
A Robust Phenotypic High-Throughput Antiviral Assay for the Discovery of Rabies Virus Inhibitors
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Viruses, and Library
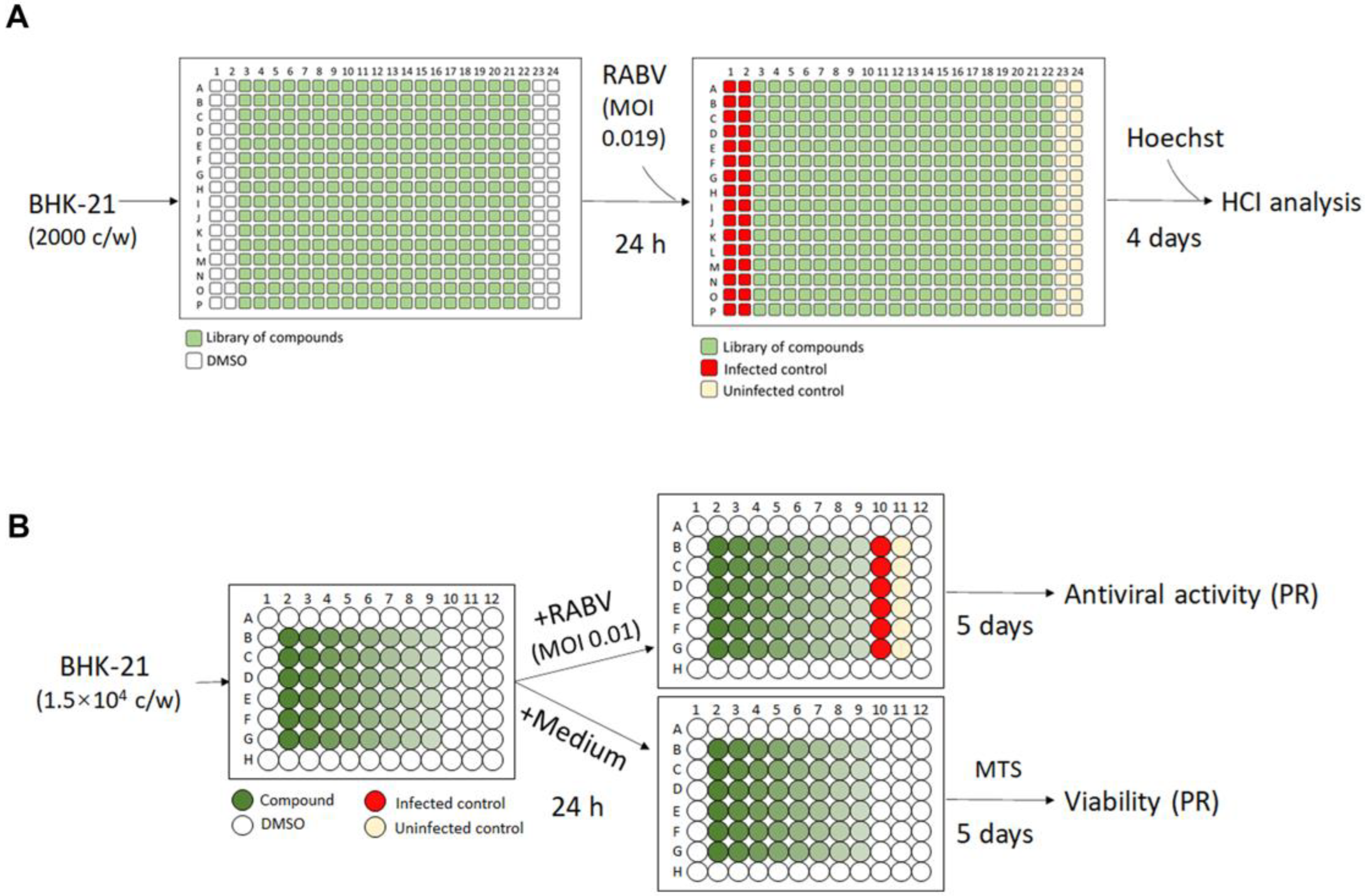
2.2. Optimized Protocol for 384-Well HTS Antiviral Assay and Image Analysis
2.3. Antiviral Assay for Hit Confirmation
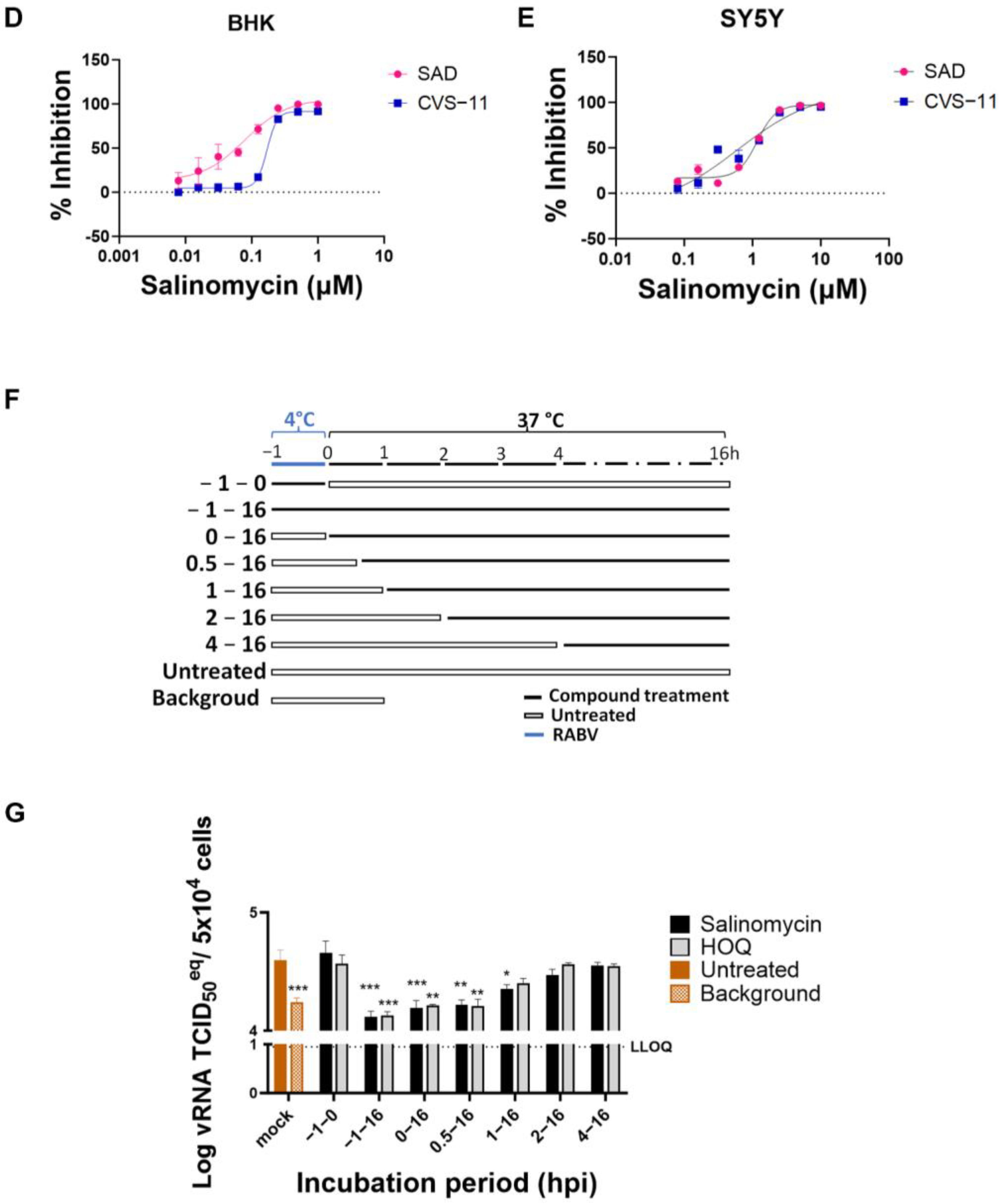
2.4. Time of Drug Addition Assay
2.5. Real-Time RT-qPCR
2.6. Statistics
3. Results
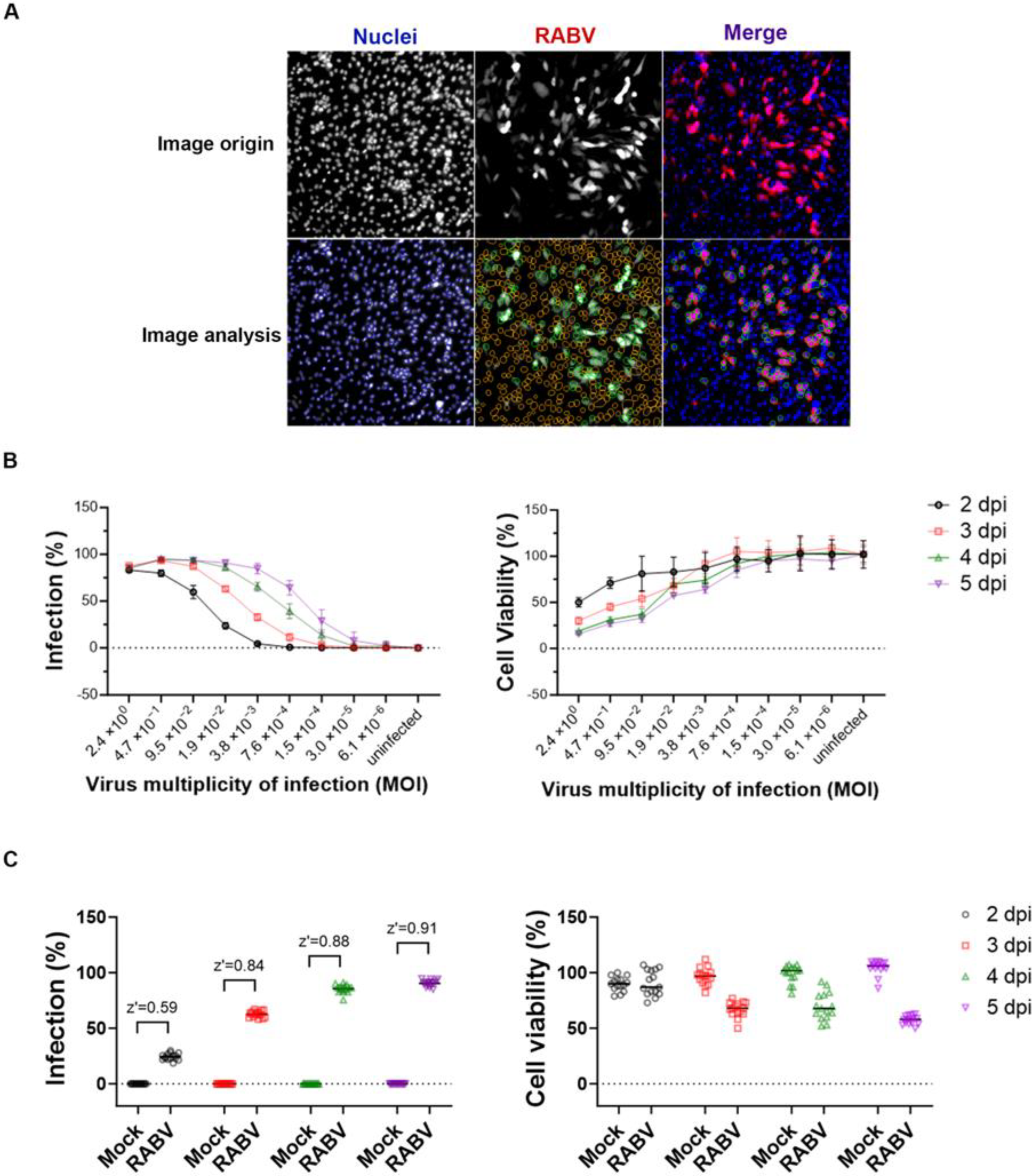
3.1. Optimization of a High-Throughput Phenotypic Anti-RABV Assay
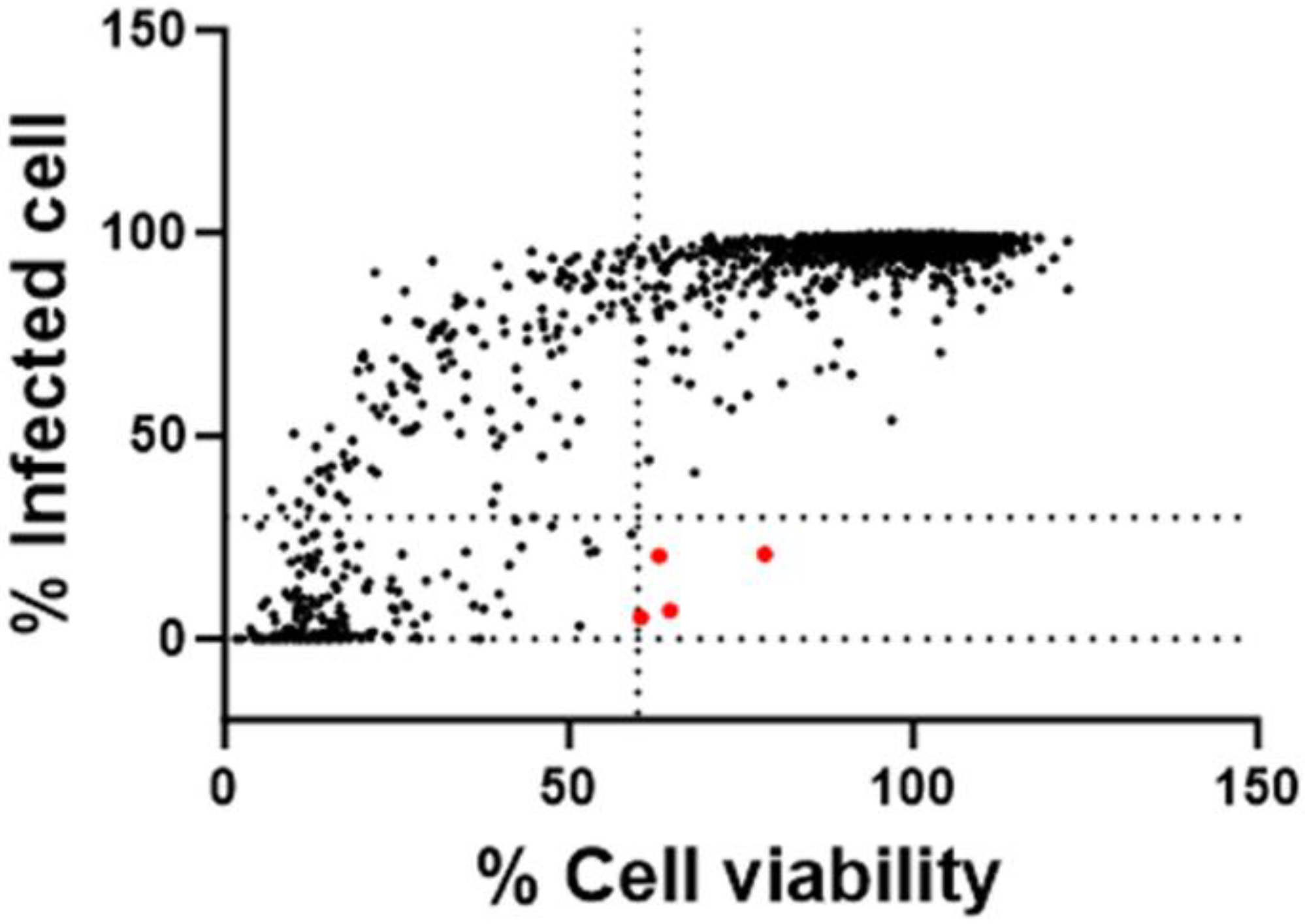
3.2. Antiviral Screening of a Repurposing Library
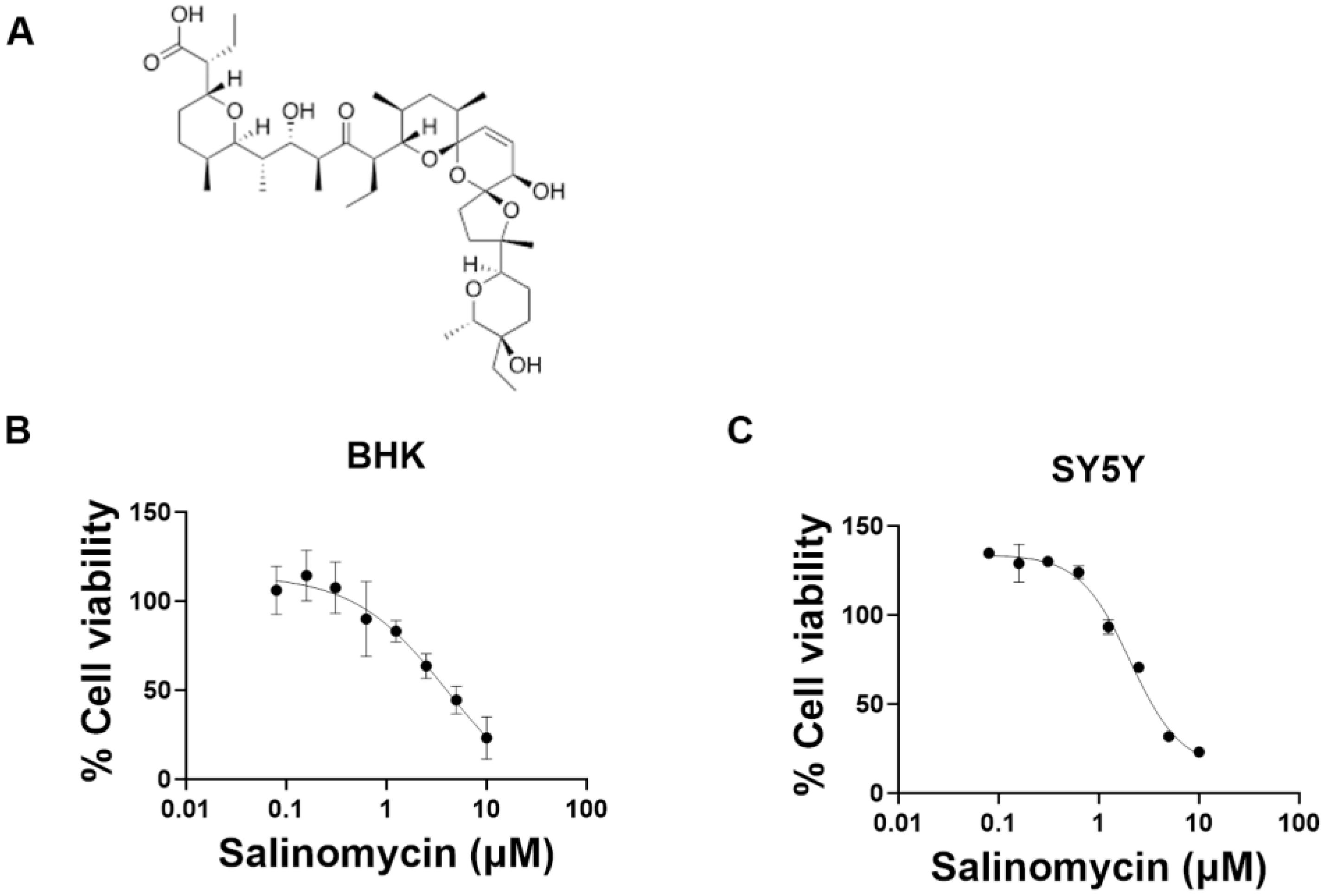
3.3. Validation of the Hit Compounds
3.4. Salinomycin Inhibits RABV at an Early Stage of the Replication Cycle
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fooks, A.R.; Cliquet, F.; Finke, S.; Freuling, C.; Hemachudha, T.; Mani, R.S.; Müller, T.; Nadin-Davis, S.; Picard-Meyer, E.; Wilde, H.; et al. Rabies. Nat. Rev. Dis. Prim. 2017, 3, 17091. [Google Scholar] [CrossRef] [PubMed]
- Badrane, H.; Bahloul, C.; Perrin, P.; Tordo, N. Evidence of Two Lyssavirus Phylogroups with Distinct Pathogenicity and Immunogenicity. J. Virol. 2001, 75, 3268–3276. [Google Scholar] [CrossRef] [PubMed]
