
Identification and Molecular Characterization of Novel Mycoviruses in Saccharomyces and Non-Saccharomyces Yeasts of Oenological Interest

 , , , , ,
, , , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Isolate Maintenance
2.2. Total RNA Extraction and RNAseq
2.3. Transcriptome Assembly
2.4. ORFan Contig Detection
2.5. Virus Detection in Specific Isolates
2.6. ORFan Detection in Specific Isolates and DNA Integration Assay
2.7. ORF Prediction and Phylogenetic Analyses
3. Results
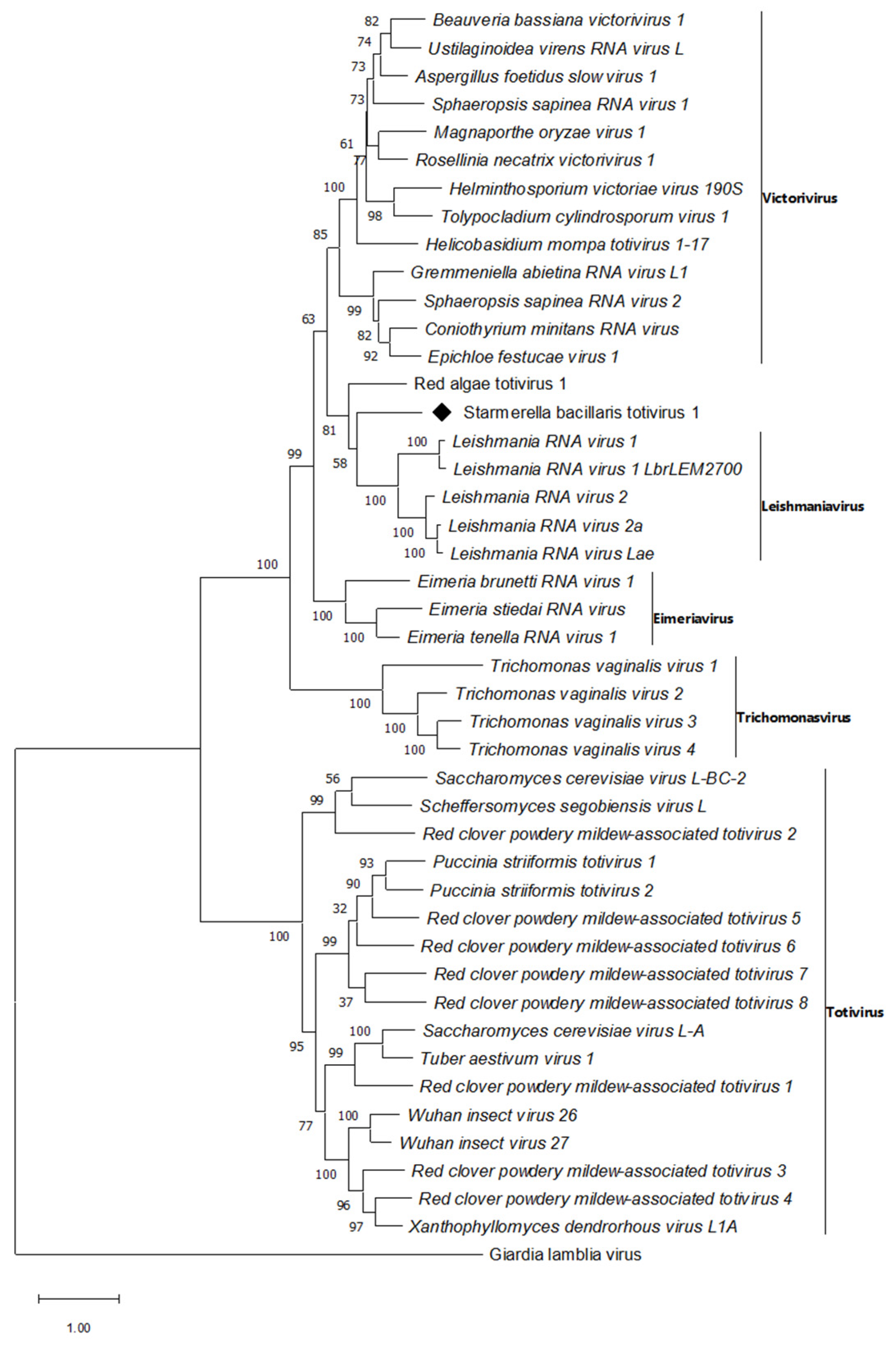
3.1. Totivirus Related Sequences
3.2. Partitivirus Related Sequences
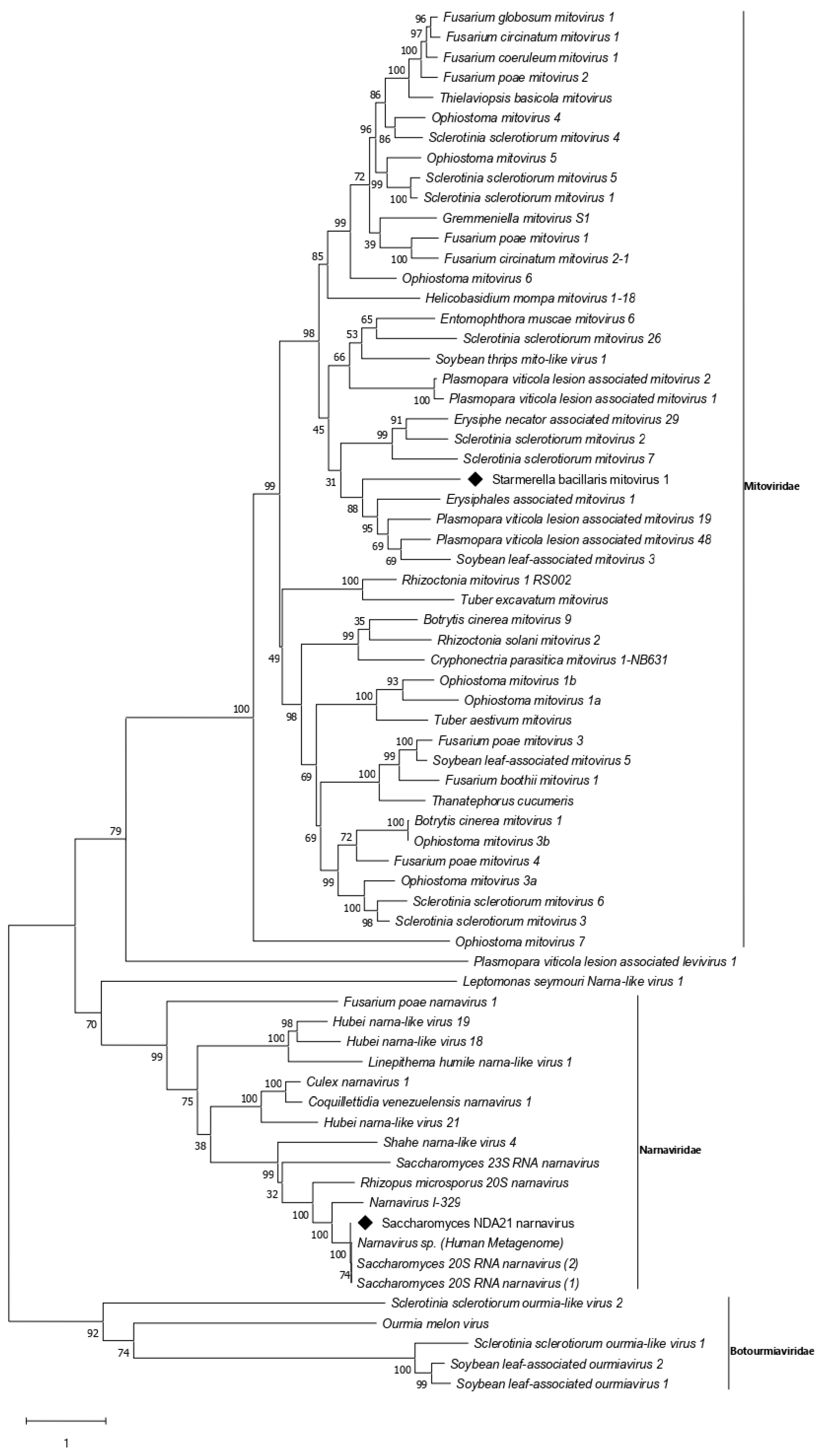
3.3. Narnavirus Related Sequences
3.4. Mitovirus Related Sequences
3.5. ORFan Sequences
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus Years of Fungal Viruses. Virology 2015, 479–480, 356–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, J.M.; Bonds, A.E.; Clemons, R.A.; Thapa, N.A.; Simmons, D.R.; Carter-House, D.; Ortanez, J.; Liu, P.; Miralles-Durán, A.; Desirò, A.; et al. Survey of Early-Diverging Lineages of Fungi Reveals Abundant and Diverse Mycoviruses. mBio 2020, 11, e02027-20. [Google Scholar] [CrossRef]
- Son, M.; Yu, J.; Kim, K.-H. Five Questions about Mycoviruses What Are Mycoviruses? PLoS Pathog. 2015, 11, e1005172. [Google Scholar] [CrossRef] [PubMed]
- Krupovic, M.; Said, A.G.; Jiang, D.; Varsani, A. Genomoviridae: A New Family of Widespread Single-Stranded DNA Viruses. Arch. Virol. 2016, 161, 2633–2643. [Google Scholar] [CrossRef] [Green Version]
- Khalifa, M.E.; Macdiarmid, R.M. A Mechanically Transmitted Dna Mycovirus Is Targeted by the Defence Machinery of Its Host, Botrytis Cinerea. Viruses 2021, 13, 1315. [Google Scholar] [CrossRef]
- Ghabrial, S.A.; Suzuki, N. Viruses of Plant Pathogenic Fungi. Annu. Rev. Phytopathol. 2009, 47, 353–384. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Jiang, D. New Insights into Mycoviruses and Exploration for the Biological Control of Crop Fungal Diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Padilla, A.; Rodríguez-Romero, J.; Gómez-Cid, I.; Pacifico, D.; Ayllón, M.A. Novel Mycoviruses Discovered in the Mycovirome of a Necrotrophic Fungus. mBio 2021, 12, e03705-20. [Google Scholar] [CrossRef]
- Sato, Y.; Castón, J.R.; Suzuki, N. The Biological Attributes, Genome Architecture and Packaging of Diverse Multi-Component Fungal Viruses. Curr. Opin. Virol. 2018, 33, 55–65. [Google Scholar] [CrossRef]
- Jia, H.; Dong, K.; Zhou, L.; Wang, G.; Hong, N.; Jiang, D.; Xu, W. A DsRNA Virus with Filamentous Viral Particles. Nat. Commun. 2017, 8, 168. [Google Scholar] [CrossRef] [Green Version]
- Mu, F.; Li, B.; Cheng, S.I.; Jia, J.; Jiang, D.; Fu, Y.; Cheng, J.; Lin, Y.; Chen, T.; Xie, J. Nine Viruses from Eight Lineages Exhibiting New Evolutionary Modes That Co-Infect a Hypovirulent Phytopathogenic Fungus. PLoS Pathog. 2021, 17, e1009823. [Google Scholar] [CrossRef] [PubMed]
- Chiapello, M.; Rodríguez-Romero, J.; Aylló, M.A.; Turina, M. Analysis of the Virome Associated to Grapevine Downy Mildew Lesions Reveals New Mycovirus Lineages. Virus Evol. 2020, 6, veaa058. [Google Scholar] [CrossRef]
- Picarelli, M.A.S.C.; Forgia, M.; Rivas, E.B.; Nerva, L.; Chiapello, M.; Turina, M.; Colariccio, A. Extreme Diversity of Mycoviruses Present in Isolates of Rhizoctonia Solani AG2-2 LP from Zoysia Japonica from Brazil. Front. Cell. Infect. Microbiol. 2019, 9, 244. [Google Scholar] [CrossRef] [Green Version]
