
Rosuvastatin as a Supplemental Treatment for the Clinical Symptoms of Nephropathia Epidemica: A Pilot Clinical Study
 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. The Severity of the Disease
2.3. Statin Treatment
2.4. Controls
2.5. Inclusion Criteria
2.6. Exclusion Criteria
2.7. Ethics Statement
2.8. Othohantavirus ELISA
2.9. Serum Lipid Analysis
2.10. RT-PCR Detection and Sequencing of PUUV Transcripts
2.11. Multiplex Analysis
2.12. Statistical Analysis
3. Results
3.1. PUUV Genetic Variations in NE Patients
3.2. Patients
3.3. Analysis of Clinical Laboratory Data in NE Patients and Controls
3.4. Analysis of Rosuvastatin Treatment on Clinical Signs and Symptoms in NE Patients
3.5. The Effect of Rosuvastatin on Serum Lipid Levels
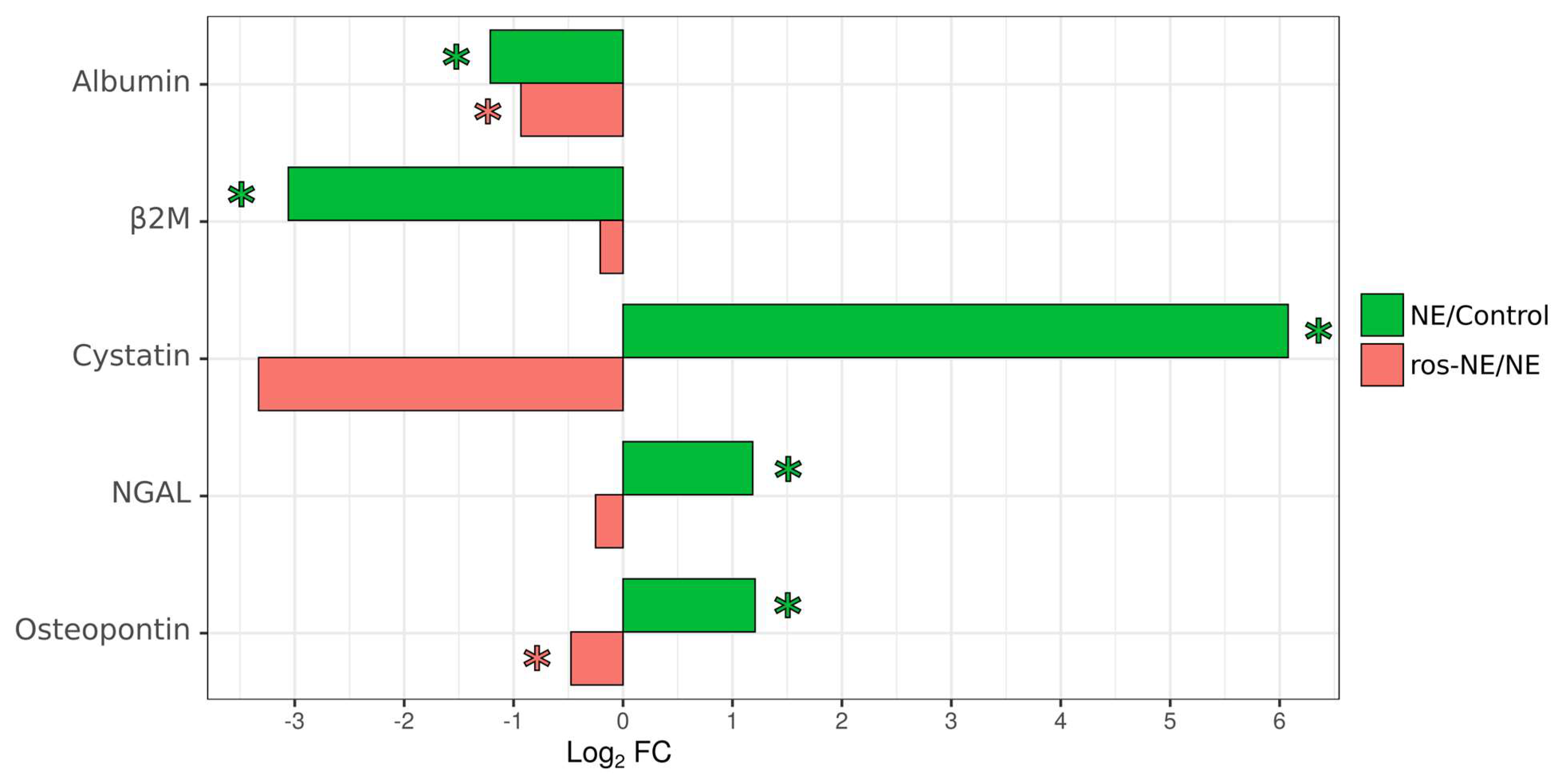
3.6. Effect of Rosuvastatin on Urine Kidney Toxicity Markers in NE Patients
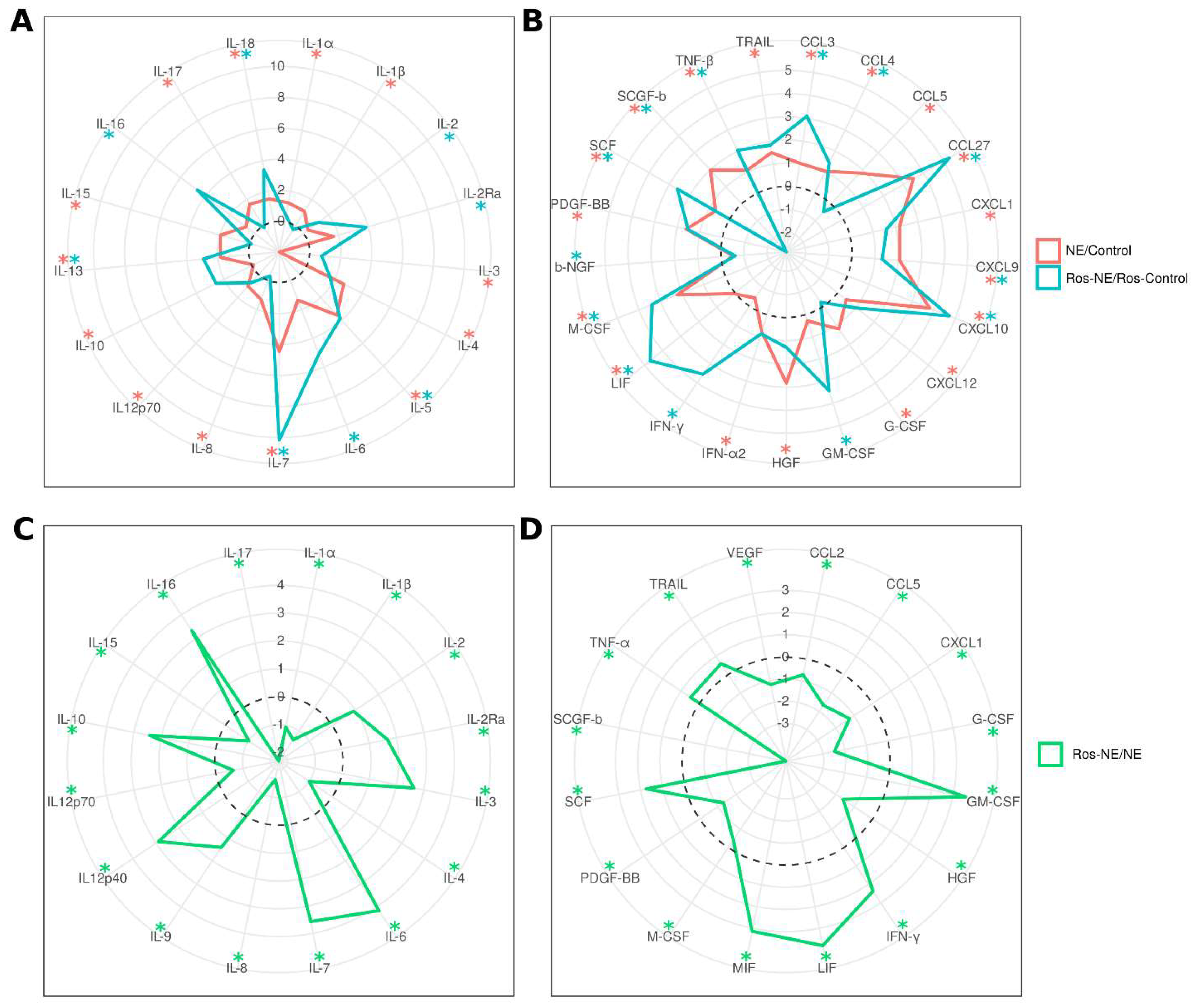
3.7. Effect of Rosuvastatin on Serum Cytokines in NE Patients
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Latus, J.; Kitterer, D.; Segerer, S.; Artunc, F.; Alscher, M.D.; Braun, N. Determination of procalcitonin levels in patients with nephropathia epidemica-a useful tool or an unnecessary diagnostic procedure? Kidney Blood Press. Res. 2015, 40, 22–30. [Google Scholar] [CrossRef]
- Stock, I. Hantavirus infections. Med. Monatsschrift Fur Pharm. 2008, 31, 127–136, quiz 137. [Google Scholar]
- Davidyuk, Y.N.; Kabwe, E.; Shakirova, V.G.; Martynova, E.V.; Ismagilova, R.K.; Khaertynova, I.M.; Khaiboullina, S.F.; Rizvanov, A.A.; Morzunov, S.P. Characterization of the Puumala orthohantavirus strains in the northwestern region of the Republic of Tatarstan in relation to the clinical manifestations in hemorrhagic fever with renal syndrome patients. Front. Pharmacol. 2019, 10, 970. [Google Scholar] [CrossRef]
- Temonen, M.; Vapalahti, O.; Holthöfer, H.; Brummer-Korvenkontio, M.; Vaheri, A.; Lankinen, H. Susceptibility of human cells to Puumala virus infection. J. Gen. Virol. 1993, 74, 515–518. [Google Scholar] [CrossRef]
- Kim, J.-H.; Park, H.-H.; Choi, I.-H.; Kim, Y.-O.; Cho, K.-H. Severely modified lipoprotein properties without a change in cholesteryl ester transfer protein activity in patients with acute renal failure secondary to Hantaan virus infection. BMB Rep. 2010, 43, 535–540. [Google Scholar] [CrossRef]