- Ledesma, L.A.; Lemos, E.R.S.; Horta, M.A. Comparing Clinical Protocols for the Treatment of Human Rabies: The Milwaukee Protocol and the Brazilian Protocol (Recife). Rev. Soc. Bras. Med. Trop. 2020, 53, e20200352. [Google Scholar] [CrossRef]
- Merigan, T.C.; Baer, G.M.; Winkler, W.G.; Bernard, K.W.; Gibert, C.G.; Chany, C.; Veronesi, R. Human Leukocyte Interferon Administration to Patients with Symptomatic and Suspected Rabies. Ann. Neurol. 1984, 16, 82–87. [Google Scholar] [CrossRef]
- Banyard, A.C.; Mansfield, K.L.; Wu, G.; Selden, D.; Thorne, L.; Birch, C.; Koraka, P.; Osterhaus, A.D.M.E.; Fooks, A.R. Re-Evaluating the Effect of Favipiravir Treatment on Rabies Virus Infection. Vaccine 2019, 37, 4686–4693. [Google Scholar] [CrossRef] [PubMed]
- Ben Mechlia, M.; Belaid, A.; Castel, G.; Jallet, C.; Mansfield, K.L.; Fooks, A.R.; Hani, K.; Tordo, N. Dermaseptins as Potential Antirabies Compounds. Vaccine 2019, 37, 4694–4700. [Google Scholar] [CrossRef] [PubMed]
- Warrell, M.J.; White, N.J.; Looareesuwan, S.; Phillips, R.E.; Sunthrasamai, P.; Chanthavanich, P.; Riganti, M.; Fisher-Hoch, S.P.; Nicholson, K.G.; Manatsathit, S.; et al. Failure of Interferon Alfa and Tribavirin in Rabies Encephalitis. Br. Med. J. 1989, 299, 830–833. [Google Scholar] [CrossRef]
- Smreczak, M.; Orłowska, A.; Marzec, A.; Trębas, P.; Kycko, A.; Reichert, M.; Koraka, P.; Osterhaus, A.D.M.E.; Żmudziński, J.F. The Effect of Combined Drugs Therapy on the Course of Clinical Rabies Infection in a Murine Model. Vaccine 2019, 37, 4701–4709. [Google Scholar] [CrossRef]
