
Human Intramuscular Hyperimmune Gamma Globulin (hIHGG) Anti-SARS-CoV-2—Characteristics of Intermediates and Final Product

 , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Production of hIHGG Anti-SARS-CoV-2
2.2. Methods of Measuring Anti-SARS-CoV-2 Antibodies
2.2.1. Immunoenzymatic Methods
- SARS-CoV-2 Ab ELISA Test (Wantai, Beijing, China)—for simultaneous detection of three antibody isotypes (IgM, IgA, IgG) directed to the S1-RBD;
- SARS-CoV-2 Ab IgM ELISA (Wantai, Beijing, China)—for detecting the IgM isotype of specific antibodies directed to the S1-RBD;
- Anti-SARS-CoV-2 IgA ELISA (EUROIMMUN, Lübeck, Germany)—for semi-quantitative in vitro evaluation of anti-SARS-CoV-2 IgA antibodies directed to the S1 protein;
- Anti-SARS-CoV-2 ELISA (IgG) (EUROIMMUN, Lübeck, Germany)—for semi-quantitative in vitro evaluation of anti-SARS-CoV-2 IgG antibodies directed to the S1 protein. The test was used in 2020, and since December 2020, anti-SARS-CoV-2 QuantiVac ELISA (IgG) (EUROIMMUN, Lübeck, Germany) quantitative test was in use;
- iFlash–SARS-CoV-2 IgG—chemiluminescence test for semi-quantitative detection of anti-SARS-CoV-2 IgG antibodies (SHENZHEN YHLO BIOTECH CO., Shenzhen, China);
- Bio-Rad Platelia SARS-CoV-2 Total Ab—for simultaneous detection of IgG, IgA, and IgM antibodies directed to the nucleocapsid (anti-N).
2.2.2. Neutralization Tests
2.2.3. Quantitative and Semi-Quantitative Antibody Concentration Assay
2.2.4. Stability Studies
2.2.5. Statistical Analysis
3. Results
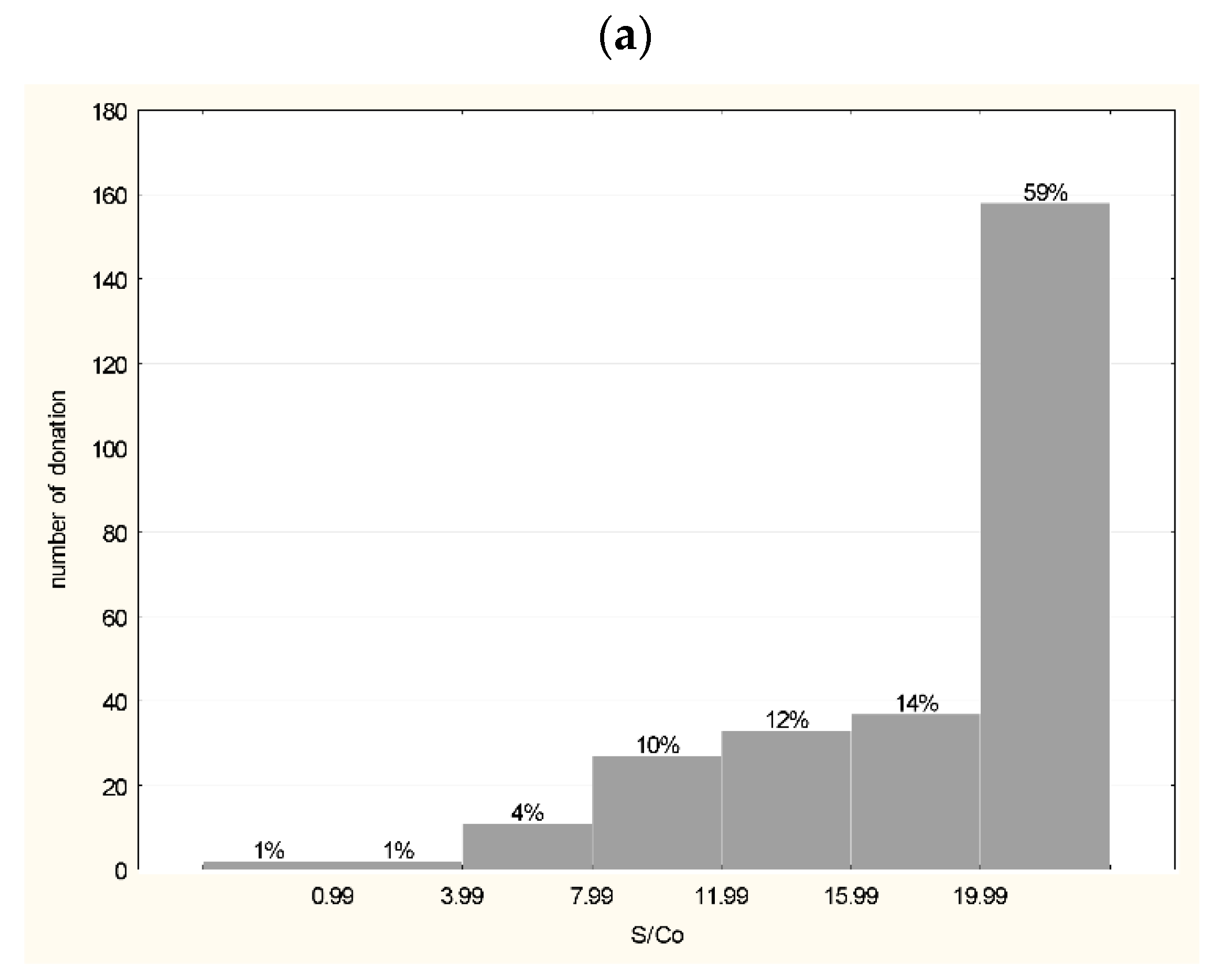
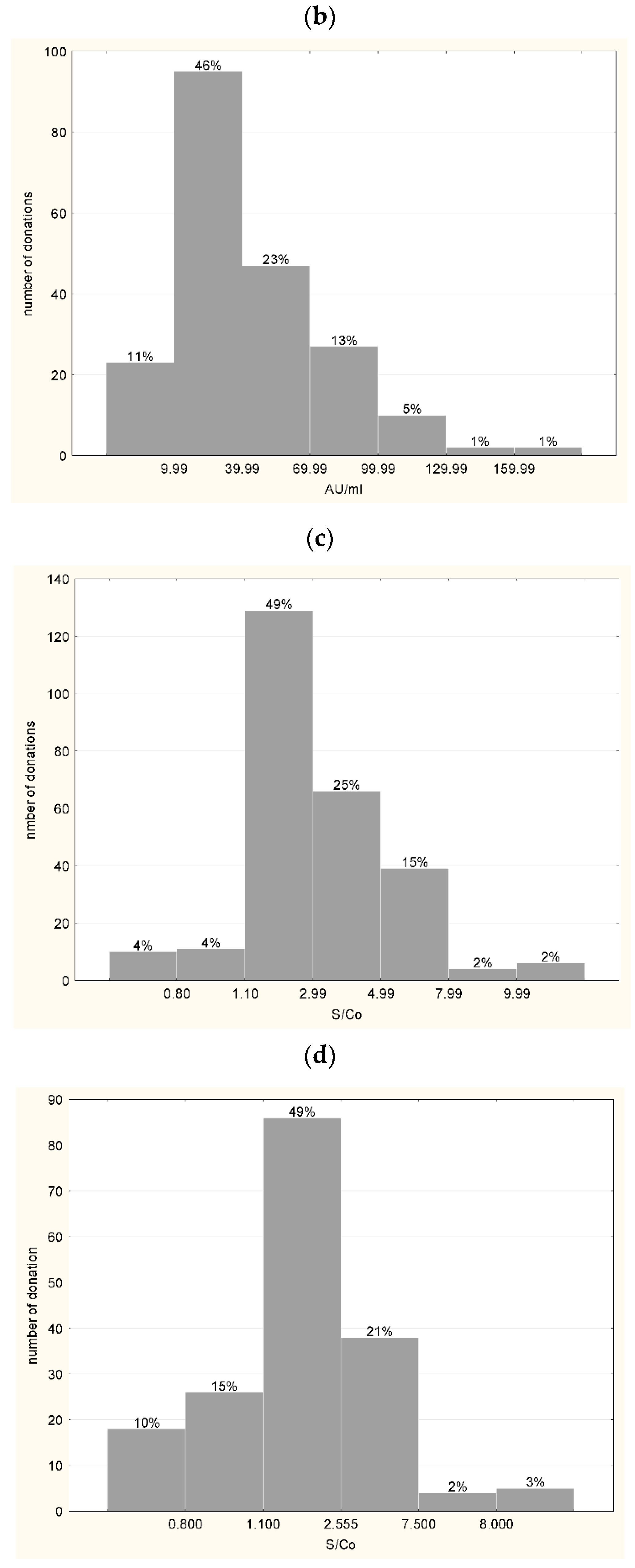
3.1. Characteristics of Plasma Used for the Production of hIHGG Anti-SARS-CoV-2
3.2. Characteristics of Intermediates
- A.