- Martynova, E.V.; Valiullina, A.H.; Gusev, O.A.; Davidyuk, Y.N.; Garanina, E.E.; Shakirova, V.G.; Khaertynova, I.; Anokhin, V.A.; Rizvanov, A.A.; Khaiboullina, S.F. High triglycerides are associated with low thrombocyte counts and high VEGF in nephropathia epidemica. J. Immunol. Res. 2016, 2016, 8528270. [Google Scholar] [CrossRef]
- McCaughey, C.; Shi, X.; Elliott, R.M.; Wyatt, D.E.; O’Neill, H.J.; Coyle, P.V. Low pH-induced cytopathic effect—A survey of seven hantavirus strains. J. Virol. Methods 1999, 81, 193–197. [Google Scholar] [CrossRef]
- Cifuentes-Muñoz, N.; Darlix, J.-L.; Tischler, N.D. Development of a lentiviral vector system to study the role of the Andes virus glycoproteins. Virus Res. 2010, 153, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.-F.; Flint, M.; Lin, J.-M.S.; Spiropoulou, C.F. Endocytic pathways used by Andes virus to enter primary human lung endothelial cells. PLoS ONE 2016, 11, e0164768. [Google Scholar] [CrossRef] [PubMed]
- Kai, S.; Elina, I. Functional rafts in cell membranes. Nature 1997, 387, 569–572. [Google Scholar]
- Silvius, J.R. Role of cholesterol in lipid raft formation: Lessons from lipid model systems. Biochim. Et Biophys. Acta BBA-Biomembr. 2003, 1610, 174–183. [Google Scholar] [CrossRef]
- Petersen, J.; Drake, M.J.; Bruce, E.A.; Riblett, A.M.; Didigu, C.A.; Wilen, C.B.; Malani, N.; Male, F.; Lee, F.-H.; Bushman, F.D. The major cellular sterol regulatory pathway is required for Andes virus infection. PLoS Pathog. 2014, 10, e1003911. [Google Scholar] [CrossRef] [PubMed]
- Kleinfelter, L.M.; Jangra, R.K.; Jae, L.T.; Herbert, A.S.; Mittler, E.; Stiles, K.M.; Wirchnianski, A.S.; Kielian, M.; Brummelkamp, T.R.; Dye, J.M. Haploid genetic screen reveals a profound and direct dependence on cholesterol for hantavirus membrane fusion. MBio 2015, 6, e00801–e00815. [Google Scholar] [CrossRef]
- Baigildina, A.A.; Khaiboullina, S.F.; Martynova, E.V.; Anokhin, V.A.; Lombardi, V.C.; Rizvanov, A.A. Inflammatory cytokines kinetics define the severity and phase of nephropathia epidemica. Biomark. Med. 2015, 9, 99–107. [Google Scholar] [CrossRef]
- Garanina, E.; Martynova, E.; Davidyuk, Y.; Kabwe, E.; Ivanov, K.; Titova, A.; Markelova, M.; Zhuravleva, M.; Cherepnev, G.; Shakirova, V.G. Cytokine storm combined with humoral immune response defect in fatal hemorrhagic fever with renal syndrome case, Tatarstan, Russia. Viruses 2019, 11, 601. [Google Scholar] [CrossRef] [PubMed]