- Chávez, J.H.; Leal, P.C.; Yunes, R.A.; Nunes, R.J.; Barardi, C.R.M.; Pinto, A.R.; Simões, C.M.O.; Zanetti, C.R. Evaluation of Antiviral Activity of Phenolic Compounds and Derivatives against Rabies Virus. Vet. Microbiol. 2006, 116, 53–59. [Google Scholar] [CrossRef]
- Lingappa, U.F.; Wu, X.; Macieik, A.; Yu, S.F.; Atuegbu, A.; Corpuz, M.; Francis, J.; Nichols, C.; Calayag, A.; Shi, H.; et al. Host-Rabies Virus Protein-Protein Interactions as Druggable Antiviral Targets. Proc. Natl. Acad. Sci. USA 2013, 110, E861–E868. [Google Scholar] [CrossRef]
- Rogée, S.; Larrous, F.; Jochmans, D.; Ben-Khalifa, Y.; Neyts, J.; Bourhy, H. Pyrimethamine Inhibits Rabies Virus Replication in Vitro. Antivir. Res. 2019, 161, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Anindita, P.D.; Sasaki, M.; Okada, K.; Ito, N.; Sugiyama, M.; Saito-Tarashima, N.; Minakawa, N.; Shuto, S.; Otsuguro, S.; Ichikawa, S.; et al. Ribavirin-Related Compounds Exert in Vitro Inhibitory Effects toward Rabies Virus. Antivir. Res. 2018, 154, 1–9. [Google Scholar] [CrossRef]
- Yamada, K.; Noguchi, K.; Komeno, T.; Furuta, Y.; Nishizono, A. Efficacy of Favipiravir (T-705) in Rabies Postexposure Prophylaxis. J. Infect. Dis. 2016, 213, 1253–1261. [Google Scholar] [CrossRef] [PubMed]
