
Characterization of the RNA Mycovirome Associated with Grapevine Fungal Pathogens: Analysis of Mycovirus Distribution and Their Genetic Variability within a Collection of Botryosphaeriaceae Isolates
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions and Characterization of Botryosphaeriaceae Isolates
2.2. Determination of Fungal Isolates’ In Vitro Growth Rates
2.3. Pathogenicity Assays: Length of Necrosis
2.4. Double-Stranded RNA Extraction and High-Throughput Sequencing Analysis
2.5. Completion of Genomic Sequences
2.6. Sequence and Phylogenetic Analyses
2.7. Total Nucleic Acid (TNA) Extraction and the Detection of Mycoviruses by RT-PCR
3. Results
3.1. RNA Virome Associated to Botryosphaeriaceae Species
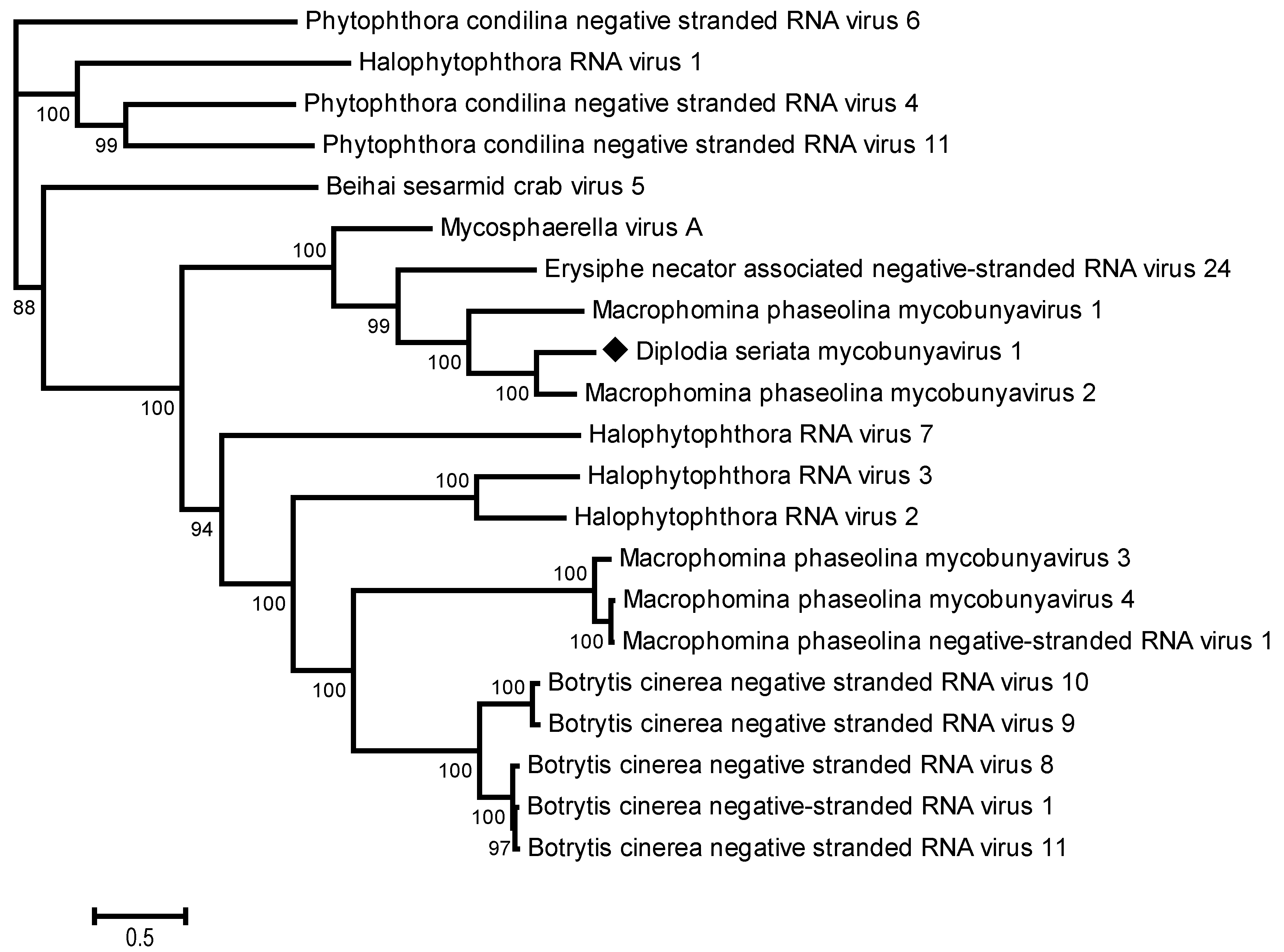
3.2. Molecular Features and Phylogenetic Relationships of the Identified Novel Viruses
3.2.1. A New Fusagravirus in D. mutila and N. parvum
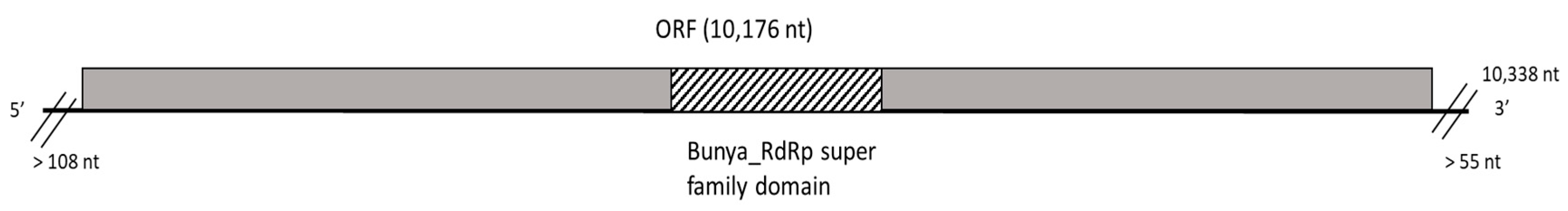
3.2.2. A New Mycobunyavirus in D. seriata
3.2.3. A New Narnavirus in D. seriata
3.3. Distribution of Mycoviruses within a Collection of Botryosphaeriaceae Isolates, and Analysis of the Genetic Diversity of the Identified Viruses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Phillips, A.J.L.; Hyde, K.D.; Alves, A.; Liu, J.K.J. Families in Botryosphaeriales: A phylogenetic, morphological and evolutionary perspective. Fungal Divers. 2019, 94, 1e22. [Google Scholar] [CrossRef]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Hrycan, J.; Hart, M.; Bowen, P.; Forge, T.; Úrbez-Torres, J.R. Grapevine trunk disease fungi: Their roles as latent pathogens and stress factors that favour disease development and symptom expression. Phytopathol. Medit. 2020, 59, 395–424. [Google Scholar] [CrossRef]
- Fontaine, F.; Gramaje, D.; Armengol, J.; Smart, R.; Nagy, Z.A.; Borgo, M.; Rego, C.; Corio-Costet, M.F. Grapevine Trunk Disease, a Review; OIV Publications: Paris, France, 2016; p. 24. [Google Scholar]
- Mugnai, L.; Graniti, A.; Surico, G. Esca (black measles) and brown wood streaking: Two old and elusive diseases of grapevines. Plant Dis. 1999, 83, 404–418. [Google Scholar] [CrossRef] [PubMed]
- Urbez-Torres, J.R.; Francesca, P.; Striegler, R.K.; Urrea-Romero, K.E.; Rupe, J.C.; Cartwright, R.D.; Gubler, W.D. Characterization of fungal pathogens associated with grapevine trunk diseases in Arkansas and Missouri. Fungal Divers. 2012, 52, 169–189. [Google Scholar] [CrossRef]
- Baskarathevan, J.; Jaspers, M.V.; Jones, E.E.; Ridgway, H.J. Incidence and distribution of botryosphaeriaceous species causing dieback and decline in New Zealand vineyards. Eur. J. Plant Pathol. 2012, 132, 549–560. [Google Scholar] [CrossRef]
- Spetik, M.; Tekielska, D.A.; Berraf-Tebbal, A.; Pecenka, J.; Stuskova, K.; Mahamedi, A.E.; Eichmeier, A. Diversity of Botryosphaeriaceae species associated with grapevine trunk diseases in the Czech Republic. Diversity 2023, 15, 800. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef]
- Comont, G.; Mayet, V.; Corio-Costet, M.F. First report of Lasiodiplodia viticola, Spencermartinsia viticola and Diplodia intermedia associated with Vitis vinifera decline in French vineyards. Plant Dis. 2016, 100, 2328. [Google Scholar] [CrossRef]
- Larignon, P. Maladies du Bois de la Vigne: Le Cycle de Vie des Champignons de la Famille des Botryosphaeriacées; IFV, 2022; Available online: https://www.vignevin.com/wp-content/uploads/2023/01/Fiche-cycle-Botryosphaeriaceesvf_Web.pdf (accessed on 7 December 2023).
- Masi, M.; Cimmino, A.; Reveglia, P.; Mugnai, L.; Surico, G.; Evidente, A. Advances on fungal phytotoxins and their role in grapevine trunk diseases. J. Agric. Food Chem. 2018, 66, 5948–5958. [Google Scholar] [CrossRef]
