
Efficacy of Salmonella Bacteriophage S1 Delivered and Released by Alginate Beads in a Chicken Model of Infection

 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Method
2.1. Bacterial Strains
2.2. Phage S1 Isolation
2.3. Enumeration of Phages
2.4. Sequencing and Annotation of Phage S1
2.5. Efficiency of Plating of Phage S1
2.6. Encapsulation of Phage S1 in ALG Beads
2.7. Determination of the Encapsulation Efficiency
2.8. Comparison of SE PT13A Growth under the Exposure of ALG and ALG + Phage
2.9. In Vitro Protection and Telease of Free Phages and Encapsulated Phages in ALG Beads
2.10. In Vivo Assays
2.10.1. Experimental Animals
2.10.2. Degradation of the ALG Beads and Phage S1 Release
2.10.3. Elimination of SE PT13A in Chickens
3. Results and Discussion
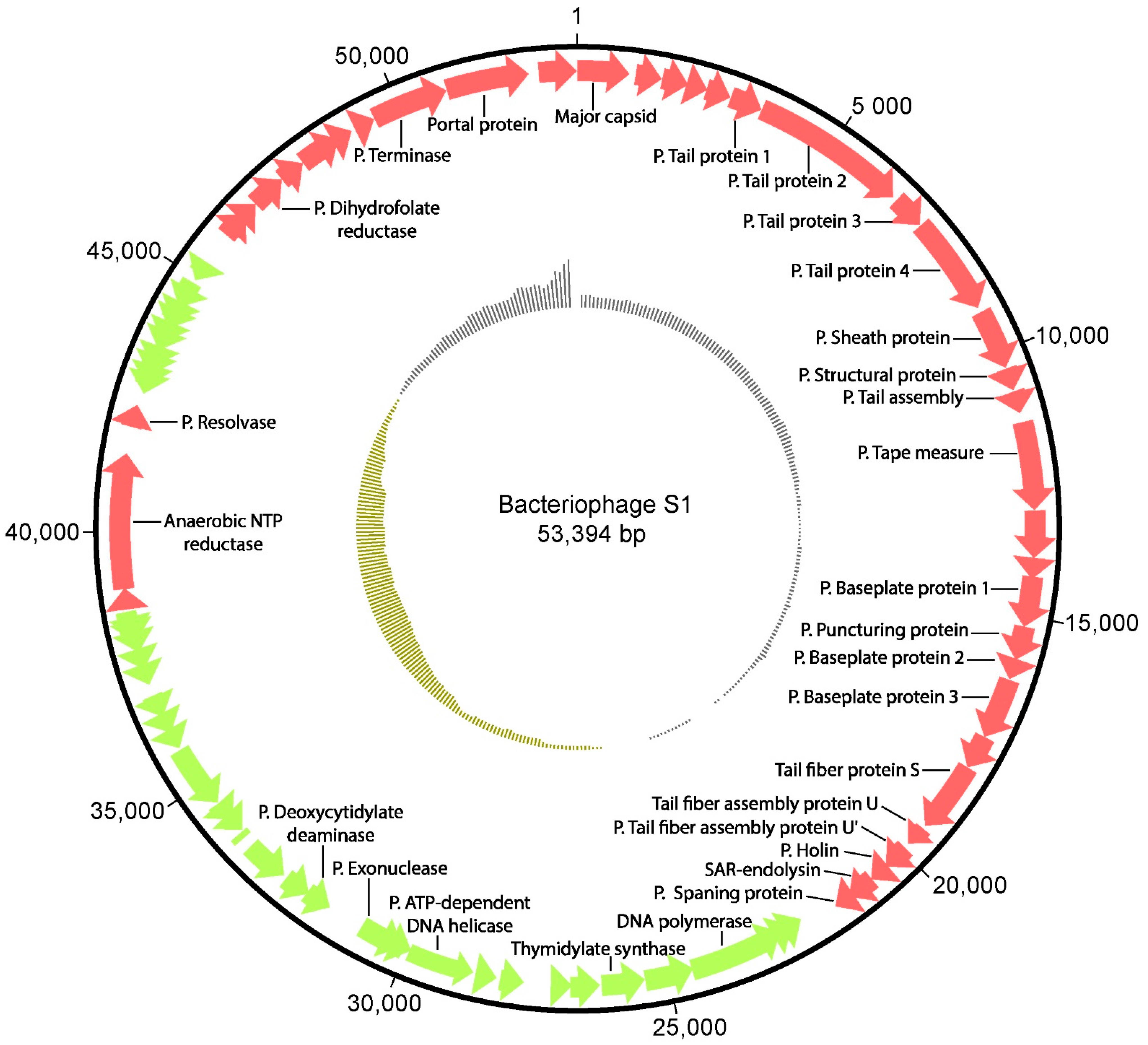
3.1. Sequencing of Phage S1
3.2. Host Range by EOP
3.3. Particle Size and Encapsulation Efficiency
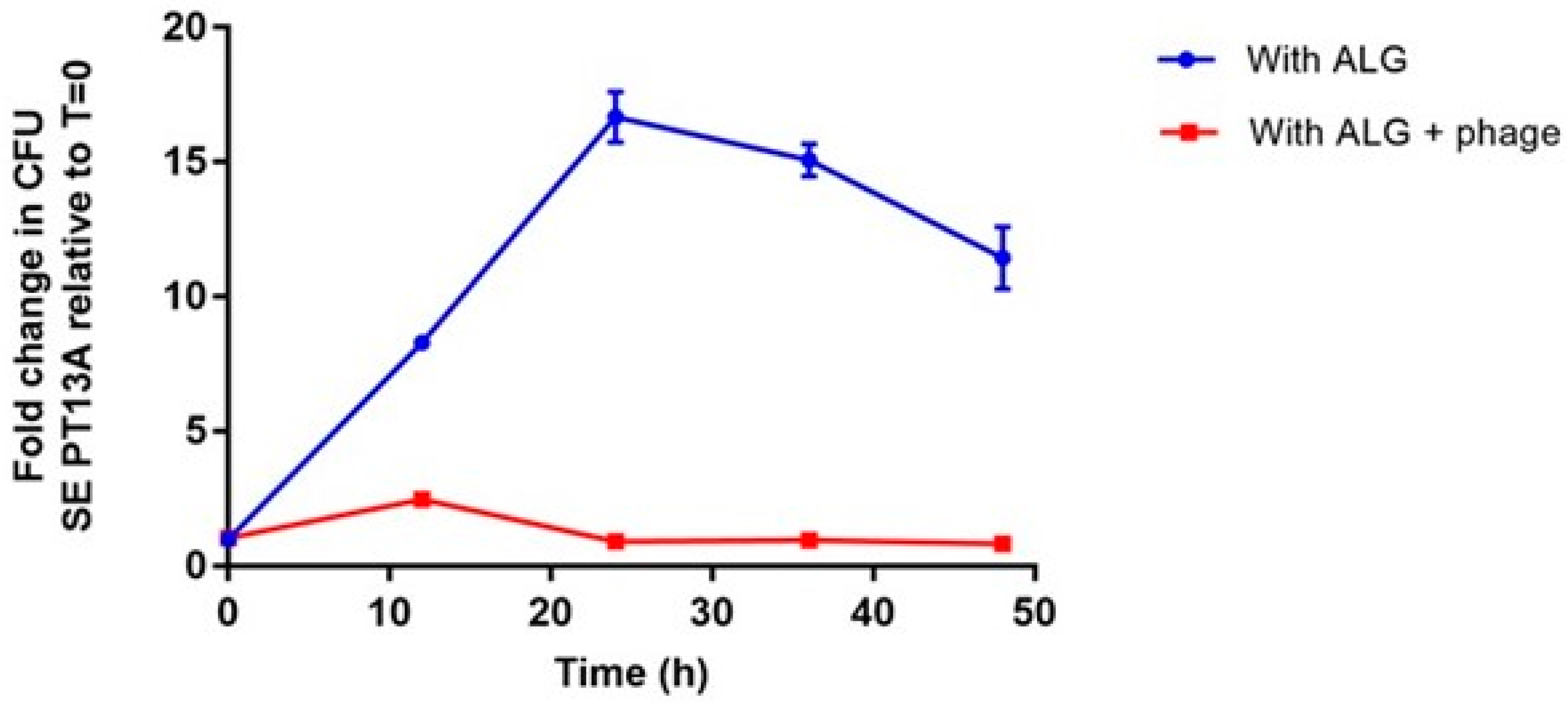
3.4. Comparison of SE PT13A Growth under the Exposure of ALG and ALG + Phage
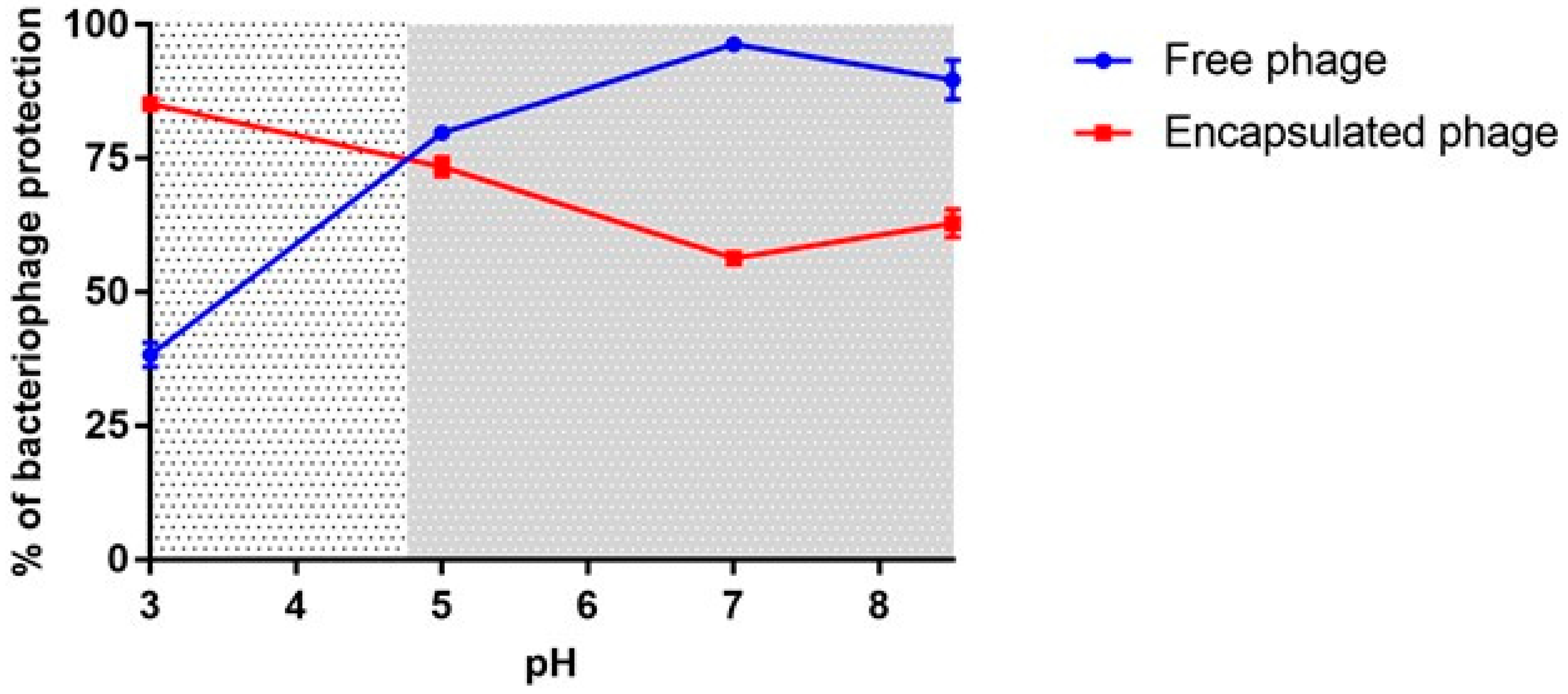
3.5. In Vitro Protection and Release of Free Phages and Encapsulated Phages in ALG Beads
3.6. In Vivo Assays
3.6.1. Degradation of the ALG Beads and Phage S1 Release
3.6.2. Elimination of SE PT13A in Chickens
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xiong, D.; Song, L.; Pan, Z.; Jiao, X. Identification and Discrimination of Salmonella enterica Serovar Gallinarum Biovars Pullorum and Gallinarum Based on a One-Step Multiplex PCR Assay. Front. Microbiol. 2018, 9, 1718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berhanu, G.; Fulas, A. Pullorum Disease and Fowl Typhoid in Poultry: A Review. Br. J. Poult. Sci. 2020, 9, 48–56. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Salmonella (Non-Typhoidal). 2018. Available online: https://www.who.int/news-room/fact-sheets/detail/salmonella-(non-typhoidal) (accessed on 1 January 2021).