- Kimitsuki, K.; Khan, S.; Kaimori, R.; Yahiro, T.; Saito, N.; Yamada, K.; Nakajima, N.; Komeno, T.; Furuta, Y.; Quiambao, B.P.; et al. Implications of the Antiviral Drug Favipiravir on Rabies Immunoglobulin for Post-Exposure Prophylaxis of Rabies in Mice Model with Category III-like Exposures. Antivir. Res. 2023, 209, 105489. [Google Scholar] [CrossRef] [PubMed]
- Poole, C.L.; James, S.H. Antiviral Therapies for Herpesviruses: Current Agents and New Directions. Clin. Ther. 2018, 40, 1282–1298. [Google Scholar] [CrossRef] [PubMed]
- Gulick, R.M.; Flexner, C. Long-Acting HIV Drugs for Treatment and Prevention. Annu. Rev. Med. 2019, 70, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Chien, R.N.; Liaw, Y.F. Current Trend in Antiviral Therapy for Chronic Hepatitis B. Viruses 2022, 14, 434. [Google Scholar] [CrossRef] [PubMed]
- Abu-Freha, N.; Mathew Jacob, B.; Elhoashla, A.; Afawi, Z.; Abu-Hammad, T.; Elsana, F.; Paz, S.; Etzion, O. Chronic Hepatitis C: Diagnosis and Treatment Made Easy. Eur. J. Gen. Pract. 2022, 28, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Perez, J.; Schafer, A.; Cheng, H.; Peet, N.; Rong, L.; Manicassamy, B. Influenza Virus: Small Molecule Therapeutics and Mechanisms of Antiviral Resistance. Curr. Med. Chem. 2018, 25, 5115–5127. [Google Scholar] [CrossRef]
- Fernandes, Q.; Inchakalody, V.P.; Merhi, M.; Mestiri, S.; Taib, N.; Moustafa Abo El-Ella, D.; Bedhiafi, T.; Raza, A.; Al-Zaidan, L.; Mohsen, M.O.; et al. Emerging COVID-19 Variants and Their Impact on SARS-CoV-2 Diagnosis, Therapeutics and Vaccines. Ann. Med. 2022, 54, 524–540. [Google Scholar] [CrossRef]