- Bellée, A.; Comont, G.; Nivault, A.; Abou-Mansour, E.; Coppin, C.; Dufour, M.C.; Corio-Costet, M.F. Life traits of four Botryosphaeriaceae species and molecular responses of different genotypes and cultivar of Vitis vinifera. Plant Pathol. 2017, 66, 763–776. [Google Scholar] [CrossRef]
- Mesguida, O.; Haidar, R.; Yacoub, A.; Dreux-Zigha, A.; Berthon, J.Y.; Guyoneaud, R.; Attard, E.; Rey, P. Microbial biological control of fungi associated with grapevine trunk diseases: A review of strain diversity, modes of action, and advantages and limits of current strategies. J. Fungi 2023, 9, 638. [Google Scholar] [CrossRef]
- Bruez, E.; Lecomte, P.; Grosman, J.; Doublet, B.; Bertsch, C.; Fontaine, F.; Ugaglia, A.; Teissedre, P.L.; Da Costa, J.P.; Guerin-Dubrana, L.; et al. Overview of grapevine trunk diseases in France in the 2000s. Phytopath. Medit. 2013, 52, 262–275. [Google Scholar] [CrossRef]
- Mondello, V.; Songy, A.; Battiston, E.; Pinto, C.; Coppin, C.; Trotel-Aziz, P.; Clement, C.; Mugnai, L.; Fontaine, F. Grapevine trunk diseases: A review of fifteen years of trials for their control with chemicals and biocontrols agents. Plant Dis. 2018, 102, 1189–1217. [Google Scholar] [CrossRef]
- Lambert, C.; Bisson, J.; Waffo-Téguo, P.; Papastamoulis, Y.; Richard, T.; Corio-Costet, M.F.; Mérillon, J.M.; Cluzet, S. Phenolics and their antifungal role in grapevine wood decay: Focus on the Botryosphaeriaceae family. J. Agric. Food. Chem. 2012, 60, 11859–11868. [Google Scholar] [CrossRef]
- Stempien, E.; Goddard, M.L.; Wilhelm, K.; Tarnus, C.; Bertsch, C.; Chong, J. Grapevine Botryosphaeria dieback fungi have specific aggressiveness factor repertory involved in wood decay and stilbene metabolization. PLoS ONE 2017, 12, e0188766. [Google Scholar] [CrossRef]
- Dawe, A.L.; Nuss, D.L. Hypoviruses and chestnut blight: Exploiting viruses to understand and modulate fungal pathogenesis. Annu. Rev. Genet. 2001, 35, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479–480, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Kotta-Loizou, I. Mycoviruses and their role in fungal pathogenesis. Curr. Opin. Microbiol. 2021, 63, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Villan Larios, D.C.; Diaz Reyes, B.M.; Pirovani, C.P.; Loguercio, L.L.; Santos, V.C.; Góes-Neto, A.; Fonseca, P.L.C.; Aguiar, E.R.G.R. Exploring the mycovirus universe: Identification, diversity, and biotechnological applications. J. Fungi 2023, 9, 361. [Google Scholar] [CrossRef]
- Hough, B.; Steenkamp, E.; Wingfield, B.; Read, D. Fungal viruses unveiled: A comprehensive review of mycoviruses. Viruses 2023, 15, 1202. [Google Scholar] [CrossRef]
- Ayllón, M.A.; Vainio, E.J. Mycoviruses as a part of the global virome: Diversity, evolutionary links and lifestyle. Adv. Virus. Res. 2023, 115, 1–86. [Google Scholar] [CrossRef]
- Wang, L.; Jiang, J.; Wang, Y.; Hong, N.; Zhang, F.; Xu, W.; Wang, G. Hypovirulence of the phytopathogenic fungus Botryosphaeria dothidea: Association with a coinfecting chrysovirus and a partitivirus. J. Virol. 2014, 88, 7517–7527. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, H.; Cao, J.; Yin, X.; Guo, Y.; Guo, L.; Wu, H.; Zhang, M. Characterization of a novel mycovirus from the phytopathogenic fungus Botryosphaeria dothidea. Viruses 2022, 14, 331. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Xiang, J.; Zhang, M.; Fu, M.; Yang, Z.; Hong, N.; Wang, G. Characterization of a novel double-stranded RNA mycovirus conferring hypovirulence from the phytopathogenic fungus Botryosphaeria dothidea. Virology 2016, 493, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Yang, M.; Zhang, M.; Hong, N.; Wang, G. Characterization of a Botybirnavirus conferring hypovirulence in the phytopathogenic fungus Botryosphaeria dothidea. Viruses 2019, 11, 266. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhai, L.; Zhang, M.; Luo, G.; Wen, Y.; Cao, T.; Xia, H.; Zhang, J.; Liu, M. Molecular characterization of a novel victorivirus isolated from Botryosphaeria dothidea, the causal agent of longan leaf spot disease. Arch. Virol. 2022, 167, 2417–2422. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, M.; Zhu, H.; Zhong, J.; Liao, X.; Zhou, Q. Molecular characterization of a novel mitovirus from the plant-pathogenic fungus Botryosphaeria dothidea. Arch. Virol. 2021, 166, 633–637. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhai, L.; Xiao, F.; Guo, Y.; Fu, M.; Hong, N.; Wang, G. Characterization of a novel victorivirus isolated from the phytopathogenic fungus Botryosphaeria dothidea. Arch. Virol. 2019, 164, 1609–1617. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Zhou, T.; Guo, L.Y. Characterization of a novel strain of Botryosphaeria dothidea chrysovirus 1 from the apple white rot pathogen Botryosphaeria dothidea. Arch. Virol. 2017, 162, 2097–2102. [Google Scholar] [CrossRef]
- Zhai, L.; Hong, N.; Zhang, M.; Wang, G. Complete dsRNA sequence of a novel victorivirus isolated from the pear stem wart fungus Botryosphaeria dothidea. Arch. Virol. 2015, 160, 613–616. [Google Scholar] [CrossRef]
- Yang, M.; Xu, W.; Zhou, X.; Yang, Z.; Wang, Y.; Xiao, F.; Guo, Y.; Hong, N.; Wang, G. Discovery and characterization of a novel bipartite Botrexvirus from the phytopathogenic fungus Botryosphaeria dothidea. Front. Microbiol. 2021, 12, 696125. [Google Scholar] [CrossRef]
- Liu, W.; Hai, D.; Mu, F.; Yu, X.; Zhao, Y.; He, B.; Xie, J.; Jiang, D.; Liu, H. Molecular characterization of a novel fusarivirus infecting the plant-pathogenic fungus Botryosphaeria dothidea. Arch. Virol. 2020, 165, 1033–1037. [Google Scholar] [CrossRef]
- He, Y.; Zou, Q.; Li, S.; Zhu, H.; Hong, N.; Wang, G.; Wang, L. Molecular characterization of a new fusarivirus infecting Botryosphaeria dothidea, the causal agent of pear ring rot disease. Arch. Virol. 2022, 167, 1893–1897. [Google Scholar] [CrossRef]
- Zou, Q.; Gao, Y.; Wang, Q.; Yang, Y.; Wang, F.; Hong, N.; Wang, G.; Wang, L. The full-length genome sequence of a novel mitovirus from Botryosphaeria dothidea, the causal agent of pear ring rot disease. Arch. Virol. 2021, 166, 2881–2885. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, H.; Lu, X.; Wang, Y.; Zhou, Q. A novel mitovirus isolated from the phytopathogenic fungus Botryosphaeria dothidea. Arch. Virol. 2021, 166, 1507–1511. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, H.; Zhou, Q. A novel mycovirus isolated from the plant-pathogenic fungus Botryosphaeria dothidea. Arch. Virol. 2021, 166, 1267–1272. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Cao, J.; Xie, S.; Wang, Y.; Yin, X.; Guo, Y.; Xu, C.; Guo, L.; Wu, H.; Zhang, M. Molecular characterization of a novel ourmia-like virus from the phytopathogenic fungus Botryosphaeria dothidea. Arch. Virol. 2023, 168, 106. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhou, X.; Zhai, L.; Xiao, F.; Hong, N.; Wang, G. Molecular characterization of a novel mycovirus infecting the phytopathogenic fungus Botryosphaeria dothidea. Arch. Virol. 2020, 165, 1667–1670. [Google Scholar] [CrossRef] [PubMed]
- Lian, Z.; Das, S.; Luo, J.; Andika, I.B.; Sun, L. Complete genome sequence of a novel ourmia-like mycovirus infecting the phytopathogenic fungus Botryosphaeria dothidea. Arch. Virol. 2021, 166, 3461–3465. [Google Scholar] [CrossRef] [PubMed]
- Dong, K.; Xu, C.; Kotta-Loizou, I.; Jiang, J.; Lv, R.; Kong, L.; Li, S.; Hong, N.; Wang, G.; Coutts, R.H.A.; et al. Novel viroid-like RNAs naturally infect a filamentous fungus. Adv. Sci. 2023, 10, e2204308. [Google Scholar] [CrossRef]
- Nerva, L.; Turina, M.; Zanzotto, A.; Gardiman, M.; Gaiotti, F.; Gambino, G.; Chitarra, W. Isolation, molecular characterization and virome analysis of culturable wood fungal endophytes in esca symptomatic and asymptomatic grapevine plants. Environ. Microbiol. 2019, 21, 2886–2904. [Google Scholar] [CrossRef]