- Desin, T.S.; Lam, P.K.; Koch, B.; Mickael, C.; Berberov, E.; Wisner, A.L.; Townsend, H.G.; Potter, A.A.; Köster, W. Salmonella enterica serovar enteritidis pathogenicity island 1 is not essential for but facilitates rapid systemic spread in chickens. Infect. Immun. 2009, 77, 2866–2875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ardoino, S.M.; Toso, R.E.; Álvarez, H.L.; Mariani, E.L.; Cachau, P.D.; Mancilla, M.V.; Oriani, D.S. Antimicrobianos como promotores de crecimiento (AGP) en alimentos balanceados para aves: Uso, resistencia bacteriana, nuevas alternativas y opciones de reemplazo. Cienc. Vet. 2017, 19, 50–66. [Google Scholar] [CrossRef]
- Lozano, A.M.C.; Arias, M.D.C. Residuos de fármacos en alimentos de origen animal: Panorama actual en Colombia. Rev. Colomb. Cienc. Pecu. 2008, 21, 121–135. [Google Scholar]
- Gonzáles, A.S.; Icochea, D.E.; Reyna, S.P.; Guzmán, G.J.; Cazorla, M.F.; Lúcar, J.; Carcelén, C.F.; San Martín, V. Effect of the supplementation of organic acids on productive parameters in broilers. Rev. Investig. Vet. Perú (RIVEP) 2013, 24, 32–37. [Google Scholar]
- Górski, A.; Jończyk-Matysiak, E.; Miedzybrodzki, R.; Weber-Dabrowska, B.; Łusiak-Szelachowska, M.; Bagińska, N.; Borysowski, J.; Łobocka, M.B.; Wegrzyn, A.; Wȩgrzyn, G. Phage therapy: Beyond antibacterial action. Front. Med. 2018, 5, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazi, M.; Annapure, U.S. Bacteriophage biocontrol of foodborne pathogens. J. Food Sci. Technol. 2016, 53, 1355–1362. [Google Scholar] [CrossRef] [Green Version]
- Berghaus, R.D.; Thayer, S.G.; Law, B.F.; Mild, R.M.; Hofacre, C.L.; Singer, R.S. Enumeration of Salmonella and Campylobacter spp. in environmental farm samples and processing plant carcass rinses from commercial broiler chicken flocks. Appl. Environ. Microbiol. 2013, 79, 4106–4114. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Whitehouse, C.A.; Li, B. Presence and Persistence of Salmonella in Water: The Impact on Microbial Quality of Water and Food Safety. Front. Public Health 2018, 6, 159. [Google Scholar] [CrossRef]
- Choińska-Pulit, A.; Mituła, P.; Śliwka, P.; Łaba, W.; Skaradzińska, A. Bacteriophage encapsulation: Trends and potential applications. Trends Food Sci. Technol. 2015, 45, 212–221. [Google Scholar] [CrossRef]
- Wernicki, A.; Nowaczek, A.; Urban-Chmiel, R. Bacteriophage therapy to combat bacterial infections in poultry. Virol. J. 2017, 14, 179. [Google Scholar] [CrossRef]
- Kim, K.H.; Lee, G.Y.; Jang, J.C.; Kim, J.E.; Kim, Y.Y. Evaluation of Anti-SE Bacteriophage as Feed Additives to Prevent Salmonella enteritidis (SE) in Broiler. Asian Australas. J. Anim. Sci. 2013, 26, 386–393. [Google Scholar] [CrossRef] [Green Version]
- Zbikowska, K.; Michalczuk, M.; Dolka, B. The use of bacteriophages in the poultry industry. Animals 2020, 10, 872. [Google Scholar] [CrossRef]
- Fiorentin, L.; Vieira, N.D.; Barioni, W., Jr. Oral treatment with bacteriophages reduces the concentration of Salmonella Enteritidis PT4 in caecal contents of broilers. Avian Pathol. 2005, 34, 258–263. [Google Scholar] [CrossRef]
- Andreatti Filho, R.L.; Higgins, J.P.; Higgins, S.E.; Gaona, G.; Wolfenden, A.D.; Tellez, G.; Hargis, B.M. Ability of bacteriophages isolated from different sources to reduce Salmonella enterica serovar enteritidis in vitro and in vivo. Poul. Sci. 2007, 86, 1904–1909. [Google Scholar] [CrossRef] [PubMed]
- Loh, B.; Singh-Gondil, V.; Manohar, P.; Mehmood-Khan, F.; Yang, H.; Leptihn, S. Encapsulation and delivery of therapeutic Phages. Appl. Environ. Microbiol. 2021, 87. [Google Scholar] [CrossRef] [PubMed]
- Malik, D.J.; Sokolov, I.J.; Vinner, G.K.; Mancuso, F.; Cinquerrui, S.; Vladisavljevic, G.T.; Clokie, M.; Garton, N.J.; Stapley, A.; Kirpichnikova, A. Formulation, stabilisation and encapsulation of bacteriophage for phage therapy. Adv. Colloid Interface Sci. 2017, 249, 100–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yongsheng, M.; Pacan, J.C.; Wang, Q.; Xu, Y.; Huang, X.; Korenevsky, A.; Sabour, P.M. Microencapsulation of bacteriophage felix o1 into chitosan-alginate microspheres for oral delivery. Appl. Environ. Microbiol. 2008, 74, 4799–4805. [Google Scholar] [CrossRef] [Green Version]
- Segundo-Arizmendi, N.; Gómez-García, J.; Flores-Cuevas, K.G.; Duque-Montaño, B.E.; López-Villegas, E.O.; Baltazar-Hernández, E.; Torres-Angeles, O. Caracterización parcial del bacteriófago S1, lítico contra Salmonella enterica, con posible uso farmacéutico. Rev. Mex. Cienc. Farm. 2017, 48, 36–48. [Google Scholar]