- Gao, M.; Nettles, R.E.; Belema, M.; Snyder, L.B.; Nguyen, V.N.; Fridell, R.A.; Serrano-Wu, M.H.; Langley, D.R.; Sun, J.H.; O’Boyle, D.R.; et al. Chemical Genetics Strategy Identifies an HCV NS5A Inhibitor with a Potent Clinical Effect. Nature 2010, 465, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Goethals, O.; Kaptein, S.J.F.; Kesteleyn, B.; Bonfanti, J.F.; Van Wesenbeeck, L.; Bardiot, D.; Verschoor, E.J.; Verstrepen, B.E.; Fagrouch, Z.; Putnak, J.R.; et al. Blocking NS3–NS4B Interaction Inhibits Dengue Virus in Non-Human Primates. Nature 2023, 615, 678–686. [Google Scholar] [CrossRef] [PubMed]
- Kaptein, S.J.F.; Goethals, O.; Kiemel, D.; Marchand, A.; Kesteleyn, B.; Bonfanti, J.F.; Bardiot, D.; Stoops, B.; Jonckers, T.H.M.; Dallmeier, K.; et al. A Pan-Serotype Dengue Virus Inhibitor Targeting the NS3–NS4B Interaction. Nature 2021, 598, 504–509. [Google Scholar] [CrossRef]
- Smith, S.P.; Shipley, R.; Drake, P.; Fooks, A.R.; Ma, J.; Banyard, A.C. Characterisation of a Live-Attenuated Rabies Virus Expressing a Secreted ScFv for the Treatment of Rabies. Viruses 2023, 15, 1674. [Google Scholar] [CrossRef]
- ter Horst, S.; Chiu, W.; Neyts, J.; Rocha-Pereira, J. Screening and in Vitro Antiviral Assessment of Small Molecules against Fluorescent Protein-Expressing Bunyamwera Virus in a Cell-Based Assay Using High-Content Imaging. Antivir. Chem. Chemother. 2021, 29, 1–8. [Google Scholar] [CrossRef]