- Marais, A.; Faure, C.; Comont, G.; Candresse, T.; Stempien, E.; Corio-Costet, M.F. Characterization of the mycovirome of the phytopathogenic fungus, Neofusicoccum parvum. Viruses 2021, 13, 375. [Google Scholar] [CrossRef]
- Khan, H.A.; Sato, Y.; Kondo, H.; Jamal, A.; Bhatti, M.F.; Suzuki, N. A novel victorivirus from the phytopathogenic fungus Neofusicoccum parvum. Arch. Virol. 2022, 167, 923–929. [Google Scholar] [CrossRef]
- Khan, H.A.; Telengech, P.; Kondo, H.; Bhatti, M.F.; Suzuki, N. Mycovirus hunting revealed the presence of diverse viruses in a single isolate of the phytopathogenic fungus Diplodia seriata from Pakistan. Front. Cell. Infect. Microbiol. 2022, 12, 913619. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are non orthologous. Molec. Phylogen. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Campbell, C.L.; Madden, L.V. Introduction to Plant Disease Epidemiology; Wiley: New York, NY, USA, 1990; p. 560. [Google Scholar]
- Marais, A.; Faure, C.; Bergey, B.; Candresse, T. Viral double-stranded RNAs (dsRNAs) from plants: Alternative nucleic acid substrates for high-throughput sequencing. Meth. Molec. Biol. 2018, 1746, 45–53. [Google Scholar] [CrossRef]
- Marais, A.; Nivault, A.; Faure, C.; Theil, S.; Comont, G.; Candresse, T.; Corio-Costet, M.F. Determination of the complete genomic sequence of Neofusicoccum luteum mitovirus 1 (NLMV1), a novel mitovirus associated with a phytopathogenic Botryosphaeriaceae. Arch. Virol. 2017, 162, 2477–2480. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing large minimum evolution trees with profiles instead of a distance matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef]
- Marais, A.; Nivault, A.; Faure, C.; Comont, G.; Theil, S.; Candresse, T.; Corio-Costet, M.F. Molecular characterization of a novel fusarivirus infecting the plant-pathogenic fungus Neofusicoccum luteum. Arch. Virol. 2018, 163, 559–562. [Google Scholar] [CrossRef]
- Marais, A.; Faure, C.; Mustafayev, E.; Barone, M.; Alioto, D.; Candresse, T. Characterization by deep sequencing of Prunus virus T, a novel Tepovirus infecting Prunus species. Phytopathology 2015, 105, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Ding, S.W.; Zhang, Y.; Zhu, S. Identification of viruses and viroids by next-generation sequencing and homology-dependent and homology-independent algorithms. Annu. Rev. Phytopathol. 2015, 53, 425–444. [Google Scholar] [CrossRef] [PubMed]
- Roossinck, M.J.; Martin, D.P.; Roumagnac, P. Plant Virus Metagenomics: Advances in Virus Discovery. Phytopathology 2015, 105, 716–727. [Google Scholar] [CrossRef]
- Valverde, R.A.; Khalifa, M.E.; Okada, R.; Fukuhara, T.; Sabanadzovic, S.; ICTV Report Consortium. ICTV virus taxonomy profile. Endornaviridae. J. Gen. Virol. 2019, 100, 1204–1205. [Google Scholar] [CrossRef]
- Vainio, E.J.; Chiba, S.; Ghabrial, S.A.; Maiss, E.; Roossinck, M.; Sabanadzovic, S.; Suzuki, N.; Xie, J.; Nibert, M.; ICTV Report Consortium. ICTV virus Taxonomy Profile: Partitiviridae. J Gen Virol. 2018, 99, 17–18. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, J.; Zhang, H.; Qiu, D.; Guo, L. Two novel relative double-stranded RNA mycoviruses infecting Fusarium poae Strain SX63. Int. J. Mol. Sci. 2016, 17, 641. [Google Scholar] [CrossRef] [PubMed]
- De Wet, J.; Bihon, W.; Preisig, O.; Wingfield, B.D.; Wingfield, M.J. Characterization of a novel dsRNA element in the pine endophytic fungus Diplodia scrobiculata. Arch. Virol. 2011, 156, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.L.; Atkins, J.F.; Gesteland, R.F. Programmed ribosomal frameshifting: Much ado about knotting! Proc. Natl. Acad. Sci. USA 1999, 96, 14177–14179. [Google Scholar] [CrossRef] [PubMed]