- Crabtree, A.M.; Kizer, E.A.; Hunter, S.S.; van Leuven, J.T.; New, D.D.; Fagnan, M.W.; Rowley, P.A. A Rapid Method for Sequencing Double-Stranded RNAs Purified from Yeasts and the Identification of a Potent K1 Killer Toxin Isolated from Saccharomyces cerevisiae. Viruses 2019, 11, 70. [Google Scholar] [CrossRef] [Green Version]
- Nerva, L.; Ciuffo, M.; Vallino, M.; Margaria, P.; Varese, G.C.; Gnavi, G.; Turina, M. Multiple Approaches for the Detection and Characterization of Viral and Plasmid Symbionts from a Collection of Marine Fungi. Virus Res. 2016, 219, 22–38. [Google Scholar] [CrossRef] [PubMed]
- Abdoulaye, A.H.; Foda, M.F.; Kotta-Loizou, I. Viruses Infecting the Plant Pathogenic Fungus Rhizoctonia Solani. Viruses 2019, 11, 1113. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, K.B.; Holcomb, E.E.; Allscheid, R.L.; Carrington, J.C. Hiding in Plain Sight: New Virus Genomes Discovered via a Systematic Analysis of Fungal Public Transcriptomes. PLoS ONE 2019, 14, e0219207. [Google Scholar] [CrossRef] [Green Version]
- Marzano, S.Y.L.; Domier, L.L. Novel Mycoviruses Discovered from Metatranscriptomics Survey of Soybean Phyllosphere Phytobiomes. Virus Res. 2016, 213, 332–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donaire, L.; Ayllon, M.A. Deep Sequencing of Mycovirus-Derived Small RNAs from Botrytis Species. Mol. Plant Pathol. 2017, 18, 1127–1137. [Google Scholar] [CrossRef] [PubMed]
- Kreuze, J.F.; Perez, A.; Untiveros, M.; Quispe, D.; Fuentes, S.; Barker, I.; Simon, R. Complete Viral Genome Sequence and Discovery of Novel Viruses by Deep Sequencing of Small RNAs: A Generic Method for Diagnosis, Discovery and Sequencing of Viruses. Virology 2009, 388, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Vainio, E.J.; Jurvansuu, J.; Streng, J.; Rajamä, M.-L.; Hantula, J.; Valkonen, J.P.T. Diagnosis and Discovery of Fungal Viruses Using Deep Sequencing of Small RNAs. J. Gen. Virol. 2015, 96, 714–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, D.; Eisenberg, D. Finding Families for Genomic ORFans. Bioinform. Discov. Note 1999, 15, 759–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartholomäus, A.; Wibberg, D.; Winkler, A.; Pühler, A.; Schlüter, A.; Varrelmann, M. Deep Sequencing Analysis Reveals the Mycoviral Diversity of the Virome of an Avirulent Isolate of Rhizoctonia Solani AG-2-2 IV. PLoS ONE 2016, 11, e0165965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andika, I.B.; Wei, S.; Cao, C.; Salaipeth, L.; Kondo, H.; Sun, L. Phytopathogenic Fungus Hosts a Plant Virus: A Naturally Occurring Cross-Kingdom Viral Infection. Proc. Natl. Acad. Sci. USA 2017, 114, 12267–12272. [Google Scholar] [CrossRef] [Green Version]
- Dolja, V.V.; Koonin, E.V. Metagenomics Reshapes the Concepts of RNA Virus Evolution by Revealing Extensive Horizontal Virus Transfer. Virus Res. 2018, 244, 36–52. [Google Scholar] [CrossRef]
- Nerva, L.; Varese, G.C.; Falk, B.W.; Turina, M. Mycoviruses of an Endophytic Fungus Can Replicate in Plant Cells: Evolutionary Implications. Sci. Rep. 2017, 7, 1908. [Google Scholar] [CrossRef] [Green Version]
- Rowley, P.A. The Frenemies within: Viruses, Retrotransposons and Plasmids That Naturally Infect Saccharomyces Yeasts. Yeast 2017, 34, 279–292. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.Y. Yeast for Virus Research. Microb. Cell 2017, 4, 311–330. [Google Scholar] [CrossRef] [Green Version]
- Mardanov, A.V.; Beletsky, A.V.; Tanashchuk, T.N.; Kishkovskaya, S.A.; Ravin, N.V. A Novel Narnavirus from a Saccharomyces cerevisiae Flor Strain. Arch. Virol. 2020, 165, 789–791. [Google Scholar] [CrossRef]