- The plasma pool
- B.
- Concentrate after fractionation, before lyophilization
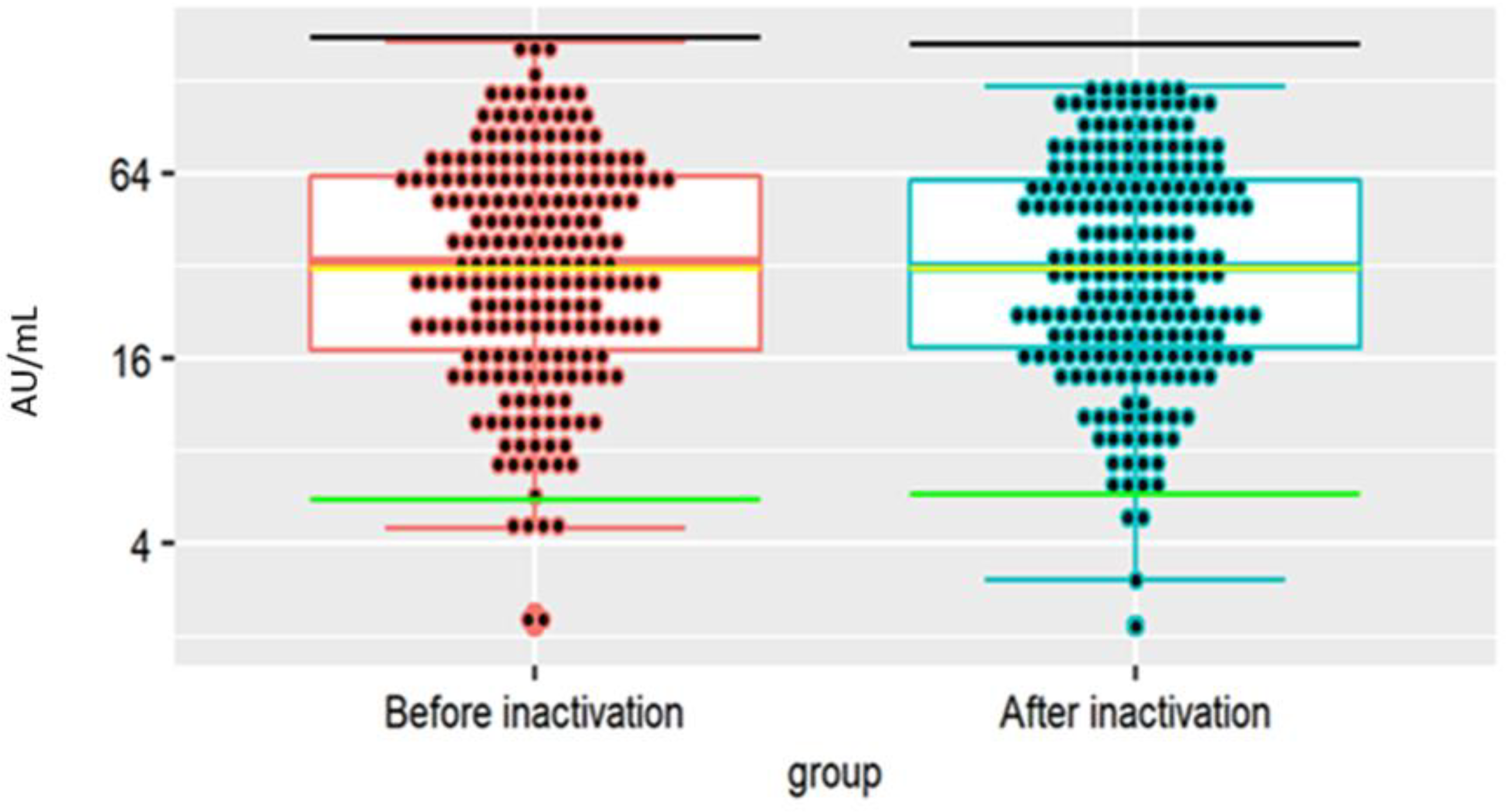
3.3. Testing the Antibodies in the FP before © and after Ampoule Filling (D)
3.4. Physico-Chemical Characteristics of hIHGG Anti-SARS-CoV-2
3.5. Product Stability
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tea, F.; Ospina Stella, A.; Aggarwal, A.; Ross Darley, D.; Pilli, D.; Vitale, D.; Merheb, V.; Lee, F.X.; Cunningham, P.; Walker, G.J.; et al. SARS-CoV-2 neutralizing antibodies: Longevity, breadth, and evasion by emerging viral variants. PLoS Med. 2021, 18, e1003656. [Google Scholar] [CrossRef] [PubMed]
- Valdez-Cruz, N.A.; García-Hernández, E.; Espitia, C.; Cobos-Marín, L.; Altamirano, C.; Bando-Campos, C.G.; Cofas-Vargas, L.F.; Coronado-Aceves, E.W.; González-Hernández, R.A.; Hernández-Peralta, P.; et al. Integrative overview of antibodies against SARS-CoV-2 and their possible applications in COVID-19 prophylaxis and treatment. Microb. Cell Factories 2021, 20, 88. [Google Scholar] [CrossRef] [PubMed]
- Wooding, D.J.; Bach, H. Treatment of COVID-19 with convalescent plasma: Lessons from past coronavirus outbreaks. Clin. Microbiol. Infect. 2020, 26, 1436–1446. [Google Scholar] [CrossRef] [PubMed]
- Tworek, A.; Jaroń, K.; Uszyńska-Kałuża, B.; Rydzewski, A.; Gil, R.; Deptała, A.; Franek, E.; Wójtowicz, R.; Życińska, K.; Walecka, I.; et al. Convalescent plasma treatment is associated with lower mortality and better outcomes in high-risk COVID-19 patients—Propensity-score matched case-control study. Int. J. Infect. Dis. 2021, 105, 209–215. [Google Scholar] [CrossRef]
