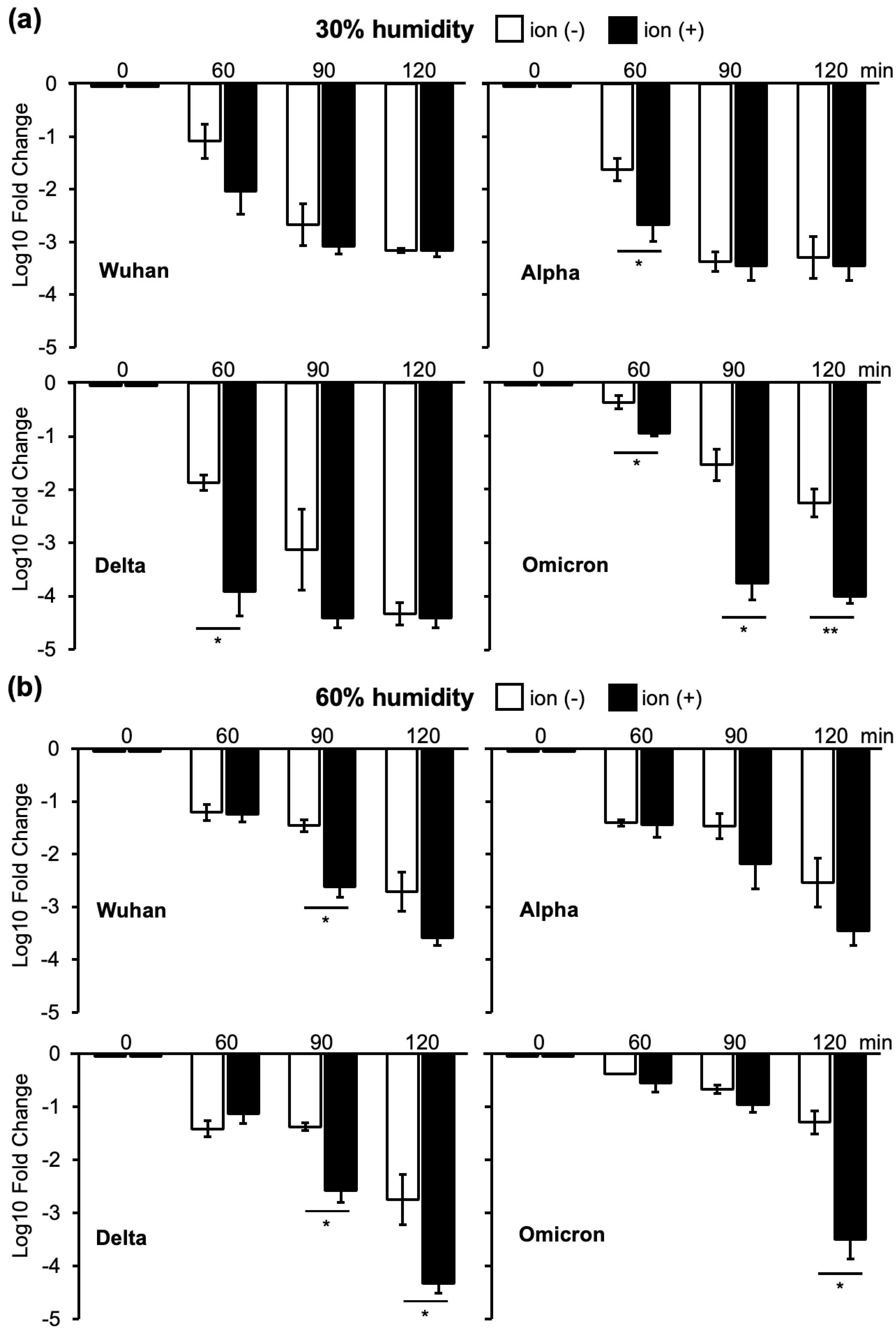
3.1. Changes in Infectivity of Four VOCs as a Result of Ion Irradiation in 30% or 60% Humidity
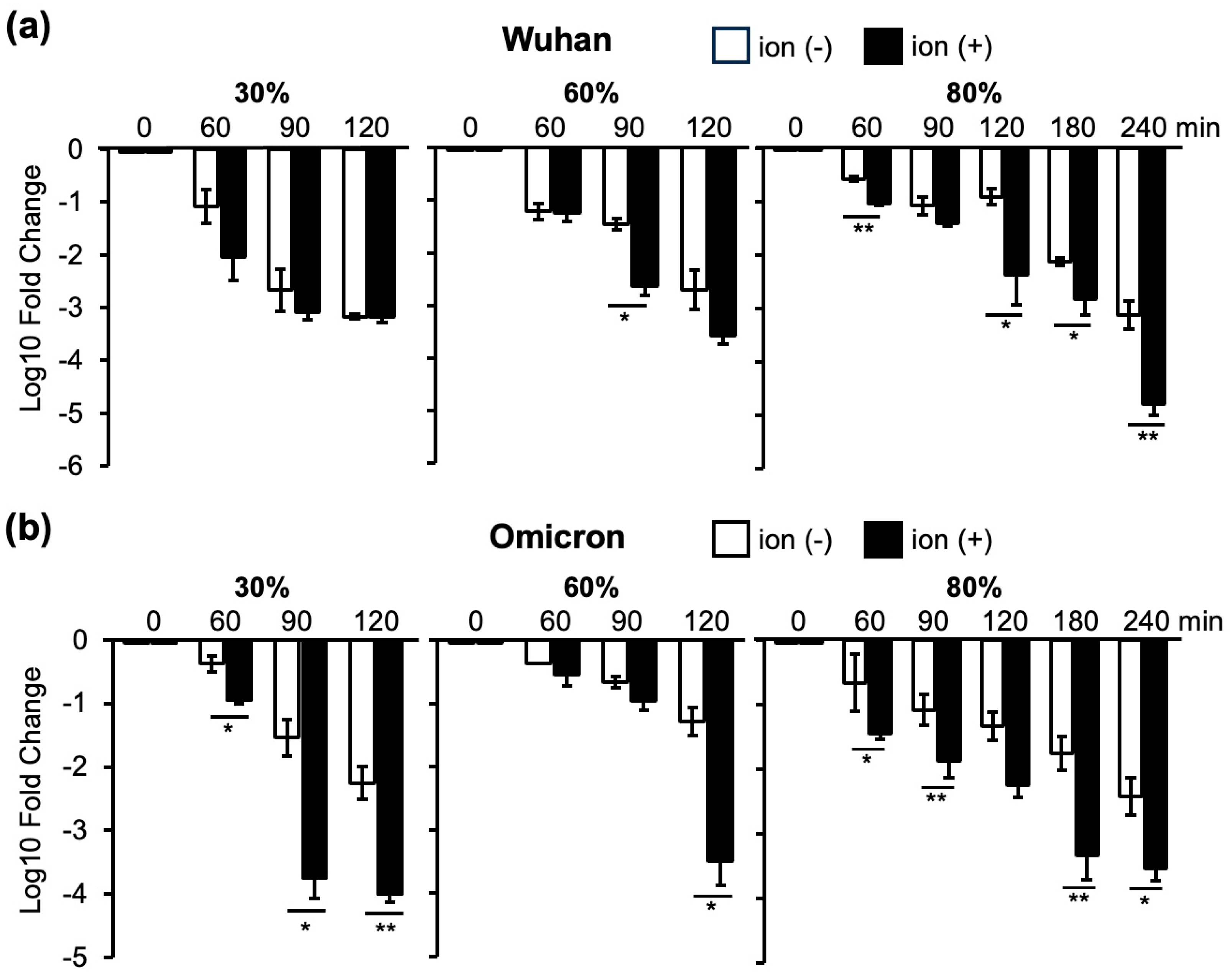
3.2. Changes in Infectivity of the Wuhan and Omicron Strains as a Result of Ion Irradiation in 30%, 60%, and 80% Humidity Environments
3.3. Changes in Infectivity of the Wuhan and Omicron Strains in Saliva at 30%, 60%, and 80% Humidity as a Result of Ion Irradiation
3.4. Changes in Infectivity of the Omicron Strain with or without Saliva as a Result of Ion Irradiation at 30%, 60%, and 80% Humidity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.L. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2021, 19, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, S.A.; Hogue, B.G.; Leibowitz, J.L.; Weiss, S.R. Severe acute respiratory syndrome Coronavirus 2 (SARS-CoV-2). In Fields Virology, 7th ed.; Howley, P.M., Knipe, D.M., Eds.; Wolters Kluwer: Philadelphia, PA, USA, 2022; Volume 2, pp. 706–740. [Google Scholar]
- Eckerle, L.D.; Becker, M.M.; Halpin, R.A.; Li, K.; Venter, E.; Lu, X.; Scherbakova, S.; Graham, R.L.; Baric, R.S.; Stockwell, T.B.; et al. Infidelity of SARS-CoV Nsp14-exonuclease mutant virus replication is revealed by complete genome sequencing. PLoS Pathog. 2010, 6, e1000896. [Google Scholar] [CrossRef]
- Carabelli, A.M.; Peacock, T.P.; Thorne, L.G.; Harvey, W.T.; Hughes, J.; COVID-19 Genomics UK Consortium; Peacock, S.J.; Barclay, W.S.; de Silva, T.I.; Towers, G.J.; et al. SARS-CoV-2 variant biology: Immune escape, transmission and fitness. Nat. Rev. Microbiol. 2023, 21, 162–177. [Google Scholar] [PubMed]
- Markov, P.V.; Ghafari, M.; Beer, M.; Lythgoe, K.; Simmonds, P.; Stilianakis, N.I.; Katzourakis, A. The evolution of SARS-CoV-2. Nat. Rev. Microbiol. 2023, 21, 361–379. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Hilgenfeld, R.; Whitley, R.; De Clercq, E. Therapeutic strategies for COVID-19: Progress and lessons learned. Nat. Rev. Drug Discov. 2023, 22, 449–475. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Q.; Vishwanath, S.; Carnell, G.W.; Chan, A.C.Y.; Heeney, J.L. Immune imprinting and next-generation coronavirus vaccines. Nat. Microbiol. 2023, 8, 1971–1985. [Google Scholar] [CrossRef]