- Rodríguez-Cousiño, N.; Esteban, R. Relationships and Evolution of Double-Stranded RNA Totiviruses of Yeasts Inferred from Analysis of L-A-2 and L-BC Variants in Wine Yeast Strain Populations. Appl. Environ. Microbiol. 2017, 83, e02991-16. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Cousiño, N.; Gómez, P.; Esteban, R. Variation and Distribution of L-A Helper Totiviruses in Saccharomyces Sensu Stricto Yeasts Producing Different Killer Toxins. Toxins 2017, 9, 313. [Google Scholar] [CrossRef]
- Rodríguez-Cousiño, N.; Maqueda, M.; Ambrona, J.; Zamora, E.; Esteban, R.; Ramírez, M. A New Wine Saccharomyces cerevisiae Killer Toxin (Klus), Encoded by a Double-Stranded RNA Virus, with Broad Antifungal Activity Is Evolutionarily Related to a Chromosomal Host Gene. Appl. Environ. Microbiol. 2011, 77, 1822–1832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, M.J.; Breinig, F. Yeast Viral Killer Toxins: Lethality and Self-Protection. Nat. Rev. Microbiol. 2006, 4, 212–221. [Google Scholar] [CrossRef]
- van Vuuren, H.J.J.; Wingfield, B.D. Killer Yeasts-Cause of Stuck Fermentations in a Wine Cellar. S. Afr. J. Enol. Vitic. 1986, 7, 113–118. [Google Scholar] [CrossRef] [Green Version]
- Bussey, H.; Sacks, W.; Galley, D.; Saville, D. Yeast Killer Plasmid Mutations Affecting Toxin Secretion and Activity and Toxin Immunity Function. Mol. Cell. Biol. 1982, 2, 346–354. [Google Scholar] [PubMed] [Green Version]
- Boynton, P.J. The Ecology of Killer Yeasts: Interference Competition in Natural Habitats. Yeast 2019, 36, 473–485. [Google Scholar] [CrossRef]
- Fredericks, L.R.; Lee, M.D.; Crabtree, A.M.; Boyer, J.M.; Kizer, E.A.; Taggart, N.T.; Roslund, C.R.; Hunter, S.S.; Kennedy, C.B.; Willmore, C.G.; et al. The Species-Specific Acquisition and Diversification of a K1-like Family of Killer Toxins in Budding Yeasts of the Saccharomycotina. PLoS Genet. 2021, 17, e1009341. [Google Scholar] [CrossRef] [PubMed]
- Melvydas, V.; Bružauskaitė, I.; Gedminienė, G.; Šiekštelė, R. A Novel Saccharomyces cerevisiae Killer Strain Secreting the X Factor Related to Killer Activity and Inhibition of S. cerevisiae K1, K2 and K28 Killer Toxins. Indian J. Microbiol. 2016, 56, 335–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csoma, H.; Zakany, N.; Capece, A.; Romano, P.; Sipiczki, M. Biological Diversity of Saccharomyces Yeasts of Spontaneously Fermenting Wines in Four Wine Regions: Comparative Genotypic and Phenotypic Analysis. Int. J. Food Microbiol. 2010, 140, 239–248. [Google Scholar] [CrossRef]
- Ivannikova, Y.V.; Naumova, E.S.; Naumov, G.I. Viral DsRNA in the Wine Yeast Saccharomyces Bayanus Var. Uvarum. Res. Microbiol. 2007, 158, 638–643. [Google Scholar] [CrossRef]
- Kadowaki, K.; Halvorson, H.O. Appearance of a New Species of Ribonucleic Acid During Sporulation in Saccharomyces cerevisiae. J. Bacteriol. 1971, 105, 826–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillman, B.I.; Esteban, R. Family Narnaviridae. In Virus Taxonomy: Ninth Report of the International Committee for the Taxonomy of Viruses; King, A., Adams, M., Carstens, E., Lefkowitz, E., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2011; pp. 1025–1030. [Google Scholar]