- Rojas, M.; Anaya, J.-M. Why will it never be known if convalescent plasma is effective for COVID-19. J. Transl Autoimmun. 2020, 3, 100069. [Google Scholar] [CrossRef]
- Sullivan, D.J.; Gebo, K.A.; Shoham, S.; Bloch, E.M.; Lau, B.; Shenoy, A.G.; Mosnaim, G.S.; Gniadek, T.J.; Fukuta, Y.; Patel, B.; et al. Randomized Controlled Trial of Early Outpatient COVID-19 Treatment with High-Titer Convalescent Plasma. medRxiv. [CrossRef]
- Deeks, E.D. Casirivimab/Imdevimab: First Approval. Drugs 2021, 81, 2047–2055. [Google Scholar] [CrossRef]
- ITAC (INSIGHT 013) Study Group. Hyperimmune immunoglobulin for hospitalised patients with COVID-19 (ITAC): A double-blind, placebo-controlled, phase 3, randomised trial. Lancet 2022, 399, 530–540. [Google Scholar] [CrossRef]
- Parikh, D.; Chaturvedi, A.; Shah, N.; Patel, P.; Patel, R.; Ray, S. Safety and efficacy of COVID-19 hyperimmune globulin (HIG) solution in the treatment of active COVID-19 infection-Findings from a Prospective, Randomized, Controlled, Multi-Centric Trial. medRxiv. [CrossRef]
- Obwieszczenie Ministra Zdrowia z dnia 30 marca 2021, r. w sprawie wymagań dobrej praktyki pobierania krwi i jej składników, badania, preparatyki, przechowywania, wydawania i transportu dla jednostek organizacyjnych publicznej służby krwi [Announcement of the Minister of Health of 30th March 2021 Regarding Good Practice Requirements for Collection, Testing, Preparation, Storage, Distribution and Transport of Blood and Blood Components in Organizational Units of Public Blood Transfusion Service]. Available online: http://dziennikmz.mz.gov.pl/DUM_MZ/2021/28/akt.pdf (accessed on 6 March 2022).