- Outinen, T.K.; Mäkelä, S.M.; Ala-Houhala, I.O.; Huhtala, H.S.; Hurme, M.; Paakkala, A.S.; Pörsti, I.H.; Syrjänen, J.T.; Mustonen, J.T. The severity of Puumala hantavirus induced nephropathia epidemica can be better evaluated using plasma interleukin-6 than C-reactive protein determinations. BMC Infect. Dis. 2010, 10, 132. [Google Scholar] [CrossRef]
- Maes, P.; Clement, J.; Groeneveld, P.H.; Colson, P.; Huizinga, T.W.; Ranst, M.V. Tumor necrosis factor-α genetic predisposing factors can influence clinical severity in nephropathia epidemica. Viral Immunol. 2006, 19, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Grunfeld, C.; Dinarello, C.A.; Feingold, K.R. Tumor necrosis factor-α, interleukin-1, and interferon alpha stimulate triglyceride synthesis in HepG2 cells. Metabolism 1991, 40, 894–898. [Google Scholar] [CrossRef]
- FEINGOLD, K.R.; SOUED, M.; Serio, M.K.; Moser, A.H.; Dinarello, C.A.; Grunfeld, C. Multiple cytokines stimulate hepatic lipid synthesis in vivo. Endocrinology 1989, 125, 267–274. [Google Scholar] [CrossRef]
- Netea, M.G.; Kullberg, B.J.; Van der Meer, J.W. Circulating cytokines as mediators of fever. Clin. Infect. Dis. 2000, 31, S178–S184. [Google Scholar] [CrossRef]
- Lim, J.; Yang, E.J.; Chang, J.H. Upregulation of TNF-α by Triglycerides is Mediated by MEK1 Activation in Jurkat T Cells. Biomed. Sci. Lett. 2018, 24, 213–220. [Google Scholar] [CrossRef]
- Ma, P.; Gil, G.; Südhof, T.C.; Bilheimer, D.W.; Goldstein, J.L.; Brown, M.S. Mevinolin, an inhibitor of cholesterol synthesis, induces mRNA for low density lipoprotein receptor in livers of hamsters and rabbits. Proc. Natl. Acad. Sci. USA 1986, 83, 8370–8374. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.-Y.; Li, H.; Tang, J.-J.; Wang, J.; Luo, J.; Liu, B.; Wang, J.-K.; Shi, X.-J.; Cui, H.-W.; Tang, J. Discovery of a potent HMG-CoA reductase degrader that eliminates statin-induced reductase accumulation and lowers cholesterol. Nat. Commun. 2018, 9, 5138. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, T.; Giwa, A.O. Rosuvastatin. In StatPearls [Internet]; StatPearls Publishing: London, UK, 2022. [Google Scholar]
- Apotex. Rosuvastatin Calcium Tablets. Available online: www.apotex.com/products/us/downloads/pre/rosu_imtb_ins.pdf (accessed on 22 November 2020).