- Vepštaitė-Monstavičė, I.; Lukša, J.; Konovalovas, A.; Ežerskytė, D.; Stanevičienė, R.; Strazdaitė-žielienė, Ž.; Serva, S.; Servienė, E. Saccharomyces Paradoxus K66 Killer System Evidences Expanded Assortment of Helper and Satellite Viruses. Viruses 2018, 10, 564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmiti, M.J.; Neuhausen, F. Killer Toxin-Secreting Double-Stranded RNA Mycoviruses in the Yeasts Hanseniaspora Uvarum and Zygosaccharomyces Bailii. J. Virol. 1994, 68, 1765–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez, M.; Velázquez, R.; Maqueda, M.; Martínez, A. Genome Organization of a New Double-Stranded RNA LA Helper Virus From Wine Torulaspora Delbrueckii Killer Yeast as Compared With Its Saccharomyces Counterparts. Front. Microbiol. 2020, 11, 593846. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.D.; Pogany, J.; Lin, J.Y. How Yeast Can Be Used as a Genetic Platform to Explore Virus–Host Interactions: From ‘Omics’ to Functional Studies. Trends Microbiol. 2014, 22, 309–316. [Google Scholar] [CrossRef]
- Sahaya Glingston, R.; Yadav, J.; Rajpoot, J.; Joshi, N.; Nagotu, S. Contribution of Yeast Models to Virus Research. Appl. Microbiol. Biotechnol. 2021, 105, 4855–4878. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, E.E.; Coletta-Filho, H.D.; Nourin, S.; Falk, B.W.; Nerva, L.; Oliveira, T.S.; Dorta, S.O.; Machado, M.A. Deep Sequencing Analysis of RNAs from Citrus Plants Grown in a Citrus Sudden Death–Affected Area Reveals Diverse Known and Putative Novel Viruses. Viruses 2017, 9, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nerva, L.; Varese, G.C.; Turina, M. Different Approaches to Discover Mycovirus Associated to Marine Organisms. In Methods in Molecular Biology; Humana Press, Ed.; Humana Press: New York, NY, USA, 2018; Volume 1746. [Google Scholar]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De Novo Transcript Sequence Reconstruction from RNA-Seq Using the Trinity Platform for Reference Generation and Analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A New Generation of Protein Database Search Programs; Oxford University Press: Oxford, UK, 1997; Volume 25. [Google Scholar]
- Langmead, B.; Wilks, C.; Antonescu, V.; Charles, R. Scaling Read Aligners to Hundreds of Threads on General-Purpose Processors. Bioinformatics 2019, 35, 421–432. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows-Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-Performance Genomics Data Visualization and Exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. TrimAl: A Tool for Automated Alignment Trimming in Large-Scale Phylogenetic Analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Mizutani, Y.; Abraham, A.; Uesaka, K.; Kondo, H.; Suga, H.; Suzuki, N.; Chiba, S. Novel Mitoviruses and a Unique Tymo-like Virus in Hypovirulent and Virulent Strains of the Fusarium Head Blight Fungus, Fusarium Boothii. Viruses 2018, 10, 584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not Your Ordinary Yeast: Non-Saccharomyces Yeasts in Wine Production Uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steensels, J.; Meersman, E.; Snoek, T.; Saels, V.; Verstrepen, K.J. Large-Scale Selection and Breeding to Generate Industrial Yeasts with Superior Aroma Production. Appl. Environ. Microbiol. 2014, 80, 6965–6975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comitini, F.; Agarbati, A.; Canonico, L.; Ciani, M. Yeast Interactions and Molecular Mechanisms in Wine Fermentation: A Comprehensive Review. Int. J. Mol. Sci. 2021, 22, 7754. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and Future of Non-Saccharomyces Yeasts: From Spoilage Microorganisms to Biotechnological Tools for Improving Wine Aroma Complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef] [Green Version]
- Díaz, M.A.; Pereyra, M.M.; Picón-Montenegro, E.; Meinhardt, F.; Dib, J.R. Killer Yeasts for the Biological Control of Postharvest Fungal Crop Diseases. Microorganisms 2020, 8, 1680. [Google Scholar] [CrossRef]