- Antoniewicz-Papis, J.; Brojer, E.; Fabijańska-Mitek, J.; Gierszon, A.; Grabarczyk, P.; Guz, K.; Kalińska, A.; Kopacz, A.; Korsak, J.; Lachert, E.; et al. Current status and achievements of Polish transfusion medicine. Acta Haematol. Pol. 2021, 52, 147–162. [Google Scholar] [CrossRef]
- Grabarczyk, P.; Kopacz, A.; Sulkowska, E.; Kubicka-Russel, D.; Mikulska, M.; Brojer, E.; Łętowska, M. Blood donor screening for blood born viruses in Poland. Przegląd Epidemiol. 2015, 69, 473–477. [Google Scholar]
- Cohn, E.J.; Strong, L.E.; Hughes, W.L., Jr.; Mulford, D.J.; Ashworth, J.N.; Melin, M.; Taylor, H.L. Preparation and properties of serum and plasma proteins. A system for the separation into fractions of the protein and lipoprotein components of biological tissues and fluids. J. Am. Chem. Soc. 1946, 68, 459–475. [Google Scholar] [CrossRef] [PubMed]
- Holst, J. Convalescent serum in the treatment of influenza. Nor. Mag. Laegevidenskaben. 1919, 80, 31–561. [Google Scholar]
- Wong, V.; Dai, D.; Wu, A.; Sung, J.J. Treatment of severe acute respiratory syndrome with convalescent plasma. Hong Kong Med. J. 2003, 9, 199–201. [Google Scholar] [PubMed]
- WHO Mers-Cov Research Group. State of Knowledge and Data Gaps of Middle East Respiratory Syndrome Coronavirus (MERS-CoV) in Humans. PLoS Curr. 2013, 5. [Google Scholar] [CrossRef]
- Lasocka, J.; Bielawski, A.; Lachert, E. Passive immunization in the combat against infectious diseases (COVID-19 included). J. Transfus. Med. 2021, 14, 50–57. [Google Scholar] [CrossRef]
- Pandey, S.; Vyas, G.N. Adverse effects of plasma transfusion. Transfusion 2012, 52 (Suppl. 1), 65S–79S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzik, S. COVID-19 Convalescent Plasma: Now Is the Time for Better Science. Transfus. Med. Rev. 2020, 34, 141–144. [Google Scholar] [CrossRef]
- Driggin, E.; Madhavan, M.V.; Bikdeli, B.; Chuich, T.; Laracy, J.; Biondi-Zoccai, G.; Brown, T.S.; Der-Nigoghossian, C.; Zidar, D.A.; Haythe, J.; et al. Cardiovascular Considerations for Patients, Health Care Workers, and Health Systems During the COVID-19 Pandemic. J. Am. Coll. Cardiol. 2020, 75, 2352–2371. [Google Scholar] [CrossRef]
- O’Brien, S.F.; Lieshout-Krikke, R.W.; Lewin, A.; Erikstrup, C.; Steele, W.R.; Uzicanin, S.; Custer, B. Research initiatives of blood services worldwide in response to the covid-19 pandemic. Vox Sang. 2021, 116, 296–304. [Google Scholar] [CrossRef]
- Ali, S.; Uddin, S.M.; Ali, A.; Anjum, F.; Ali, R.; Shalim, E.; Khan, M.; Ahmed, I.; Muhaymin, S.M.; Bukhari, U. Production of hyperimmune anti-SARS-CoV-2 intravenous immunoglobulin from pooled COVID-19 convalescent plasma. Immunotherapy 2021, 13, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Vandeberg, P.; Cruz, M.; Diez, J.M.; Merritt, W.K.; Santos, B.; Trukawinski, S.; Wellhouse, A.; Jose, M.; Willis, T. Production of anti-SARS-CoV-2 hyperimmune globulin from convalescent plasma. Transfusion 2021, 61, 1705–1709. [Google Scholar] [CrossRef] [PubMed]
- Flegel, W.A. Pathogenesis and mechanisms of antibody-mediated hemolysis. Transfusion 2015, 55 (Suppl. 2), S47–S58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinti, I.; Mortari, E.P.; Fernandez Salinas, A. IgA Antibodies and IgA Deficiency in SARS-CoV-2 Infection. Front Cell. Infect. Microbiol. 2021, 11, 655896. [Google Scholar] [CrossRef]