- Yushchuk, N. Infectious Diseases; National Leadership: Moscow, Russia, 2020; Volume 1, p. 1230. [Google Scholar]
- Martynova, E.; Davidyuk, Y.; Kabwe, E.; Garanina, E.E.; Shakirova, V.; Pavelkina, V.; Uskova, Y.; Stott, R.J.; Foster, T.L.; Markelova, M. Cytokine, Chemokine, and Metalloprotease Activation in the Serum of Patients with Nephropathia Epidemica from the Republic of Tatarstan and the Republic of Mordovia, Russia. Pathogens 2021, 10, 527. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- R-Project. The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 22 November 2020).
- Davidyuk, Y.; Shamsutdinov, A.; Kabwe, E.; Ismagilova, R.; Martynova, E.; Belyaev, A.; Shuralev, E.; Trifonov, V.; Savitskaya, T.; Isaeva, G. Prevalence of the Puumala orthohantavirus Strains in the Pre-Kama Area of the Republic of Tatarstan, Russia. Pathogens 2020, 9, 540. [Google Scholar] [CrossRef]
- Davidyuk, Y.N.; Kabwe, E.; Shamsutdinov, A.F.; Knyazeva, A.V.; Martynova, E.V.; Ismagilova, R.K.; Trifonov, V.A.; Savitskaya, T.A.; Isaeva, G.S.; Urbanowicz, R.A. The Distribution of Puumala orthohantavirus Genome Variants Correlates with the Regional Landscapes in the Trans-Kama Area of the Republic of Tatarstan. Pathogens 2021, 10, 1169. [Google Scholar] [CrossRef]
- Shakirova, V.; Khaertynova, I.; Markelova, M.; Tarlinton, R.; Behnke, J.; Martynova, E.; Garanina, E.; Rizvanov, A.; Khaiboullina, S. Serum Cytokine Alterations Associated with Age of Patients with Nephropathia Epidemica. BioMed Res. Int. 2022, 2022, 4685288. [Google Scholar] [CrossRef]
- Kata, D.; Földesi, I.; Feher, L.; Hackler Jr, L.; Puskas, L.; Gulya, K. Rosuvastatin enhances anti-inflammatory and inhibits pro-inflammatory functions in cultured microglial cells. Neuroscience 2016, 314, 47–63. [Google Scholar] [CrossRef]
- Khurana, S.; Gupta, S.; Bhalla, H.L.; Nandwani, S.; Gupta, V. Comparison of anti-inflammatory effect of atorvastatin with rosuvastatin in patients of acute coronary syndrome. J. Pharmacol. Pharmacother. 2015, 6, 130–135. [Google Scholar] [CrossRef]
- R McGuire, T.; C Kalil, A.; P Dobesh, P.; G Klepser, D.; M Olsen, K. Anti-inflammatory effects of rosuvastatin in healthy subjects: A prospective longitudinal study. Curr. Pharm. Des. 2014, 20, 1156–1160. [Google Scholar] [CrossRef] [PubMed]
- Qasim, S.; Alamgeer; Kalsoom, S.; Shahzad, M.; Bukhari, I.A.; Vohra, F.; Afzal, S. Rosuvastatin Attenuates Rheumatoid Arthritis-Associated Manifestations via Modulation of the Pro-and Anti-inflammatory Cytokine Network: A Combination of In Vitro and In Vivo Studies. ACS Omega 2021, 6, 2074–2084. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Overview of the IL-1 family in innate inflammation and acquired immunity. Immunol. Rev. 2018, 281, 8–27. [Google Scholar] [CrossRef]
- Eigenbrod, T.; Park, J.-H.; Harder, J.r.; Iwakura, Y.; Núñez, G. Cutting edge: Critical role for mesothelial cells in necrosis-induced inflammation through the recognition of IL-1α released from dying cells. J. Immunol. 2008, 181, 8194–8198. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, Y.; Nishimagi, E.; Tochimoto, A.; Kawamoto, M.; Katsumata, Y.; Soejima, M.; Kanno, T.; Kamatani, N.; Hara, M. Intracellular IL-1α-binding proteins contribute to biological functions of endogenous IL-1α in systemic sclerosis fibroblasts. Proc. Natl. Acad. Sci. USA 2006, 103, 14501–14506. [Google Scholar] [CrossRef] [PubMed]