- Bruenn, J.A. A closely related group of RNA-dependent RNA polymerases from double-stranded RNA viruses. Nucleic Acids Res. 1993, 21, 5667–5669. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ni, Y.; Liu, X.; Zhao, H.; Xiao, Y.; Xiao, X.; Li, S.; Liu, H. Divergent RNA viruses in Macrophomina phaseolina exhibit potential as virocontrol agents. Virus Evol. 2020, 7, veaa095. [Google Scholar] [CrossRef] [PubMed]
- Picarelli, M.A.S.C.; Forgia, M.; Rivas, E.B.; Nerva, L.; Chiapello, M.; Turina, M.; Colariccio, A. Extreme diversity of mycoviruses present in isolates of Rhizoctonia solani AG2-2 LP from Zoysia japonica From Brazil. Front. Cell Infect. Microbiol. 2019, 9, 244. [Google Scholar] [CrossRef] [PubMed]
- Slippers, B.; Crous, P.W.; Jami, F.; Groenewald, J.Z.; Wingfield, M.J. Diversity in Botryosphaeriales: Looking back, looking forward. Fungal Biol. 2017, 121, 307–321. [Google Scholar] [CrossRef]
- Arjona-Lopez, J.M.; Telengech, P.; Jamal, A.; Hisano, S.; Kondo, H.; Yelin, M.D.; Arjona-Girona, I.; Kanematsu, S.; Lopez-Herrera, C.J.; Suzuki, N. Novel, diverse RNA viruses from Mediterranean isolates of the phytopathogenic fungus, Rosellinia necatrix: Insights into evolutionary biology of fungal viruses. Environ. Microbiol. 2018, 20, 1464–1483. [Google Scholar] [CrossRef]
- Deng, F.; Xu, R.; Boland, G.J. Hypovirulence-associated double-stranded RNA from Sclerotinia homoeocarpa is conspecific with Ophiostoma novo-ulmi mitovirus 3a-Ld. Phytopathology 2003, 93, 1407–1414. [Google Scholar] [CrossRef]
- Vainio, E.J.; Pennanen, T.; Rajala, T.; Hantula, J. Occurrence of similar mycoviruses in pathogenic, saprotrophic and mycorrhizal fungi inhabiting the same forest stand. FEMS Microbiol. Ecol. 2017, 93, fix003. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Zhou, K.; Wu, M.; Zhang, J.; Yang, L.; Chen, W.; Li, G. Viral cross-class transmission results in disease of a phytopathogenic fungus. ISME J. 2022, 16, 2763–2774. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Zhang, L.; Li, G.; Jiang, D.; Ghabrial, S.A. Genome characterization of a debilitation-associated mitovirus infecting the phytopathogenic fungus Botrytis cinerea. Virology 2010, 406, 117–126. [Google Scholar] [CrossRef]
- Ghabrial, S.A. Origin, adaptation and evolutionary pathways of fungal viruses. Virus Genes 1998, 16, 119–131. [Google Scholar] [CrossRef]
- Myers, J.M.; James, T.Y. Mycoviruses. Curr. Biol. 2022, 32, R150–R155. [Google Scholar] [CrossRef]
- Wu, S.; Cheng, J.; Fu, Y.; Chen, T.; Jiang, D.; Ghabrial, S.A.; Xie, J. Virus-mediated suppression of host non-self recognition facilitates horizontal transmission of heterologous viruses. PLoS Pathog. 2017, 13, e1006234. [Google Scholar] [CrossRef]
- Xie, J.; Jiang, D. New insights into mycoviruses and exploration for the biological control of crop fungal diseases. Annu. Rev. Phytopath. 2014, 52, 45–68. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, B.; Fu, Y.; Xie, J.; Cheng, J.; Ghabrial, S.A.; Li, G.; Yi, X.; Jiang, D. Extracellular transmission of a DNA mycovirus and its use as a natural fungicide. Proc. Natl. Acad. Sci. USA 2013, 110, 1452–1457. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Xie, J.; Cheng, J.; Li, B.; Chen, T.; Fu, Y.; Li, G.; Wang, M.; Jin, H.; Wan, H.; et al. Fungal DNA virus infects a mycophagous insect and utilizes it as a transmission vector. Proc. Natl. Acad. Sci. USA 2016, 113, 12803–12808. [Google Scholar] [CrossRef] [PubMed]
- Hao, F.; Wu, M.; Li, G. Molecular characterization and geographic distribution of a mymonavirus in the population of Botrytis cinerea. Viruses 2018, 10, 432. [Google Scholar] [CrossRef] [PubMed]