- Fadaei, A. Viral inactivation with emphasis on SARS-CoV-2 using physical and chemical disinfectants. Sci. World J. 2021, 2021, 9342748. [Google Scholar] [CrossRef] [PubMed]
- Aho Glele, L.S.; de Rougemont, A. Non-pharmacological strategies and interventions for effective COVID-19 control: A Narrative Review. J. Clin. Med. 2023, 12, 6465. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.Y.; Ma, A.; Ramachandran, S. Negative air ions and their effects on human health and air quality improvement. Int. J. Mol. Sci. 2018, 19, 2966. [Google Scholar] [CrossRef]
- Seo, K.H.; Mitchell, B.W.; Holt, P.S.; Gast, R.K. Bactericidal effects of negative air ions on airborne and surface Salmonella enteritidis from an artificially generated aerosol. J. Food Prot. 2001, 64, 113–116. [Google Scholar] [CrossRef]
- Kawamoto, H.; Umezu, S. Electrostatic micro-ozone fan that utilizes ionic wind induced in pin-to-plate corona discharge system. J. Electrostat. 2008, 66, 445–454. [Google Scholar] [CrossRef]
- Digel, I.; Temiz Artmann, A.; Nishikawa, K.; Cook, M.; Kurulgan, E.; Artmann, G.M. Bactericidal effects of plasma-generated cluster ions. Med. Biol. Eng. Comput. 2005, 43, 800–807. [Google Scholar] [CrossRef] [PubMed]
- Hiramoto, K.; Orita, K.; Yamate, Y.; Sato, E.F.; Okano, H.; Nishikawa, K.; Inoue, M. Plasma cluster ions decrease the antigenicity of mite allergens and suppress atopic dermatitis in NC/Nga mice. Arch. Dermatol. Res. 2011, 303, 367–370. [Google Scholar] [CrossRef]
- Okano, H.; Fujimura, T.; Fukuoka, N.; Hayashi, T.; Nishikawa, K.; Ono, K.; Kawamoto, S. Plasma cluster ions reduce the IgE-binding capacity of house dust mite allergens under a simulated indoor environmental condition. Int. Arch. Allergy Immunol. 2017, 173, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Verheyen, C.A.; Bourouiba, L. Associations between indoor relative humidity and global COVID-19 outcomes. J. R. Soc. Interface 2022, 19, 20210865. [Google Scholar] [CrossRef]
- Biryukov, J.; Boydston, J.A.; Dunning, R.A.; Yeager, J.J.; Wood, S.; Reese, A.L.; Ferris, A.; Miller, D.; Weaver, W.; Zeitouni, N.E.; et al. Increasing temperature and relative humidity accelerates inactivation of SARS-CoV-2 on surfaces. mSphere 2020, 5, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Pérez, P.; Kato, T.; Mikami, Y.; Okuda, K.; Gilmore, R.C.; Conde, C.D.; Gasmi, B.; Stein, S.; Beach, M.; et al. SARS-CoV-2 infection of the oral cavity and saliva. Nat. Med. 2021, 27, 892–903. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Caballero, M.; Davis, R.D.; Fuques, E.; Gomez, O.M.; Huynh, E.; Handorean, A.; Ushijima, S.; Tolbert, M.; Hernandez, M. Carbohydrate vitrification in aerosolized saliva is associated with the humidity-dependent infectious potential of airborne coronavirus. PNAS Nexus 2022, 2, pgac301. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, S.; Nao, N.; Shirato, K.; Kawase, M.; Saito, S.; Takayama, I.; Nagata, N.; Sekizuka, T.; Katoh, H.; Kato, F.; et al. Enhanced isolation of SARS-CoV-2 by TMPRSS2-expressing cells. Proc. Natl. Acad. Sci. USA 2020, 117, 7001–7003. [Google Scholar] [CrossRef]
- Laxton, C.S.; Peno, C.; Hahn, A.M.; Allicock, O.M.; Perniciaro, S.; Wyllie, A.L. The potential of saliva as an accessible and sensitive sample type for the detection of respiratory pathogens and host immunity. Lancet Microbe 2023, 4, e837–e850. [Google Scholar] [CrossRef]
- Lin, G.; Hamilton, A.; Gatalo, O.; Haghpanah, F.; Igusa, T.; Klein, E. Investigating the effects of absolute humidity and movement on COVID-19 seasonality in the United States. Sci. Rep. 2022, 12, 16729. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Lee, S.G.; Oh, J.S.; Nam, M.; Barrett, S.; Lee, S.; Hwang, W. The effects of indoor temperature and humidity on local transmission of COVID-19 and how it relates to global trends. PLoS ONE 2022, 17, e0271760. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Dong, Z.; Fan, W.; Xu, K.; Tang, S.; Wang, Y.; Wu, F. A narrative review on the role of temperature and humidity in COVID-19: Transmission, persistence, and epidemiological evidence. Eco-Environ. Health 2022, 1, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Casanova, L.M.; Jeon, S.; Rutala, W.A.; Weber, D.J.; Sobsey, M.D. Effects of air temperature and relative humidity on coronavirus survival on surfaces. Appl. Environ. Microbiol. 2010, 76, 2712–2717. [Google Scholar] [CrossRef] [PubMed]
- Morris, D.H.; Yinda, K.C.; Gamble, A.; Rossine, F.W.; Huang, Q.; Bushmaker, T.; Fischer, R.J.; Matson, M.J.; Van Doremalen, N.; Vikesland, P.J.; et al. Mechanistic theory predicts the effects of temperature and humidity on inactivation of SARS-CoV-2 and other enveloped viruses. ELife 2021, 10, e65902. [Google Scholar] [CrossRef] [PubMed]