- Pang, Y.; Applegate, T.J. Effects of dietary copper supplementation and copper source on digesta pH, calcium, zinc, and copper complex size in the gastrointestinal tract of the broiler chicken. Poult. Sci. 2007, 86, 531–537. [Google Scholar] [CrossRef]
- Morgan, N.K.; Walk, C.L.; Bedfort, M.R.; Burton, E.J. The effect of dietary calcium inclusion of broiler gastrointestinal pH: Quantification and method optimization. Poult. Sci. 2014, 93, 354–363. [Google Scholar] [CrossRef]
- Adams, M.H. Bacteriophages, Enumeration of bacteriophage particles. Intersci. Publ. 1959, 74, 164. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Jackman, S.D.; Vandervalk, B.P.; Mohamadi, H.; Chu, J.; Yeo, S.; Hammond, S.A.; Jahesh, G.; Khan, H.; Coombe, L.; Warren, R.L.; et al. ABySS 2.0, Resource-efficient assembly of large genomes using a Bloom filter. Genome Res. 2017, 27, 768–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Eddy, S.R. Accelerated Profile HMM Searches. PLoS Comput. Biol. 2011, 7, e1002195. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, L.; Stephens, A.; Nam, S.Z.; Rau, D.; Kübler, J.; Lozajic, M.; Gabler, F.; Söding, J.; Lupas, A.N.; Alva, V. A Completely Reimplemented MPI Bioinformatics Toolkit with a New HHpred Server at its Core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Yoshida, T.; Kuronishi, M.; Uehara, H.; Ogata, H.; Goto, S. ViPTree: The viral proteomic tree server. Bioinformatics 2017, 33, 2379–2380. [Google Scholar] [CrossRef]
- Milne, I.; Bayer, M.; Cardle, L.; Shaw, P.; Stephen, G.; Wright, F.; Marshall, D. Tablet—next generation sequence assembly visualization. Bioinformatics 2010, 26, 401–402. [Google Scholar] [CrossRef] [Green Version]
- Carver, T.; Harris, S.R.; Berriman, M.; Parkhill, J.; McQuillan, J.A. Artemis: An integrated platform for visualization and analysis of high-throughput sequence-based experimental data. Bioinformatics 2012, 28, 464–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carver, T.; Thomson, N.; Bleasby, A.; Berriman, M.; Parkhill, J. DNAPlotter: Circular and linear interactive genome visualization. Bioinformatics 2009, 25, 119–120. [Google Scholar] [CrossRef] [PubMed]
- Labrie, S.J.; Mosterd, C.; Loignon, S.; Dupuis, M.E.; Desjardins, P.; Tremblay, D.M.; Romero, D.A.; Horvath, P.; Fremaux, C.; Moineau, S. A mutation in the methionine aminopeptidase gene provides phage resistance in Streptococcus thermophilus. Sci. Rep. 2019, 9, 13816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tynecki, P.; Guziński, A.; Kazimierczak, J.; Jadczuk, M.; Dastych, J.; Onisko, A. PhageAI—Bacteriophage Life Cycle Recognition with Machine Learning and Natural Language Processing. BioRxiv 2020. [Google Scholar] [CrossRef]
- Montso, P.K.; Mlambo, V.; Ateba, C.N. Characterization of Lytic Bacteriophages Infecting Multidrug-Resistant Shiga Toxigenic Atypical Escherichia coli O177 Strains Isolated From Cattle Feces. Front. Public Health 2019, 7, 355. [Google Scholar] [CrossRef] [PubMed]
- Colom, J.; Cano-Sarabia, M.; Otero, J.; Cortés, P.; Maspoch, D.; Llagostera, M. Liposome-Encapsulated Bacteriophages for Enhanced Oral Phage Therapy against Salmonella spp. Appl. Environ. Microbiol. 2015, 81, 4841–4849. [Google Scholar] [CrossRef] [Green Version]
- Boggione, D.; Batalha, L.S.; Gontijo, M.; Lopez, M.; Teixeira, A.; Santos, I.; Mendonça, R. Evaluation of microencapsulation of the UFV-AREG1 bacteriophage in alginate-Ca microcapsules using microfluidic devices. Colloids Surf. B Biointerfaces 2017, 158, 182–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravindran, V. Feed enzymes: The science, practice, and metabolic realities. J. Appl. Poult. Res. 2013, 22, 628–636. [Google Scholar] [CrossRef]
- Jajere, S.M. A review of Salmonella enterica with particular focus on the pathogenicity and virulence factors, host specificity and antimicrobial resistance icluding multidrug resistence. Vet. World 2019, 12, 504–521. [Google Scholar] [CrossRef] [Green Version]
- NOM-051-ZOO-1995. Trato Humanitario en la Movilización de Animales. Available online: https://www.gob.mx/cms/uploads/attachment/file/563487/NOM-051-ZOO-1995_230398.pdf (accessed on 1 January 2020).
- American Society of Animal Science. Guide for the Care and Use of Agricultural Animals in Research and Teaching, 3rd ed. Available online: https://www.asas.org/services/ag-guide (accessed on 1 April 2020).