- Kuroki, M.; Yaguchi, T.; Urayama, S.; Hagiwara, D. Experimental verification of strain-dependent relationship between mycovirus and its fungal host. iScience 2023, 26, 107337. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Species | Isolate | Country | Region | Host | Cultivar | Sampling Date | Symptoms | Sample Localization | AUC at 28 °C ** | Necrosis Length (mm) | Viruses |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Botryosphaeria | B. dothidea | CBS110302 * | Portugal | Montemor-o-Novo | V. vinifera | na | 1996 | na | na | na | na | 0 |
| Botryosphaeria | B. dothidea | LAT32 | France | New Aquitaine | V. vinifera | Cabernet franc | 2008 | S | N | na | na | DsEV1 |
| Botryosphaeria | B. dothidea | OGE14 | France | Champagne | V. vinifera | Pinot noir | 2008 | S | B | na | na | 0 |
| Diplodia | D. intermedia | BEI06 | France | Burgundy | V. vinifera | Chardonnay | 2008 | S | B | na | na | 0 |
| Diplodia | D. intermedia | BEI39 | France | Burgundy | V. vinifera | Chardonnay | 2008 | H | B | 819 | 7.3 | 0 |
| Diplodia | D. mutila | ARB07 | France | Jura | V. vinifera | Trousseau | 2008 | S | B | na | na | 0 |
| Diplodia | D. mutila | ARB44 | France | Jura | V. vinifera | Trousseau | 2008 | H | B | na | na | 0 |
| Diplodia | D. mutila | ARB45 | France | Jura | V. vinifera | Trousseau | 2008 | H | B | 245 | 2.75 | DmFV1 |
| Diplodia | D. mutila | BEI36 | France | Burgundy | V. vinifera | Chardonnay | 2008 | H | B | 330 | 3.67 | 0 |
| Diplodia | D. mutila | BRA08 | France | Champagne | V. vinifera | Pinot noir | 2008 | S | B | 1150 | 8.6 | DsPV1 DsEV1 |
| Diplodia | D. mutila | CBS112553 * | Portugal | Montemor-o-Novo | V. vinifera | na | 1997 | na | na | na | na | 0 |
| Diplodia | D. mutila | GRA09 | France | New Aquitaine | V. vinifera | Ugni blanc | 2008 | S | N | na | na | 0 |
| Diplodia | D. mutila | LAG01 | France | New Aquitaine | V. vinifera | Cabernet Sauvignon | 2008 | S | N | 928 | 13.3 | DsEV1 |
| Diplodia | D. mutila | LAG27 | France | New Aquitaine | V. vinifera | Cabernet Sauvignon | 2008 | S | N | na | na | 0 |
| Diplodia | D. mutila 1 | CBS43.182 * | Netherlands | Maarseveen | Fraxinus excelsior | na | 1982 | na | na | na | na | 0 |
| Diplodia | D. rosulata | CBS116470 * | Ethiopia | na | Prunus africana | na | 2001 | na | na | na | na | 0 |
| Diplodia | D. sapinea | CBS109725 * | South Africa | Habinsaran | Pinus patula | na | 2001 | na | na | na | na | NpEV1 |
| Diplodia | D. scrobiculata | CBS118110 * | USA | Wisconsin | Pinus banksiana | na | na | na | na | na | na | NpEV1 DsEV1 |
| Diplodia | D. seriata | ARB01 | France | Jura | V. vinifera | Trousseau | 2008 | S | B | na | na | 0 |
| Diplodia | D. seriata | ARB18 | France | Jura | V. vinifera | Trousseau | 2008 | S | N | na | na | 0 |
| Diplodia | D. seriata | BEI03 | France | Burgundy | V. vinifera | Chardonnay | 2008 | S | N | na | na | 0 |
| Diplodia | D. seriata | BEI25 | France | Burgundy | V. vinifera | Chardonnay | 2008 | S | N | na | na | 0 |
| Diplodia | D. seriata | BoF00-14 | France | Champagne | V. vinifera | Pinot Meunier | 2000 | S | na | na | na | NpEV1 |
| Diplodia | D. seriata | BoF00-5 | France | New Aquitaine | V. vinifera | Cabernet Sauvignon | 2000 | S | N | na | na | 0 |
| Diplodia | D. seriata | BoF98-1 | France | Languedoc Roussillon | V. vinifera | Syrah | 1998 | S | na | 945 | 9.82 | DsBMV1 DsNV1 |
| Diplodia | D. seriata | BoF99-7 | France | Rhône Valley | V. vinifera | Clairette | 1999 | S | na | na | na | NpEV1 |
| Diplodia | D. seriata | BoF99-8 | France | Languedoc Roussillon | V. vinifera | Syrah | 1999 | S | na | na | na | DsEV1 |
| Diplodia | D. seriata | BRA16 | France | Champagne | V. vinifera | Pinot noir | 2008 | S | B | 1306 | 6.8 | 0 |
| Diplodia | D. seriata | CBS112555 * | Portugal | Montemor-o-Novo | V. vinifera | na | 1997 | na | na | na | na | 0 |
| Diplodia | D. seriata | IRA 21 | France | Burgundy | V. vinifera | Pinot noir | 2008 | S | B | na | na | DsEV1 |