- Wang, X.; Terrie, L.; Wu, G.; Van Damme, E.J.M.; Thorrez, L.; Fooks, A.R.; Banyard, A.C.; Jochmans, D.; Neyts, J. Urtica Dioica Agglutinin Prevents Rabies Virus Infection in a Muscle Explant Model. Pharmaceutics 2023, 15, 1353. [Google Scholar] [CrossRef] [PubMed]
- Jochmans, D.; Leyssen, P.; Neyts, J. A Novel Method for High-Throughput Screening to Quantify Antiviral Activity against Viruses That Induce Limited CPE. J. Virol. Methods 2012, 183, 176–179. [Google Scholar] [CrossRef]
- Antoszczak, M.; Huczyński, A. Salinomycin and Its Derivatives—A New Class of Multiple-Targeted “Magic Bullets”. Eur. J. Med. Chem. 2019, 176, 208–227. [Google Scholar] [CrossRef]
- Wu, J.; Cao, S.; Lei, S.; Liu, Q.; Li, Y.; Yu, Y.; Xie, H.; Li, Q.; Zhao, X.; Chen, R.; et al. Clofazimine: A Promising Inhibitor of Rabies Virus. Front. Pharmacol. 2021, 12, 598241. [Google Scholar] [CrossRef]
- Du Pont, V.; Wirblich, C.; Yoon, J.-J.; Cox, R.M.; Schnell, M.J.; Plemper, R.K. Identification and Characterization of a Small-Molecule Rabies Virus Entry Inhibitor. J. Virol. 2020, 94, 10–1128. [Google Scholar] [CrossRef]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug Repurposing: Progress, Challenges and Recommendations. Nat. Rev. Drug Discov. 