- Cohen, I.; Rider, P.; Carmi, Y.; Braiman, A.; Dotan, S.; White, M.R.; Voronov, E.; Martin, M.U.; Dinarello, C.A.; Apte, R.N. Differential release of chromatin-bound IL-1α discriminates between necrotic and apoptotic cell death by the ability to induce sterile inflammation. Proc. Natl. Acad. Sci. USA 2010, 107, 2574–2579. [Google Scholar] [CrossRef] [PubMed]
- Mukaida, N. Pathophysiological roles of interleukin-8/CXCL8 in pulmonary diseases. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2003, 284, L566–L577. [Google Scholar] [CrossRef] [PubMed]
- Martín-Sánchez, F.; Diamond, C.; Zeitler, M.; Gomez, A.; Baroja-Mazo, A.; Bagnall, J.; Spiller, D.; White, M.; Daniels, M.; Mortellaro, A. Inflammasome-dependent IL-1β release depends upon membrane permeabilisation. Cell Death Differ. 2016, 23, 1219–1231. [Google Scholar] [CrossRef]
- Gleeson, T.A.; Nordling, E.; Kaiser, C.; Lawrence, C.B.; Brough, D.; Green, J.P.; Allan, S.M. Looking into the IL-1 of the storm: Are inflammasomes the link between immunothrombosis and hyperinflammation in cytokine storm syndromes? Discov. Immunol. 2022, 1, kyac005. [Google Scholar] [CrossRef]
- Kolb, M.; Margetts, P.J.; Anthony, D.C.; Pitossi, F.; Gauldie, J. Transient expression of IL-1β induces acute lung injury and chronic repair leading to pulmonary fibrosis. J. Clin. Investig. 2001, 107, 1529–1536. [Google Scholar] [CrossRef]
- Potere, N.; Del Buono, M.G.; Caricchio, R.; Cremer, P.C.; Vecchié, A.; Porreca, E.; Dalla Gasperina, D.; Dentali, F.; Abbate, A.; Bonaventura, A. Interleukin-1 and the NLRP3 inflammasome in COVID-19: Pathogenetic and therapeutic implications. EBioMedicine 2022, 85, 104299. [Google Scholar] [CrossRef]
- Cheng, R.; Wu, Z.; Li, M.; Shao, M.; Hu, T. Interleukin-1β is a potential therapeutic target for periodontitis: A narrative review. Int. J. Oral Sci. 2020, 12, 2. [Google Scholar] [CrossRef]
- Dinarello, C.A. Interleukin-1 in the pathogenesis and treatment of inflammatory diseases. Blood J. Am. Soc. Hematol. 2011, 117, 3720–3732. [Google Scholar] [CrossRef]
- Park, H.; Li, Z.; Yang, X.O.; Chang, S.H.; Nurieva, R.; Wang, Y.-H.; Wang, Y.; Hood, L.; Zhu, Z.; Tian, Q. A distinct lineage of CD4 T cells regulates tissue inflammation by producing interleukin 17. Nat. Immunol. 2005, 6, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- McGinley, A.M.; Sutton, C.E.; Edwards, S.C.; Leane, C.M.; DeCourcey, J.; Teijeiro, A.; Hamilton, J.A.; Boon, L.; Djouder, N.; Mills, K.H. Interleukin-17A serves a priming role in autoimmunity by recruiting IL-1β-producing myeloid cells that promote pathogenic T cells. Immunity 2020, 52, 342–356.e6. [Google Scholar] [CrossRef] [PubMed]
- Di Paolo, N.C.; Shayakhmetov, D.M. Interleukin 1α and the inflammatory process. Nat. Immunol. 2016, 17, 906–913. [Google Scholar] [CrossRef] [PubMed]
- Weber, A.; Wasiliew, P.; Kracht, M. Interleukin-1 (IL-1) pathway. Sci. Signal. 2010, 3, cm1. [Google Scholar] [CrossRef]
- Bernhard, S.; Hug, S.; Stratmann, A.E.P.; Erber, M.; Vidoni, L.; Knapp, C.L.; Thomaß, B.D.; Fauler, M.; Nilsson, B.; Nilsson Ekdahl, K. Interleukin 8 elicits rapid physiological changes in neutrophils that are altered by inflammatory conditions. J. Innate Immun. 2021, 13, 225–241. [Google Scholar] [CrossRef] [PubMed]
- Leonovich, O.; Dzagurova, T. Effects of Cholesterol on Internalization Puumala and Dobrava Hantavirus; FEBS: Milan, Italy, 2021; pp. 316–317. [Google Scholar]