| Diplodia | D. seriata | IRA33 | France | Burgundy | V. vinifera | Pinot noir | 2008 | S | B | na | na | DsEV1 |
| Diplodia | D. seriata | LAG13 | France | New Aquitaine | V. vinifera | Cabernet Sauvignon | 2008 | S | N | na | na | 0 |
| Diplodia | D. seriata | LAT16 | France | New Aquitaine | V. vinifera | Cabernet franc | 2008 | S | N | 1182 | 5.03 | 0 |
| Diplodia | D. seriata | LAT28 | France | New Aquitaine | V. vinifera | Cabernet franc | 2008 | S | B | 1292 | 5.63 | 0 |
| Diplodia | D. seriata | MOT02 | France | Burgundy | V. vinifera | Chardonnay | 2008 | S | N | na | na | 0 |
| Diplodia | D. seriata | PER01 | France | Champagne | V. vinifera | Chardonnay | 2008 | S | B | na | na | DsEV1 |
| Diplodia | D. seriata | PLU03 | France | Champagne | V. vinifera | Chardonnay | 2008 | S | N | na | na | 0 |
| Diplodia | D. seriata | ROM14 | France | Champagne | V. vinifera | Chardonnay | 2008 | S | B | na | na | DsEV1 |
| Diplodia | D. seriata | ROU03 | France | Alsace | V. vinifera | Gewurztraminer | 2008 | S | B | na | na | 0 |
| Diplodia | D. seriata | TUR16 | France | Alsace | V. vinifera | Auxerrois | 2008 | S | N | na | na | 0 |
| Diplodia | D. seriata | VIE51 | France | Champagne | V. vinifera | Chardonnay | 2008 | H | B | na | na | 0 |
| Lasiodiplodia | L. pseudotheobromae | CBS116459 * | Costa Rica | San Carlos | Gmelina arborea | na | na | na | na | na | na | 0 |
| Lasiodiplodia | L. pseudotheobromae | CBS116460 * | Costa Rica | San Carlos | Acacia mangium | na | na | na | na | na | na | 0 |
| Lasiodiplodia | L. viticola | CBS128313 * | USA | Arkansas | V. vinifera | Vignoles | na | S | na | na | na | 0 |
| Lasiodiplodia | L. viticola | LAG05 | France | New Aquitaine | V. vinifera | Cabernet Sauvignon | 2008 | S | H-p | 1336 | 23.30 | DsPV1 |
| Lasiodiplodia | L. viticola | LAG78 | France | New Aquitaine | V. vinifera | Cabernet-Sauvignon | 2008 | S | H-p | na | na | 0 |
| Neofusicoccum | N. luteum | CAP37 | Portugal | na | V. vinifera | na | na | na | na | na | na | NlMV1 NlFV1 |
| Neofusicoccum | N. luteum | CBS110299 * | Portugal | Oeiras | V. vinifera | na | 1996 | na | na | na | na | NlMV1 NlFV1 |
| Neofusicoccum | N. parvum | ALI03 | France | Languedoc Roussillon | V. vinifera | Sauvignon | 2008 | S | N | na | na | AtNSRV1 NpVV1 |
| Neofusicoccum | N. parvum | ALI30 | France | Languedoc Roussillon | V. vinifera | Sauvignon | 2008 | H | N | na | na | NpVV1 NpMV3 DsEV1 |
| Neofusicoccum | N. parvum | AUD25 | France | Languedoc Roussillon | V. vinifera | na | 2008 | S | H-p | 1093 | 13.62 | 0 |
| Neofusicoccum | N. parvum | AUD31 | France | Languedoc Roussillon | V. vinifera | na | 2008 | S | N | na | na | 0 |
| Neofusicoccum | N. parvum | BdF00-14 | France | New Aquitaine | V. vinifera | Merlot | 2000 | S | na | na | na | 0 |
| Neofusicoccum | N. parvum | BdF00-21 | France | Champagne | V. vinifera | na | 2000 | S | N | na | na | 0 |
| Neofusicoccum | N. parvum | BdF00-3 | France | Languedoc Roussillon | V. vinifera | Syrah | 2000 | S | na | na | na | 0 |
| Neofusicoccum | N. parvum | BdF00-8 | France | New Aquitaine | V. vinifera | Merlot | 2000 | S | N | na | na | 0 |
| Neofusicoccum | N. parvum | CBS110301 * | Portugal | na | V. vinifera | na | 1996 | - | na | na | na | 0 |
| Neofusicoccum | N. parvum | COLB | France | Burgundy | V. vinifera | Chardonnay | 2009 | S | na | na | na | NpNV3 NpEV1 NpMV2 NpMV3 NpVV1 NpVV2 |
| Neofusicoccum | N. parvum | COU02 | France | New Aquitaine | V. vinifera | Cabernet Sauvignon | 2008 | S | N | na | na | NpMV3 NpEV1 |
| Neofusicoccum | N. parvum | PER04 | France | Champagne | V. vinifera | Chardonnay | 2008 | S | B | na | na | AtNSRV1 |
| Neofusicoccum | N. parvum | PER20 | France | Champagne | V. vinifera | Chardonnay | 2008 | S | N | 1717 | 19.22 | AtNSRV1 NpVV1 DmFV1 NpNV3 |