- Muccilli, S.; Restuccia, C. Bioprotective Role of Yeasts. Microorganisms 2015, 3, 588–611. [Google Scholar] [CrossRef] [Green Version]
- Drinnenberg, I.A.; Fink, G.R.; Bartel, D.P. Compatibility with Killer Explains the Rise of RNAi-Deficient Fungi. Science 2011, 333, 1592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.L.; Leu, J.Y.; Chang, T.H. A Population Study of Killer Viruses Reveals Different Evolutionary Histories of Two Closely Related Saccharomyces Sensu Stricto Yeasts. Mol. Ecol. 2015, 24, 4312–4322. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, M.; Velázquez, R.; López-Piñeiro, A.; Naranjo, B.; Roig, F.; Llorens, C. New Insights into the Genome Organization of Yeast Killer Viruses Based on “Atypical” Killer Strains Characterized by High-Throughput Sequencing. Toxins 2017, 9, 292. [Google Scholar] [CrossRef] [PubMed]
- Mackay, I.M.; Arden, K.E.; Nitsche, A. survey and summary Real-Time PCR in Virology. Nucleic Acids Res. 2002, 30, 1292–1305. [Google Scholar] [CrossRef] [Green Version]
- Chiba, Y.; Tomaru, Y.; Shimabukuro, H.; Kimura, K.; Hirai, M.; Takaki, Y.; Hagiwara, D.; Nunoura, T.; Urayama, S.I. Viral Rna Genomes Identified from Marine Macroalgae and a Diatom. Microbes Environ. 2020, 35, ME20016. [Google Scholar] [CrossRef]
- Vainio, E.J.; Chiba, S.; Ghabrial, S.A.; Maiss, E.; Roossinck, M.; Sabanadzovic, S.; Suzuki, N.; Xie, J.; Nibert, M. ICTV Virus Taxonomy Profile: Partitiviridae. J. Gen. Virol. 2018, 99, 17–18. [Google Scholar] [CrossRef]
- Aitmanaitė, L.; Konovalovas, A.; Medvedevas, P.; Servienė, E.; Serva, S. Specificity Determination in Saccharomyces Cerevisiae Killer Virus Systems. Microorganisms 2021, 9, 236. [Google Scholar] [CrossRef] [PubMed]
- Lukša, J.; Ravoitytė, B.; Aitmanaitė, A.; Lina, K.; Butenko, A.; Yurchenko, V.; Serva, S.; Servienė, E. Different Metabolic Pathways Are Involved in Response of Saccharomyces cerevisiae to L-A and M Viruses. Toxins 2017, 9, 233. [Google Scholar] [CrossRef]
- Dignard, D.; Whiteway, M.; Germain, D.; Tessier, D.; Thomas, D.Y. Expression in Yeast of a EDNA Copy of the K2 Killer Toxin Gene. Mol. Genet. Genom. 1991, 227, 127–136. [Google Scholar] [CrossRef]
- Lopez, V.; Gil, R.; Carbonell, J.V.; Navarro, A. Occurrence of 20S RNA and 23S RNA Replicons in Industrial Yeast Strains and Their Variation under Nutritional Stress Conditions. Yeast 2002, 19, 545–552. [Google Scholar] [CrossRef] [Green Version]
- Esteban, R.; Vega, L.; Fujimura, T. Launching of the Yeast 20 S RNA Narnavirus by Expressing the Genomic or Antigenomic Viral RNA in Vivo. J. Biol. Chem. 2005, 280, 33725–33734. [Google Scholar] [CrossRef] [Green Version]
- Nibert, M.L.; Ghabrial, S.A.; Maiss, E.; Lesker, T.; Vainio, E.J.; Jiang, D.; Suzuki, N. Taxonomic Reorganization of Family Partitiviridae and Other Recent Progress in Partitivirus Research. Virus Res. 2014, 188, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Englezos, V.; Giacosa, S.; Rantsiou, K.; Rolle, L.; Cocolin, L. Starmerella Bacillaris in Winemaking: Opportunities and Risks. Curr. Opin. Food Sci. 2017, 17, 30–35. [Google Scholar] [CrossRef]
- Romancino, D.P.; di Maio, S.; Muriella, R.; Oliva, D. Analysis of Non-Saccharomyces Yeast Populations Isolated from Grape Musts from Sicily (Italy). J. Appl. Microbiol. 2008, 105, 2248–2254. [Google Scholar] [CrossRef] [PubMed]
- di Maio, S.; Genna, G.; Gandolfo, V.; Amore, G.; Ciaccio, M.; Oliva, D. Presence of Candida Zemplinina in Sicilian Musts and Selection of a Strain for Wine Mixed Fermentations. S. Afr. J. Enol. Vitic. 2012, 33, 80–87. [Google Scholar] [CrossRef]