- Manual of Good Livestock Practices in Broiler Production Units, 2nd ed. Available online: http://oncesega.org.mx/archivos/Manual_de_Buenas_Pr_cticas_Pecuarias_de_Producci_n_de_Pollo_de_Engorda_4.pdf (accessed on 1 January 2020).
- NOM-033-SAG/ZOO-2014. Métodos Para dar Muerte a los Animales Domésticos y Silvestres. Available online: https://www.gob.mx/senasica/documentos/nom-033-sag-zoo-2014?state=published (accessed on 1 January 2020).
- Gambino, M.; Nørgaard-Sørensen, A.; Ahern, S.; Smyrlis, G.; Gencay, Y.E.; Hendrix, H.; Neve, H.; Noben, J.P.; Lavigne, R.; Brøndsted, L. Phage S144, A New Polyvalent Phage Infecting Salmonella spp. and Cronobacter sakazakii. Int. J. Mol. 2020, 21, 5196. [Google Scholar] [CrossRef]
- Mohamed, A.; Taha, O.; El-Sherif, H.M.; Connerton, P.L.; Hooton, S.P.T.; Basim, N.D.; Connerton, I.F.; El-Shibiny, A. Bacteriophage ZCSE2 is a Potent Antimicrobial Against Salmonella enterica Serovars: Ultrastructure, Genomics and Efficacy. Viruses 2020, 12, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fong, K.; LaBossiere, B.; Switt, A.I.M.; Delaquis, P.; Goodridge, L.; Levesque, R.C.; Danyuk, M.D.; Wang, S. Characterization of Four Novel Bacteriophages Isolated from British Columbia for Control of Non-typhoidal Salmonella in vitro and on Sprouting Alfalfa Seeds. Front. Microbiol. 2017, 8, 2193. [Google Scholar] [CrossRef] [PubMed]
- Porwollik, S.; Wong, R.M.; McClelland, M. Evolutionary genomics of Salmonella: Gene acquisitions revealed by microarray analysis. Proc. Natl. Acad. Sci. USA 2002, 99, 8956–8961. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Wang, Z.; Qiang, B.; Xu, Y.; Chen, X.; Li, Q.; Jiao, X. Loss and Gain in the Evolution of the Salmonella enterica Serovar Gallinarum Biovar Pullorum Genome. Msphere 2019, 4, e00627-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Johnston, R.N.; Liu, G.R.; Liu, S.L.; Feng, Y. Genomic comparison between Salmonella Gallinarum and Pullorum: Differential pseudogene formation under common host restriction. PLoS ONE 2013, 8, e59427. [Google Scholar] [CrossRef] [Green Version]
- Lupo, B.; González, C.; Maestro, A. Microencapsulación con alginato en alimentos. Técnicas y aplicaciones. Revista Venezolana de Ciencia y Tecnología de Alimentos 2012, 3, 130–151. [Google Scholar]
- Aguado, B.A.; Mulyasasmita, W.; Su, J.; Lampe, K.J.; Heilshorn, S.C. Improving viability of stem cells during syringe needle flow through the design of hydrogel cell carriers. Tissue Eng. Part A 2012, 18, 806–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Menéndez, E.; Fernández, L.; Gutiérrez, D.; Pando, D.; Martínez, B.; Rodríguez, A.; García, P. Strategies to Encapsulate the Staphylococcus aureus Bacteriophage phiIPLA-RODI. Viruses 2018, 10, 495. [Google Scholar] [CrossRef] [Green Version]
- Abdelsattar, A.S.; Abdelrahman, F.; Dawoud, A.; Connerton, I.F.; El-Shibiny, A. Encapsulation of E. coli phage ZCEC5 in chitosan–alginate beads as a delivery system in phage therapy. AMB Express 2019, 9, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amerah, A.M.; Ravindran, V.; Lentle, R.G.; Thomas, D.G. Influence of feed particle size and feed form on the performance, energy utilization, digestive tract development, and digesta parameters of broiler starters. Poult. Sci. 2007, 86, 2615–2623. [Google Scholar] [CrossRef]
- Chewning, C.G.; Stark, C.R.; Brake, J. Effects of particle size and feed form on broiler performance. J. Appl. Poult. Res. 2012, 21, 830–837. [Google Scholar] [CrossRef]