2018, 18, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Hickey, E.E.; Wong, H.S.; Khazandi, M.; Ogunniyi, A.D.; Petrovski, K.R.; Garg, S.; Page, S.W.; O’Handley, R.; Trott, D.J. Repurposing Ionophores as Novel Antimicrobial Agents for the Treatment of Bovine Mastitis Caused by Gram-Positive Pathogens. J. Vet. Pharmacol. Ther. 2018, 41, 746–754. [Google Scholar] [CrossRef]
- Antoszczak, M.; Maj, E.; Stefańska, J.; Wietrzyk, J.; Janczak, J.; Brzezinski, B.; Huczyński, A. Synthesis, Antiproliferative and Antibacterial Activity of New Amides of Salinomycin. Bioorg. Med. Chem. Lett. 2014, 24, 1724–1729. [Google Scholar] [CrossRef]
- Jang, Y.; Shin, J.S.; Yoon, Y.-S.; Go, Y.Y.; Lee, H.W.; Kwon, O.S.; Park, S.; Park, M.-S.; Kim, M.; Soo Shin, J.; et al. Salinomycin Inhibits Influenza Virus Infection by Disrupting Endosomal Acidification and Viral Matrix Protein 2 Function. J. Virol. 2018, 92, 1441–1459. [Google Scholar] [CrossRef] [PubMed]
- Dittmar, M.; Lee, J.S.; Whig, K.; Segrist, E.; Li, M.; Kamalia, B.; Castellana, L.; Ayyanathan, K.; Cardenas-Diaz, F.L.; Morrisey, E.E.; et al. Drug Repurposing Screens Reveal Cell-Type-Specific Entry Pathways and FDA-Approved Drugs Active against SARS-Cov-2. Cell Rep. 2021, 35, 108959. [Google Scholar] [CrossRef] [PubMed]
- Qi, D.; Liu, Y.; Li, J.; Huang, J.H.; Hu, X.; Wu, E. Salinomycin as a Potent Anticancer Stem Cell Agent: State of the Art and Future Directions. Med. Res. Rev. 2022, 42, 1037–1063. [Google Scholar] [CrossRef]