- Rudowska, E.; Basta, L.; Piwowarska, G.; Drybańska, B.; Dylag, S. The epidemiological trends connected with detection of HIV, HCV, HBV and syphilis markers in blood donors in the years 2006–2009. Polski Merkur. Lek. 2011, 30, 181–186. [Google Scholar]
- Wiersum-Osselton, J.C. Risk factors for complications in donors at first and repeat whole blood donation: A cohort study with assessment of the impact on donor return. Blood Transfus. 2014, 12 (Suppl. 1), s28–s36. [Google Scholar]
- Raster, J.; Zimmermann, K.; Wesche, J.; Aurich, K.; Greinacher, A.; Selleng, K. Effect of Methylene Blue Pathogen Inactivation on the Integrity of Immunoglobulin M and G. Transfus. Med. Hemother. 2021, 48, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Kostin, A.; Lundgren, M.; Bulanov, A. Impact of pathogen reduction technologies on immunological properties of the COVID-19 convalescent plasma. medRxiv. [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test/Assay | Production Stage | Compaction Factor | |||
|---|---|---|---|---|---|
| Plasma Pool | Concentrate Prior to Liofilization | Product | |||
| Before Filling | After Filling | ||||
| A | B | C | D | D/A | |
| anti-RBD S1 total (Wantai) | 110.90 | 471.04 | 721.28 | 801.84 | 7.23 |
| anti-S1 IgG (EuroImmun) | 4.21 | 33.34 | 61.87 | 63.31 | 15.02 |
| anti-S1 IgA (EuroImmun) | 1 | undetectable | undetectable | undetectable | - |
| anti-RBD S1 IgM (Wantai) | 115.52 | 20 | 21.14 | 24.43 | 0.21 |
| anti-N total (BioRad) | 13.99 | 83.61 | 123.97 | 87.57 | 6.74 |
| Neutralization VN | 40 | 160 | nt. | 160 | 4 |
| PVN (IC50) | 63.71 | nt. | nt. | 313.55 | 4.92 |
| Month | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | |
| Anti-RBD S1 total Wantai (titer) | 981 | 899 | 503 | nt | 801 | nt | 868 | 640 | nt | 680 | 827 | 777 | 960 | |
| Anti-S1(IgG) Euroimmun (semiquantitative; titer) | 55 | 49 | 47 | |||||||||||
| Anti-S1(IgG) Euroimmun (quantitative) BAU/mL | 1765 | 1653 | 1667 | 1566 | 1606 | 2104 | 1855 | 1798 | 1833 | 1953 | 1650 | 1765 | ||
| Neutralization assay (VN) titer at 100% neutralization | 160 | 160 | 160 | |||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lachert, E.; Lasocka, J.; Bielawski, A.; Sulkowska, E.; Guz, K.; Pyrc, K.; Dabrowska, A.; Wawryniuk-Malmon, A.; Letowska, M.; Tomasiewicz, K.; et al. Human Intramuscular Hyperimmune Gamma Globulin (hIHGG) Anti-SARS-CoV-2—Characteristics of Intermediates and Final Product. Viruses 2022, 14, 1328. https://doi.org/10.3390/v14061328
Lachert E, Lasocka J, Bielawski A, Sulkowska E, Guz K, Pyrc K, Dabrowska A, Wawryniuk-Malmon A, Letowska M, Tomasiewicz K, et al. Human Intramuscular Hyperimmune Gamma Globulin (hIHGG) Anti-SARS-CoV-2—Characteristics of Intermediates and Final Product. Viruses. 2022; 14(6):1328. https://doi.org/10.3390/v14061328
Chicago/Turabian StyleLachert, Elzbieta, Joanna Lasocka, Artur Bielawski, Ewa Sulkowska, Katarzyna Guz, Krzysztof Pyrc, Agnieszka Dabrowska, Agata Wawryniuk-Malmon, Magdalena Letowska, Krzysztof Tomasiewicz, and et al. 2022. "Human Intramuscular Hyperimmune Gamma Globulin (hIHGG) Anti-SARS-CoV-2—Characteristics of Intermediates and Final Product" Viruses 14, no. 6: 1328. https://doi.org/10.3390/v14061328