- Zaefarian, F.; Abdollahi, M.R.; Ravindran, V. Particle size and feed form in broiler diets: Impact on gastrointestinal tract development and gut health. Worlds Poult. Sci. J. 2016, 72, 277–290. [Google Scholar] [CrossRef] [Green Version]
- Moghtader, F.; Eğri, S.; Piskin, E. Phages in modified alginate beads. Artif. Cells Nanomed. Biotechnol. 2017, 45, 357–363. [Google Scholar] [CrossRef] [Green Version]
- Mahony, J.; McAuliffe, O.; Ross, R.P.; van Sinderen, D. Bacteriophages as biocontrol agents of food pathogens. Curr. Opin. Biotechnol. 2011, 22, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Kutateladze, M. Experience of the Eliava Institute in bacteriophage therapy. Virol. Sin. 2015, 30, 80–81. [Google Scholar] [CrossRef] [PubMed]
- Hjorth, T.; Karlsen, J. Alginate in Drug Delivery Systems. Drug Dev. Ind. Pharm. 2002, 28, 621–630. [Google Scholar] [CrossRef]
- Gheorghita-Puscaselu, R.; Lobiuc, A.; Dimian, M.; Covasa, M. Alginate: From food industry to biomedical applications and management of metabolic disorders. Polymers 2020, 12, 2417. [Google Scholar] [CrossRef]
- Lee, K.Y.; Mooney, D.J. Alginate: Properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Flores, G.; Casas, F.C.; Quintana-López, J.A.; Cedillo-Peláez, C.; Urquiza-Bravo, O. Patogenia de Salmonella enteritidis FT 13a y Salmonella enteritidis biovar Issatschenko en pollos de engorda. Vet. México 2008, 39, 145–160. [Google Scholar]
- Svihus, B. The gizzard: Function, influence of diet structure and effects on nutrient availability. Worlds Poult. Sci. J. 2011, 67, 207–224. [Google Scholar] [CrossRef]
- Roa, I.; Meruane, M. Desarrollo del Aparato Digestivo. Int. J. Morphol. 2012, 30, 1285–1294. [Google Scholar] [CrossRef] [Green Version]
- Barrow, G.; Feltham, R. Enterobacteria. In Cowan and Steel’s Manual for the Identification of Medical Bacteria; Barrow, G.I., Feltham, R.K.A., Eds.; Cambridge University Press: Cambridge, UK, 1993; p. 141. [Google Scholar]
- Bardina, C.; Spricigo, D.A.; Cortés, P.; Llagostera, M. Significance of the bacteriophage treatment schedule in reducing Salmonella colonization of poultry. Appl. Environ. Microbiol. 2012, 78, 6600–6607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grygorcewicz, B.; Grudziński, M.; Wasak, A.; Augustyniak, A.; Pietruszka, A.; Nawrotek, P. Bacteriophage-mediated reduction of Salmonella Enteritidis in swine slurry. Appl. Soil. Ecol. 2017, 119, 179–182. [Google Scholar] [CrossRef]
- Cristobal-Cueto, P.; García-Quintanilla, A.; Esteban, J.; García-Quintanilla, M. Phages in Food Industry Biocontrol and Bioremediation. Antibiotics 2021, 10, 786. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Infection | Treatments | ||
|---|---|---|---|---|
| Day 1 | Day 2 | Day3 | Day 8 | |
| A | No infection | ALG beads + S1 at 4.9 × 109 PFU | ||
| B | S.E. at 4 × 106 UFC | S.E. at 4 × 107 UFC | S.E. at 2.5 × 109 UFC | No treatment |
| C | Antibiotic treatment | |||
| D | ALG beads + S1 at 4.9 × 109 PFU | |||
| E | ALG beads | |||
| ORF | Product | Chromosomal Locus (nt) | Accession Number | Database | e-Value |
|---|---|---|---|---|---|
| 1 | Major capsid protein | 1–978 | YP009821716.1 | NCBI | 5.2 × 10213 |
| 6 | Putative tail protein 1 | 2907–3527 | QMV47842.1 | NCBI | 3 × 10−134 |
| 7 | Putative tail protein 2 | 3527–6484 | Q6QGE2 | UniProtKB | 0 |
| 8 | Putative tail protein 3 | 6554–7195 | Q6QGE2 | UniProtKB | 9.5 × 10−10 |
| 9 | Putative tail protein 4 | 7208–9082 | QMV47844.1 | NCBI | 6.8 × 10−32 |
| 10 | Putative sheath protein | 9170–10,309 | Q24LI4 | UniProtKB | 5.6 × 10−19 |
| 11 | Putative structural protein | 10,320–10,751 | D6RRG7 | UniProtKB | 1.8 × 10−13 |
| 12 | Putative tail assembly chaperone protein | 10,768–11,196 | PF10876.10 | Pfam | 5.6 × 10−15 |