- Giaramida, P.; Ponticello, G.; di Maio1, S.; Squadrito, M.; Genna, G.; Barone, E.; Scacco, A.; Corona, O.; Amore, G.; di Stefano, R.; et al. Candida Zemplinina for Production of Wines with Less Alcohol and More Glycerol. S. Afr. J. Enol. Vitic. 2013, 34, 204–211. [Google Scholar] [CrossRef] [Green Version]
- Jamal, A.; Sato, Y.; Shahi, S.; Shamsi, W.; Kondo, H.; Suzuki, N. Novel Victorivirus from a Pakistani Isolate of Alternaria Alternata Lacking a Typical Translational Stop/Restart Sequence Signature. Viruses 2019, 11, 577. [Google Scholar] [CrossRef] [Green Version]
- Wickner, R.B.; Ghabrial, S.A.; Nibert, M.L.; Patterson, J.L.; Wang, C.C. Totiviridae. In Virus Taxonomy: Classification and Nomenclature of Viruses: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier: San Diego, CA, USA, 2012; pp. 639–650. [Google Scholar]
- Gómez-Arreaza, A.; Haenni, A.L.; Dunia, I.; Avilán, L. Viruses of Parasites as Actors in the Parasite-Host Relationship: A “Ménage à Trois. ” Acta Trop. 2017, 166, 126–132. [Google Scholar] [CrossRef]
- Roossinck, M.J. The Good Viruses: Viral Mutualistic Symbioses. Nat. Rev. Microbiol. 2011, 9, 99–108. [Google Scholar] [CrossRef]
- Ives, A.; Ronet, C.; Prevel, F.; Ruzzante, G.; Fuertes-Marraco, S.; Schutz, F.; Zangger, H.; Revaz-Breton, M.; Lye, L.F.; Hickerson, S.M.; et al. Leishmania RNA Virus Controls the Severity of Mucocutaneous Leishmaniasis. Science 2011, 331, 775–778. [Google Scholar] [CrossRef] [Green Version]
- Fichorova, R.N.; Lee, Y.; Yamamoto, H.S.; Takagi, Y.; Hayes, G.R.; Goodman, R.P.; Chepa-Lotrea, X.; Buck, O.R.; Murray, R.; Kula, T.; et al. Endobiont Viruses Sensed by the Human Host-Beyond Conventional Antiparasitic Therapy. PLoS ONE 2012, 7, e48418. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, M.C.; Higgins, J.; Abrahante, J.E.; Kniel, K.E.; O’Brien, C.; Trout, J.; Lancto, C.A.; Abrahamsen, M.S.; Fayer, R. Fecundity of Cryptosporidium Parvum Is Correlated with Intracellular Levels of the Viral Symbiont CPV. Int. J. Parasitol. 2008, 38, 1051–1055. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.L.; Wang, A.L.; Wang, C.C. Purification and Characterization of the Giardia Lamblia Double-Stranded RNA Virus. Mol. Biochem. Parasitol. 1988, 28, 189–195. [Google Scholar] [CrossRef]
- Syller, J. Facilitative and Antagonistic Interactions between Plant Viruses in Mixed Infections. Mol. Plant Pathol. 2012, 13, 204–216. [Google Scholar] [CrossRef] [PubMed]
- Dobbs, E.; Deakin, G.; Bennett, J.; Fleming-Archibald, C.; Jones, I.; Grogan, H.; Burton, K.; Dobbs, C.E. Viral Interactions and Pathogenesis during Multiple Viral Infections in Agaricus Bisporus. mBio 2021, 12, e03470-20. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Source | Country | Killer Phenotype | Brand | |
|---|---|---|---|---|---|---|
| 1 | Saccharomyces cerevisiae | Lalvin BA11 | Grape must | Portugal | S | Lallemand |
| 2 | Saccharomyces cerevisiae | Lalvin ICV D254 | Grape must | France | N | Lallemand |
| 3 | Saccharomyces cerevisiae | Lalvin Rhone L2226 | Grape must | France | K | Lallemand |
| 4 | Saccharomyces cerevisiae | Lalvin EC1118 | Grape must | France | K | Lallemand |
| 5 | Saccharomyces cerevisiae | Lalvin RC212 | Grape must | France | S | Lallemand |
| 6 | Saccharomyces cerevisiae | Lalvin Rhone 2323 | Grape must | France | K | Lallemand |
| 7 | Saccharomyces cerevisiae | Lalvin QA23 | Grape must | Portugal | K | Lallemand |
| 8 | Saccharomyces cerevisiae | Uvaferm 43 | Grape must | France | N | Lallemand |