| Neofusicoccum | N. parvum | SAI07 | France | Burgundy | V. vinifera | Ugni blanc | 2008 | S | N | na | na | 0 |
| Neofusicoccum | N. parvum | VIE35 | France | Champagne | V. vinifera | Chardonnay | 2008 | S | H-p | 1665 | 33.70 | 0 |
| Neofusicoccum | N. ribis | CBS114472 * | Hawaii | na | Leucadron Safari Sunset | na | 1998 | na | na | na | na | 0 |
| Neofusicoccum | N. ribis | CBS115475 * | USA | New York | Ribes sp. | na | 1998 | na | na | na | na | 0 |
| Spencermartinsia | S. viticola | CBS117009 * | Spain | Catalonia | V. vinifera | Garnatxa negra | 2004 | na | na | na | na | 0 |
| Spencermartinsia | S. viticola | CBS121000 * | USA | California | V. vinifera | Cabernet Sauvignon | 2008 | na | na | 666 | 8 | 0 |
| Spencermartinsia | S. viticola | GAR09 | France | Languedoc-Roussillon | V. vinifera | Sauvignon | 2008 | S | B | 276 | 2.39 | 0 |
| Spencermartinsia | S. viticola | GAR47 | France | Languedoc Roussillon | V. vinifera | Sauvignon | 2008 | H | B | na | na | 0 |
| Species | Isolate | Virus | Total Reads | Mapped Reads (% of Total Reads) | Average Coverage | Contig Length (nt) | Predicted Protein Encoded 1 | Accession Number |
|---|---|---|---|---|---|---|---|---|
| D. mutila | ARB45 | DmFV1 | 446,665 | 347,375 (77.8%) | 4905 | 8725 | HP, RdRp | ON236579 2 |
| D. mutila | BRA08 | DsPV1 RNA1 | 1,499,218 | 82,833 (5.5%) | 12,068 | 1529 | RdRp | ON236584 3 |
| DsPV1 RNA2 | 50,293 (3.4%) | 6759 | 1456 | CP | ON236585 3 | |||
| DsEV1 | 875,766 (58.4%) | 52,996 | 9848 | Polyprotein | ON236581 3 | |||
| D. mutila | LAG01 | DsEV1 | 1,313,885 | 1,103,013 (84%) | 13,199 | 10,127 | Polyprotein | ON236580 2 |
| L. viticola | LAG05 | DsPV1 RNA1 | 958,446 | 324,630 (33.9%) | 52,996 | 1338 | RdRp | ON236582 3 |
| DsPV1 RNA2 | 109,971 (11.5%) | 15,666 | 1449 | CP | ON236583 3 | |||
| N. parvum | PER20 | NpVV1 | 1,302,602 | 845 (0.1%) | 29 | 5188 | CP, RdRp | ON236575 3 |
| DmFV1 | 112,024 (8.6%) | 2909 | 8185 | HP, RdRp | ON236578 3 | |||
| NpNV3 | 1258 (0.1%) | 130 | 2071 | RdRp | ON236576 3 | |||
| AtNsRV1 | 254,239 (19.5%) | 6356 | 8921 | RdRp | ON236577 3 | |||
| D. seriata | BoF981 | DsMBV1 | 1,314,206 | 94,816 (7.2%) | 2064 | 10,339 | RdRp | ON236586 3 |
| DsNV1 | 9504 (0.7%) | 616 | 3652 | RdRp | ON236587 3 |
| Virus | Number of Positive Isolates | Range of Pairwise Nucleotide Divergence |
|---|---|---|
| AtNSRV1 | 3 | 1.5–10.2% |
| NpVV1 | 4 | 0–9.8% |
| NpMV3 | 3 | 0–4.2% |
| DsPV1 | 2 | 0.7–4.7% |
| NpEV1 | 6 | 0–11.5% |
| DsEV1 | 10 | 4.3–12.5% |
| NpNV3 | 2 | 0–2.6% |
| NlFV1 | 2 | 0 |
| NlMV1 | 2 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Comont, G.; Faure, C.; Candresse, T.; Laurens, M.; Valière, S.; Lluch, J.; Lefebvre, M.; Gambier, S.; Jolivet, J.; Corio-Costet, M.-F.; et al. Characterization of the RNA Mycovirome Associated with Grapevine Fungal Pathogens: Analysis of Mycovirus Distribution and Their Genetic Variability within a Collection of Botryosphaeriaceae Isolates. Viruses 2024, 16, 392. https://doi.org/10.3390/v16030392
Comont G, Faure C, Candresse T, Laurens M, Valière S, Lluch J, Lefebvre M, Gambier S, Jolivet J, Corio-Costet M-F, et al. Characterization of the RNA Mycovirome Associated with Grapevine Fungal Pathogens: Analysis of Mycovirus Distribution and Their Genetic Variability within a Collection of Botryosphaeriaceae Isolates. Viruses. 2024; 16(3):392. https://doi.org/10.3390/v16030392
Chicago/Turabian StyleComont, Gwenaëlle, Chantal Faure, Thierry Candresse, Marie Laurens, Sophie Valière, Jérôme Lluch, Marie Lefebvre, Sébastien Gambier, Jérôme Jolivet, Marie-France Corio-Costet, and et al. 2024. "Characterization of the RNA Mycovirome Associated with Grapevine Fungal Pathogens: Analysis of Mycovirus Distribution and Their Genetic Variability within a Collection of Botryosphaeriaceae Isolates" Viruses 16, no. 3: 392. https://doi.org/10.3390/v16030392