- Venugopal, S.K.; Anoruo, M.; Jialal, I. Biochemistry, Low Density Lipoprotein; StatPearls Publishing: Treasure Island, FL, USA, 2018. [Google Scholar]
- Duewell, P.; Kono, H.; Rayner, K.J.; Sirois, C.M.; Vladimer, G.; Bauernfeind, F.G.; Abela, G.S.; Franchi, L.; Nunez, G.; Schnurr, M. NLRP3 inflammasomes are required for atherogenesis and activated by cholesterol crystals. Nature 2010, 464, 1357–1361. [Google Scholar] [CrossRef]
- Latus, J.; Schwab, M.; Tacconelli, E.; Pieper, F.-M.; Wegener, D.; Dippon, J.; Müller, S.; Zakim, D.; Segerer, S.; Kitterer, D. Clinical course and long-term outcome of hantavirus-associated nephropathia epidemica, Germany. Emerg. Infect. Dis. 2015, 21, 76. [Google Scholar] [CrossRef]
- Mironov, A.A.; Beznoussenko, G.V. Opinion: On the Way towards the New Paradigm of Atherosclerosis. Int. J. Mol. Sci. 2022, 23, 2152. [Google Scholar] [CrossRef]
- Mäkelä, S.; Ala-Houhala, I.; Mustonen, J.; Koivisto, A.-M.; Kouri, T.; Turjanmaa, V.; Vapalahti, O.; Vaheri, A.; Pasternack, A. Renal function and blood pressure five years after Puumala virus-induced nephropathy. Kidney Int. 2000, 58, 1711–1718. [Google Scholar] [CrossRef]
- Martynova, E.V.; Maksudova, A.N.; Shakirova, V.G.; Abdulkhakov, S.R.; Khaertynova, I.M.; Anokhin, V.A.; Ivanova, V.V.; Abiola, I.M.; Garanina, E.E.; Tazetdinova, L.G. Urinary clusterin is upregulated in nephropathia epidemica. Dis. Markers 2018, 2018, 8658507. [Google Scholar] [CrossRef]
- Koehler, F.C.; Di Cristanziano, V.; Späth, M.R.; Hoyer-Allo, K.J.R.; Wanken, M.; Müller, R.-U.; Burst, V. The kidney in hantavirus infection—Epidemiology, virology, pathophysiology, clinical presentation, diagnosis and management. Clin. Kidney J. 2022, 15, 1231–1252. [Google Scholar] [CrossRef] [PubMed]
- Bolisetty, S.; Agarwal, A. Urine albumin as a biomarker in acute kidney injury. Am. J. Physiol.-Ren. Physiol. 2011, 300, F626–F627. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Jin, H.; Holder, D.; Ozer, J.S.; Villarreal, S.; Shughrue, P.; Shi, S.; Figueroa, D.J.; Clouse, H.; Su, M. Urinary biomarkers trefoil factor 3 and albumin enable early detection of kidney tubular injury. Nat. Biotechnol. 2010, 28, 470–477. [Google Scholar] [CrossRef]
- Outinen, T.K.; Mäkelä, S.; Clement, J.; Paakkala, A.; Pörsti, I.; Mustonen, J. Community acquired severe acute kidney injury caused by hantavirus-induced hemorrhagic fever with renal syndrome has a favorable outcome. Nephron 2015, 130, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Lupuşoru, G.; Lupuşoru, M.; Ailincăi, I.; Bernea, L.; Berechet, A.; Spătaru, R.; Ismail, G. Hanta hemorrhagic fever with renal syndrome: A pathology in whose diagnosis kidney biopsy plays a major role. Exp. Ther. Med. 2021, 22, 984. [Google Scholar] [CrossRef] [PubMed]
- Owens, E.; Tan, K.-S.; Ellis, R.; Del Vecchio, S.; Humphries, T.; Lennan, E.; Vesey, D.; Healy, H.; Hoy, W.; Gobe, G. Development of a biomarker panel to distinguish risk of progressive chronic kidney disease. Biomedicines 2020, 8, 606. [Google Scholar] [CrossRef] [PubMed]
- Madsen, K.M.; Zhang, L.; Shamat, A.A.; Siegfried, S.; Cha, J.-H. Ultrastructural localization of osteopontin in the kidney: Induction by lipopolysaccharide. J. Am. Soc. Nephrol. 1997, 8, 1043–1053. [Google Scholar] [CrossRef]
- Xie, Y.; Nishi, S.; Iguchi, S.; Imai, N.; Sakatsume, M.; Saito, A.; Ikegame, M.; Iino, N.; Shimada, H.; Ueno, M. Expression of osteopontin in gentamicin-induced acute tubular necrosis and its recovery process. Kidney Int. 2001, 59, 959–974. [Google Scholar] [CrossRef]