- To, K.K.; Tsang, O.T.; Yip, C.C.; Chan, K.H.; Wu, T.C.; Chan, J.M.; Leung, W.S.; Chik, T.S.; Choi, C.Y.; Kandamby, D.H.; et al. Consistent detection of 2019 novel Coronavirus in saliva. Clin. Infect. Dis. 2020, 71, 841–843. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Yanagi, U.; Kagi, N.; Kim, H.; Ogata, M.; Hayashi, M. Environmental factors involved in SARS-CoV-2 transmission: Effect and role of indoor environmental quality in the strategy for COVID-19 infection control. Environ. Health Prev. Med. 2020, 25, 66. [Google Scholar] [CrossRef]
- Hirose, R.; Itoh, Y.; Ikegaya, H.; Miyazaki, H.; Watanabe, N.; Yoshida, T.; Bandou, R.; Daidoji, T.; Nakaya, T. Differences in environmental stability among SARS-CoV-2 variants of concern: Both Omicron BA.1 and BA.2 have higher stability. Clin. Microbiol. Infect. 2022, 11, 1486–1491. [Google Scholar] [CrossRef]
- Chin, A.W.H.; Lai, A.M.Y.; Peiris, M.; Man Poon, L.L. Increased stability of SARS-CoV-2 omicron variant over ancestral strain. Emerg. Infect. Dis. 2022, 28, 1515–1517. [Google Scholar] [CrossRef]
- Ye, G.; Liu, B.; Li, F. Cryo-EM structure of a SARS-CoV-2 omicron spike protein ectodomain. Nat. Commun. 2022, 13, 1214. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad Wadi, A.F.A.; Onomura, D.; Funamori, H.; Khatun, M.M.; Okada, S.; Iizasa, H.; Yoshiyama, H. Effects of Strain Differences, Humidity Changes, and Saliva Contamination on the Inactivation of SARS-CoV-2 by Ion Irradiation. Viruses 2024, 16, 520. https://doi.org/10.3390/v16040520
Ahmad Wadi AFA, Onomura D, Funamori H, Khatun MM, Okada S, Iizasa H, Yoshiyama H. Effects of Strain Differences, Humidity Changes, and Saliva Contamination on the Inactivation of SARS-CoV-2 by Ion Irradiation. Viruses. 2024; 16(4):520. https://doi.org/10.3390/v16040520
Chicago/Turabian StyleAhmad Wadi, Afifah Fatimah Azzahra, Daichi Onomura, Hirokazu Funamori, Mst Mahmuda Khatun, Shunpei Okada, Hisashi Iizasa, and Hironori Yoshiyama. 2024. "Effects of Strain Differences, Humidity Changes, and Saliva Contamination on the Inactivation of SARS-CoV-2 by Ion Irradiation" Viruses 16, no. 4: 520. https://doi.org/10.3390/v16040520