| 9 | Saccharomyces cerevisiae | Zymaflore VL1 | Grape must | France | N | Laffort |
| 10 | Saccharomyces cerevisiae | Zymaflore F15 | Grape must | France | S | Laffort |
| 11 | Saccharomyces cerevisiae | Zymaflore X5 | Breeding | n.a. | K | Laffort |
| 12 | Saccharomyces cerevisiae | Zymaflore FX10 | Breeding | n.a. | N | Laffort |
| 13 | Saccharomyces cerevisiae | Fermicru VR5 | Grape must | France | N | Oenobrands |
| 14 | Saccharomyces cerevisiae | PDM | Grape must | France | K | Maurivin |
| 15 | Saccharomyces cerevisiae | EZFERM44 | Grape must | n.a. | N | Enartis |
| 16 | Saccharomyces cerevisiae | IRVO NDA21 | Grape must | Italy | n.a. | Fermentis |
| 17 | Metschnikowia pulcherrima | IRVO Non-Sac2 | Grape must | Italy | n.a. | n.a. |
| 18 | Hanseniaspora uvarum | IRVO Non-Sac7 | Grape must | Italy | n.a. | n.a. |
| 19 | Kluyveromyces marxianus | IRVO Non-Sac10 | Human feces | Italy | n.a. | n.a. |
| 20 | Starmerella bacillaris (sin. Candida zemplinina) | IRVO Cz12 | Grape must | Italy | n.a. | n.a. |
| 21 | Lachancea (Kluyveromyces) thermotolerans | DBVPG 6232 | Plum conserve | USSR | n.a. | n.a. |
| 22 | Hanseniaspora guilliermondii | DBVPG 6796 | Infected nail | South Africa | n.a. | n.a. |
| 23 | Candida stellata | DBVPG 6714 | Grapevine | Germany | n.a | n.a. |
| 24 | Dekkera anomala | DBVPG 4075 | Beer | UK | n.a. | n.a. |
| Genus | Isolate | Isolate Abbreviation | GenBank Accession No. |
|---|---|---|---|
| Totivirus | Saccharomyces cerevisiae virus L-A-lus strain D254 | ScV-LA-lus-D254 | OK377006 |
| Saccharomyces cerevisiae virus L-A-2 strain L2226 | ScV-LA2-L2226 | OK377007 | |
| Saccharomyces cerevisiae virus L-A-2 strain EC1118 | ScV-LA2-EC1118 | OK377008 | |
| Saccharomyces cerevisiae virus L-A-2 strain 2323 | ScV-LA2-2323 | OK377009 | |
| Saccharomyces cerevisiae virus L-A-2 strain QA23 | ScV-LA2-QA23 | OK377010 | |
| Saccharomyces cerevisiae virus L-A-lus strain U43 | ScV-LA-lus-U43 | OK377011 | |
| Saccharomyces cerevisiae virus L-A-2 strain VL1 | ScV-LA2-VL1 | OK377012 | |
| Saccharomyces cerevisiae virus L-A-2 strain X5 | ScV-LA2-X5 | OK377013 | |
| Saccharomyces cerevisiae virus L-A-2 strain FX10 | ScV-LA2-FX10 | OK377014 | |
| Saccharomyces cerevisiae virus L-A-2 strain VR5 | ScV-LA2-VR5 | OK377015 | |
| Saccharomyces cerevisiae virus L-A-2 strain PDM | ScV-LA2-PDM | OK377016 | |
| Saccharomyces cerevisiae virus L-A-2 strain EZ44 | ScV-LA2-EZ44 | OK377017 | |
| Saccharomyces cerevisiae virus L-BC-2 strain EZ44 | ScV-LBC2-EZ44 | OK377025 | |
| Saccharomyces cerevisiae virus L-BC-2 strain PDM | ScV-LBC2-PDM | OK377024 | |
| Saccharomyces cerevisiae virus L-BC-2 strain VR5 | ScV-LBC2-VR5 | OK377023 | |
| Saccharomyces cerevisiae virus L-BC-2 strain X5 | ScV-LBC2-X5 | OK377022 | |
| Saccharomyces cerevisiae virus L-BC-2 strain QA23 | ScV-LBC2-QA23 | OK377021 | |
| Saccharomyces cerevisiae virus L-BC (La) strain RC212 | ScV-LBC-La-RC212 | OK377020 | |
| Saccharomyces cerevisiae virus L-BC-2 strain EC1118 | ScV-LBC2-EC1118 | OK377019 | |
| Saccharomyces cerevisiae virus L-BC (La) strain D254 | ScV-LBC-La-D254 | OK377018 | |
| Saccharomyces cerevisiae virus satellite dsRNA Mlus strain D254 | ScV-Mlus-D254 | OK412909 | |
| Saccharomyces cerevisiae virus satellite dsRNA M2 strain L2226 | ScV-M2-L2226 | OK412898 | |
| Saccharomyces cerevisiae virus satellite dsRNA M2 strain EC1118 | ScV-M2-EC1118 | OK412899 | |
| Saccharomyces cerevisiae virus satellite dsRNA M2 strain 2323 | ScV-M2-2323 | OK412900 | |
| Saccharomyces cerevisiae virus satellite dsRNA M2 strain QA23 | ScV-M2-QA23 | OK412901 | |