- Hampel, D.J.; Sansome, C.; Romanov, V.I.; Kowalski, A.J.; Denhardt, D.T.; Goligorsky, M.S. Osteopontin traffic in hypoxic renal epithelial cells. Nephron Exp. Nephrol. 2003, 94, e66–e76. [Google Scholar] [CrossRef]
- Kim, H.; Sung, J.; Bae, J.Y.; Lee, P.; Oh, Y.K.; Kim, H. Identification of osteopontin as a urinary biomarker for autosomal dominant polycystic kidney disease progression. Kidney Res. Clin. Pract. 2022, 41, 730. [Google Scholar] [CrossRef]
- Giachelli, C.M.; Pichler, R.; Lombardi, D.; Denhardt, D.T.; Alpers, C.E.; Schwartz, S.M.; Johnson, R.J. Osteopontin expression in angiotensin II-induced tubulointerstitial nephritis. Kidney Int. 1994, 45, 515–524. [Google Scholar] [CrossRef]
- Seier, A.M.; Renkl, A.C.; Schulz, G.; Uebele, T.; Sindrilaru, A.; Iben, S.; Liaw, L.; Kon, S.; Uede, T.; Weiss, J.M. Antigen-specific induction of osteopontin contributes to the chronification of allergic contact dermatitis. Am. J. Pathol. 2010, 176, 246–258. [Google Scholar] [CrossRef]
- Guan, H.; Nagarkatti, P.S.; Nagarkatti, M. Role of CD44 in the differentiation of Th1 and Th2 cells: CD44-deficiency enhances the development of Th2 effectors in response to sheep RBC and chicken ovalbumin. J. Immunol. 2009, 183, 172–180. [Google Scholar] [CrossRef]
- Ashkar, S.; Weber, G.F.; Panoutsakopoulou, V.; Sanchirico, M.E.; Jansson, M.; Zawaideh, S.; Rittling, S.R.; Denhardt, D.T.; Glimcher, M.J.; Cantor, H. Eta-1 (osteopontin): An early component of type-1 (cell-mediated) immunity. Science 2000, 287, 860–864. [Google Scholar] [CrossRef]
- Iguchi, S.; Nishi, S.; Ikegame, M.; Hoshi, K.; Yoshizawa, T.; Kawashima, H.; Arakawa, M.; Ozawa, H.; Gejyo, F. Expression of osteopontin in cisplatin-induced tubular injury. Nephron Exp. Nephrol. 2004, 97, e96–e105. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| NE | ros-NE | p Value | |
|---|---|---|---|
| Sex (m/f) | 36/3 | 21/1 | 1 * |
| Age (years) | 36.23 ± 11.08 | 36.95 ± 12.63 | 0.93 § |
| Fever 1 (days) | 6.44 ± 2.73 | 5.64 ± 2.15 | 0.27 § |
| Fever 2 (days) | 0.82 ± 1.70 | 0.00 ± 0.00 | 0.01 § |
| Lower back pain (days) | 6.54 ± 4.25 | 4.09 ± 2.16 | 0.02 § |
| Vomiting (yes/no) | 12/27 | 7/15 | 1 * |
| Diarrhea (yes/no) | 15/24 | 3/19 | 0.047 * |
| Headache (days) | 6.21 ± 2.91 | 3.91 ± 1.02 | 0.001 § |
| Insomnia (days) | 2.58 ± 3.09 | 1.82 ± 1.76 | 0.6 § |
| Days | NE | Control | NE vs. Control (p Value) | ros-NE | ros-Control | ros-NE vs. ros-Control (p Value) | NE vs. ros-NE (p Value) | |
|---|---|---|---|---|---|---|---|---|
| Urea (mM/L) | 1 | 10.77 ± 8.95 | 4.42 ± 1.07 | <0.001 | 8.07 ± 3.70 | 4.17 ± 0.65 | 0.005 | 0.77 |
| 5 | 11.46 ± 9.65 | <0.001 | 8.27 ± 4.35 | 0.002 | 0.93 | |||
| 10 | 5.27 ± 1.65 | 0.10 | 5.72 ± 2.62 | 0.07 | 0.91 | |||
| Creatinine (mg/dL) | 1 | 210.77 ± 206.13 | 85.22 ± 11.84 | <0.001 | 156.09 ± 75.21 | 86.14 ± 10.19 | 0.002 | 0.29 |
| 5 | 204.18 ± 178.03 | <0.001 | 156.64 ± 96.35 | 0.002 | 0.95 | |||
| 10 | 105.95 ± 34.06 | <0.001 | 100.36 ± 21.32 | 0.17 | 0.30 | |||
| ALT (units/L) | 1 | 51.35 ± 32.27 | 20.04 ± 4.32 | <0.001 | 71.03 ± 34.65 | 16.71 ± 4.42 | <0.001 | 0.21 |
| 5 | 58.26 ± 41.74 | <0.001 | 68.79 ± 36.93 | <0.001 | 0.36 | |||
| 10 | 71.78 ± 81.72 | <0.001 | 80.00 ± 58.50 | <0.001 | 0.60 | |||
| AST (units/L) | 1 | 65.88 ± 58.06 | 22.35 ± 3.47 | <0.001 | 72.66 ± 38.62 | 20.29 ± 4.50 | <0.001 | 0.25 |