- Boehmerle, W.; Endres, M. Salinomycin Induces Calpain and Cytochrome C-Mediated Neuronal Cell Death. Cell Death Dis. 2011, 2, e168. [Google Scholar] [CrossRef]
- Story, P.; Doube, A. A Case of Human Poisoning by Salinomycin, an Agricultural Antibiotic. N. Z. Med. J. 2004, 117, U799. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus Strain | 1 EC50 (µM) | CC50 (µM) | ||
|---|---|---|---|---|
| BHK-21 | SH-SY5Y | BHK-21 | SH-SY5Y | |
| SAD B19 | 0.051 ± 0.020 | 0.90 ± 0.070 | 4.0 ± 0.18 | 4.0 ± 0.060 |
| CVS-11 | 0.18 ± 0.0040 | 0.67 ± 0.11 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Chiu, W.; Klaassen, H.; Marchand, A.; Chaltin, P.; Neyts, J.; Jochmans, D. A Robust Phenotypic High-Throughput Antiviral Assay for the Discovery of Rabies Virus Inhibitors. Viruses 2023, 15, 2292. https://doi.org/10.3390/v15122292
Wang X, Chiu W, Klaassen H, Marchand A, Chaltin P, Neyts J, Jochmans D. A Robust Phenotypic High-Throughput Antiviral Assay for the Discovery of Rabies Virus Inhibitors. Viruses. 2023; 15(12):2292. https://doi.org/10.3390/v15122292
Chicago/Turabian StyleWang, Xinyu, Winston Chiu, Hugo Klaassen, Arnaud Marchand, Patrick Chaltin, Johan Neyts, and Dirk Jochmans. 2023. "A Robust Phenotypic High-Throughput Antiviral Assay for the Discovery of Rabies Virus Inhibitors" Viruses 15, no. 12: 2292. https://doi.org/10.3390/v15122292