| 13 | Putative tape measure protein | 11,362–13,023 | Q6KGH8 | UniProtKB | 7.2 × 10−06 |
| 16 | Putative baseplate protein 1 | 14,250–15,200 | 10312 | UniProtKB | 1.8 × 10−02 |
| 17 | Putative puncturing protein | 15,190–15,834 | PF18352.3 | Pfam | 5 × 10−18 |
| 18 | Putative baseplate protein 2 | 15,843–16,214 | P09425 | UniProtKB | 4.1 × 10−7 |
| 19 | Putative baseplate protein 3 | 16,218–17,381 | Q9T1V2 | UniProtKB | 3 × 10−10 |
| 21 | Tail fiber protein S | 18,020–19,369 | Q9T1V0 | UniProtKB | 6.9 × 10−18 |
| 22 | Tail fiber assembly protein U | 19,369–19,911 | Q71TD6 | UniProtKB | 2.9 × 10−18 |
| 23 | Putative tail fiber assembly protein U’ | 19,914–20,423 | Q71TD7 | UniProtKB | 6.9 × 10−18 |
| 24 | Putative holin | 20,525–20,788 | PF16080.7 | Pfam | 6.7 × 10−04 |
| 25 | SAR-endolysin | 20,763–21,308 | Q37875 | UniProtKB | 7.4 × 10−13 |
| 26 | Putative spanin inner membrane subunit | 21,287–21,616 | Q9T1X1 | UniProtKB | 2 × 10−9 |
| 29 | DNA polymerase | 22,342–24,312c | P19822 | UniProtKB | 1.5 × 10−49 |
| 31 | Thymidylate synthase | 25,340–26,224c | P00471 | UniProtKB | 3.4 × 10−40 |
| 36 | Putative ATP-dependent DNA helicase | 28,586–30,259c | P20703 | UniProtKB | 2.3 × 10−32 |
| 38 | Putative exonuclease | 31,377–30,466c | P03697 | UniProtKB | 1.8 × 10−04 |
| 39 | Putative deoxycytidylate deaminase | 31,343–31,807c | P00814 | UniProtKB | 1.4 × 10−13 |
| 54 | Anaerobic NTP reductase small subunit | 38,921–41,455 | YP009821771.1 | NCBI | 0 |
| 55 | Putative resolvase | 41,950–42,378 | Q98VP9 | UniProtKB | 1.6 × 10−04 |
| 68 | Putative dihydrofolate reductase | 46,756–47,427 | Q6QGJ4 | UniProtKB | 6.4 × 10−06 |
| 73 | Putative terminase large subunit | 49,484–50,932 | P54308 | UniProtKB | 6.3 × 10−16 |
| 74 | Portal protein | 50,934–52,496 | O64207 | UniProtKB | 6.5 × 10−10 |
| Bacteria | Identification of Bacteria | EOP |
|---|---|---|
| S. Enteritidis (SE PT13A) | Host strain | 1 |
| S. Typhi | ICTH | 0 |
| S. Pullorum | ICTP | 0.61 |
| S. Gallinarum | ICTP | 0.58 |
| S. Cholerasuis | ICTH | 0 |
| C. freundii | ICTH | 0 |
| E. cloacae | ICTH | 0 |
| E. coli | ICTH | 0 |
| Section of the GIT | 1 H Post Infection | 3 H Post Infection | 5 H Post Infection | 17 H Post Infection | ||||
|---|---|---|---|---|---|---|---|---|
| C #1 | C #2 | C #1 | C #2 | C #1 | C #2 | C #1 | C #2 | |
| Crop | + | + | − | + | − | + | − | − |
| Gizzard | − | − | − | − | − | − | − | − |
| Duodenum | − | − | − | + | − | + | − | − |
| Cecum | − | − | + | + | − | + | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomez-Garcia, J.; Chavez-Carbajal, A.; Segundo-Arizmendi, N.; Baron-Pichardo, M.G.; Mendoza-Elvira, S.E.; Hernandez-Baltazar, E.; Hynes, A.P.; Torres-Angeles, O. Efficacy of Salmonella Bacteriophage S1 Delivered and Released by Alginate Beads in a Chicken Model of Infection. Viruses 2021, 13, 1932. https://doi.org/10.3390/v13101932
Gomez-Garcia J, Chavez-Carbajal A, Segundo-Arizmendi N, Baron-Pichardo MG, Mendoza-Elvira SE, Hernandez-Baltazar E, Hynes AP, Torres-Angeles O. Efficacy of Salmonella Bacteriophage S1 Delivered and Released by Alginate Beads in a Chicken Model of Infection. Viruses. 2021; 13(10):1932. https://doi.org/10.3390/v13101932
Chicago/Turabian StyleGomez-Garcia, Janeth, Alejandra Chavez-Carbajal, Nallelyt Segundo-Arizmendi, Miriam G. Baron-Pichardo, Susana E. Mendoza-Elvira, Efren Hernandez-Baltazar, Alexander P. Hynes, and Oscar Torres-Angeles. 2021. "Efficacy of Salmonella Bacteriophage S1 Delivered and Released by Alginate Beads in a Chicken Model of Infection" Viruses 13, no. 10: 1932. https://doi.org/10.3390/v13101932