| Saccharomyces cerevisiae virus satellite dsRNA M2 strain U43 | ScV-M2-U43 | OK412902 | |
| Saccharomyces cerevisiae virus satellite dsRNA M2 strain VL1 | ScV-M2-VL1 | OK412903 | |
| Saccharomyces cerevisiae virus satellite dsRNA M2 strain X5 | ScV-M2-X5 | OK412904 | |
| Saccharomyces cerevisiae virus satellite dsRNA M2 strain FX10 | ScV-M2-FX10 | OK412905 | |
| Saccharomyces cerevisiae virus satellite dsRNA M2 strain VR5 | ScV-M2-VR5 | OK412906 | |
| Saccharomyces cerevisiae virus satellite dsRNA M2 strain PDM | ScV-M2-PDM | OK412907 | |
| Saccharomyces cerevisiae virus satellite dsRNA M2 strain EZ44 | ScV-M2-EZ44 | OK412908 | |
| Starmerella bacillaris totivirus 1 * | SbTV1 * | OK412911 | |
| Cryspovirus | Saccharomyces cerevisiae cryspovirus 1 -D254 (RNA2) * | ScCV1-D254 | OK412913 |
| Saccharomyces cerevisiae cryspovirus 1 -U43 (RNA2) * | ScCV1-U43 | OK412914 | |
| Saccharomyces cerevisiae cryspovirus 1 -D254 (RNA1) * | ScCV1-D254 | OK412915 | |
| Saccharomyces cerevisiae cryspovirus 1 -U43 (RNA1) * | ScCV1-U43 | OK412916 | |
| Narnavirus | Saccharomyces NDA21 narnavirus | ScNV-NDA21 | OK412912 |
| Mitovirus | Starmerella bacillaris mitovirus 1 * | SbMV1 * | OK412910 |
| Totiviridae | Partitiviridae | Narnaviridae | Mitoviridae | |||||
|---|---|---|---|---|---|---|---|---|
| Strain | ScV-L-A | ScV-M2 | ScV-Mlus | ScV-L-BC | SbTV1 | ScCV1 | ScNV-20S | SbMV1 |
| Lalvin ICV D254 | ScV-LA-lus-D254 (20.25) | ScV-Mlus-D254 (19.33) | ScV-LBC-La-D254 (26.86) | ScCV1-D254 RNA1 (19.58) RNA2, (20.13) | ||||
| Lalvin Rhone L2226 | ScV-LA2-L2226 (23.17) | ScV-M2-L2226(21.04) | ||||||
| Lalvin EC1118 | ScV-LA2-EC118 (22.34) | ScV-M2-EC118 (20.44) | ScV-LBC2-EC1118 (26.37) | |||||
| Lalvin RC212 | ScV-LBC-La-RC212 (26.22) | |||||||
| Lalvin Rhone 2323 | ScV-LA2-2323(23.05) | ScV-M2-2323 (19.05) | ||||||
| Lalvin QA23 | ScV-LA2-QA23 (22.33) | ScV-M2-QA23(18.23) | ScV-LBC2-QA23 (26.91) | |||||
| Uvaferm 43 | ScV-LA-lus-U43 (24.28) | ScV-M2-U43 (18.18) | ScCV1-U43 (RNA1, 20.71) (RNA2, 16.30) | |||||
| Zymaflore VL1 | ScV-LA2-VL1 (22.78) | ScV-M2-VL1 (18.61) | ||||||
| Zymaflore X5 | ScV-LA2-X5 (23.61) | ScV-M2-X5 (21.68) | ScV-LBC2-X5(27.08) | |||||
| Zymaflore FX10 | ScV-LA2-FX10(22.51) | ScV-M2-FX10(21.33) | ||||||
| Fermicru VR5 | ScV-LA2-VR5 (22.72) | ScV-M2-VR5 (21.47) | ScV-LBC2-VR5 (27.72) | |||||
| PDM | ScV-LA2-PDM (23.50) | ScV-M2-PDM (21.78) | ScV-LBC2-PDM (28.30) | |||||
| EZFERM44 | ScV-LA2-EZ44(24.38) | ScV-M2-EZ44 (20.56) | ScV-LBC2-EZ44 (28.01) | |||||
| NDA21 | ScNV-20S- NDA21 (18.00) | |||||||
| Cz12 | SbTV1-Cz12 (20.35) | SbMV1-Cz12 (22.92) | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crucitti, D.; Chiapello, M.; Oliva, D.; Forgia, M.; Turina, M.; Carimi, F.; La Bella, F.; Pacifico, D. Identification and Molecular Characterization of Novel Mycoviruses in Saccharomyces and Non-Saccharomyces Yeasts of Oenological Interest. Viruses 2022, 14, 52. https://doi.org/10.3390/v14010052
Crucitti D, Chiapello M, Oliva D, Forgia M, Turina M, Carimi F, La Bella F, Pacifico D. Identification and Molecular Characterization of Novel Mycoviruses in Saccharomyces and Non-Saccharomyces Yeasts of Oenological Interest. Viruses. 2022; 14(1):52. https://doi.org/10.3390/v14010052
Chicago/Turabian StyleCrucitti, Dalila, Marco Chiapello, Daniele Oliva, Marco Forgia, Massimo Turina, Francesco Carimi, Francesca La Bella, and Davide Pacifico. 2022. "Identification and Molecular Characterization of Novel Mycoviruses in Saccharomyces and Non-Saccharomyces Yeasts of Oenological Interest" Viruses 14, no. 1: 52. https://doi.org/10.3390/v14010052