| 5 | 53.51 ± 28.53 | <0.001 | 61.30 ± 26.64 | <0.001 | 0.28 | |||
| 10 | 46.09 ± 28.93 | <0.001 | 46.50 ± 25.73 | <0.001 | 0.72 | |||
| Leukocytes (cells/μL ×109) | 1 | 9.91 ± 3.45 | 10.96 ± 3.35 | 0.24 | ||||
| 5 | 10.21 ± 3.58 | 11.21 ± 3.34 | 0.16 | |||||
| 10 | 9.16 ± 2.25 | 9.55 ± 2.27 | 0.56 | |||||
| Platelets (cells/μL) | 1 | 91.05 ± 54.74 | 91.05 ± 61.51 | 0.82 | ||||
| 5 | 171.00 ± 101.25 | 170.14 ± 76.59 | 0.74 | |||||
| 10 | 261.25 ± 95.92 | 272.09 ± 54.93 | 0.97 | |||||
| Hemoglobin (g/dL) | 1 | 144.92 ± 29.55 | 154.09 ± 21.17 | 0.37 | ||||
| 5 | 135.56 ± 15.73 | 141.59 ± 14.26 | 0.16 | |||||
| 10 | 139.21 ± 10.67 | 143.86 ± 14.67 | 0.23 | |||||
| ESR (mm/h) | 1 | 14.39 ± 9.83 | 14.59 ± 4.82 | 0.38 | ||||
| 5 | 19.41 ± 14.38 | 14.00 ± 5.68 | 0.43 | |||||
| 10 | 19.18 ± 11.74 | 12.77 ± 5.48 | 0.11 | |||||
| Urine protein (mg/dL) | 1 | 0.70 ± 0.90 | 0.83 ± 2.07 | 0.85 | ||||
| 5 | 0.07 ± 0.17 | 0.01 ± 0.06 | 0.09 | |||||
| Urine gravity | 1 | 1014.92 ± 7.48 | 1015.14 ± 8.45 | 0.98 | ||||
| 5 | 1009.59 ± 5.88 | 1008.23 ± 4.70 | 0.44 | |||||
| 10 | 1014.92 ± 4.94 | 1016.05 ± 5.28 | 0.50 | |||||
| Urine volume (ml/day) | 1 | 883.46 ± 706.31 | 938.18 ± 577.58 | 0.57 | ||||
| 5 | 2152.44 ± 1150.92 | 1976.82 ± 1437.28 | 0.26 | |||||
| 10 | 2188.21 ± 892.52 | 2210.91 ± 1023.77 | 0.74 | |||||
| Potassium (mEq/L) | 1 | 4.06 ± 0.53 | 4.32 ± 0.53 | 0.08 | ||||
| 5 | 4.16 ± 0.53 | 4.39 ± 0.58 | 0.15 | |||||
| 10 | 4.20 ± 0.36 | 4.28 ± 0.54 | 0.61 | |||||
| antibody (PC) | IgM IgG | 11.6 ± 5.2 13.1 ± 7.4 | 13.8 ± 3.6 19.2 ± 8.0 | 0.04 0.005 |
| Days | NE | Control | NE vs. Control (p Value) | ros-NE | ros-Control | ros-NE vs. ros-Control (p Value) | NE vs. ros-NE (p Value) | Control vs. ros-Control (p Value) | |
|---|---|---|---|---|---|---|---|---|---|
| HDL-C (mM/L) | 5 | 0.92 ± 0.18 | 2.56 ± 0.82 | >0.001 | 0.84 ± 0.25 | 1.82 ± 0.65 | 0.001 | 0.40 | 0.47 |
| 10 | 1.04 ± 0.21 | 2.50 ± 1.30 | >0.001 | 1.01 ± 0.37 | 1.41 ± 0.64 | 0.11 | 0.34 | 0.12 | |
| LDL-C (mM/L) | 5 | 2.40 ± 0.81 | 3.13 ± 1.05 | 0.14 | 1.64 ± 0.76 | 2.86 ± 1.03 | 0.004 | 0.003 | 0.67 |
| 10 | 2.59 ± 0.88 | 3.26 ± 1.02 | 0.27 | 1.87 ± 1.03 | 2.77 ± 1.80 | 0.27 | 0.01 | 0.25 | |
| Triglyceride (mM/L) | 5 | 2.64 ± 1.70 | 0.82 ± 0.16 | >0.001 | 2.46 ± 0.93 | 0.80 ± 0.22 | >0.001 | 0.74 | 0.99 |
| 10 | 2.87 ± 1.60 | 1.15 ± 0.38 | >0.001 | 2.55 ± 0.89 | 1.27 ± 0.53 | 0.004 | 0.97 | 0.77 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shakirova, V.; Markelova, M.; Davidyuk, Y.; Stott-Marshall, R.J.; Foster, T.L.; Khaiboullina, S.; Rizvanov, A.; Martynova, E. Rosuvastatin as a Supplemental Treatment for the Clinical Symptoms of Nephropathia Epidemica: A Pilot Clinical Study. Viruses 2024, 16, 306. https://doi.org/10.3390/v16020306
Shakirova V, Markelova M, Davidyuk Y, Stott-Marshall RJ, Foster TL, Khaiboullina S, Rizvanov A, Martynova E. Rosuvastatin as a Supplemental Treatment for the Clinical Symptoms of Nephropathia Epidemica: A Pilot Clinical Study. Viruses. 2024; 16(2):306. https://doi.org/10.3390/v16020306
Chicago/Turabian StyleShakirova, Venera, Maria Markelova, Yuriy Davidyuk, Robert J. Stott-Marshall, Toshana L. Foster, Svetlana Khaiboullina, Albert Rizvanov, and Ekaterina Martynova. 2024. "Rosuvastatin as a Supplemental Treatment for the Clinical Symptoms of Nephropathia Epidemica: A Pilot Clinical Study" Viruses 16, no. 2: 306. https://doi.org/10.3390/v16020306