Urine and Free Immunoglobulin Light Chains as Analytes for Serodiagnosis of Hantavirus Infection

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Samples and Ethics Statement
2.2. Proteins and Antibodies
2.3. LFRET Assays
2.4. Immunofluorescence Assay (IFA)
2.5. Western Blot (WB)
2.6. Immunoprecipitation (IP) of FLCs and PUUV N Protein
2.7. Purification and Analysis of Binding Specificities of Free Light Chains in Urine
2.8. Reference Diagnostic Methods
2.9. Statistics
3. Results
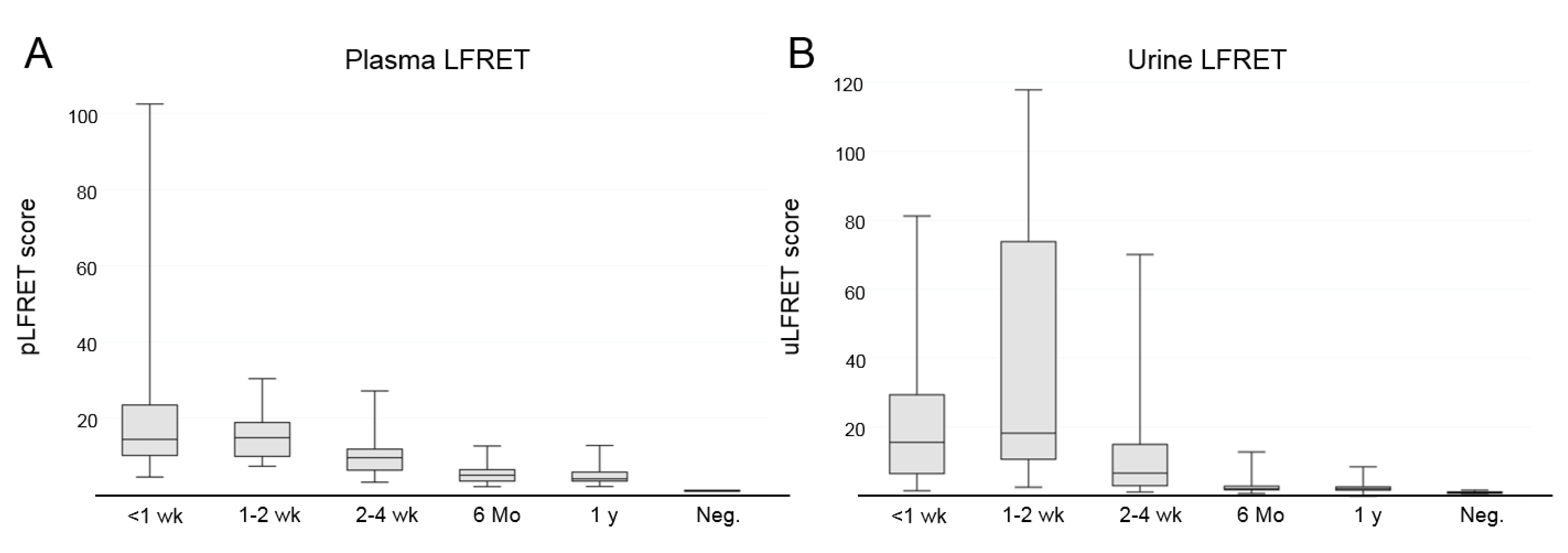
3.1. Detection of Hantavirus-Specific Antibody Responses from Urine Using LFRET
3.2. IgG, IgM, and IgA Antibodies to PUUV in Urine by IFA
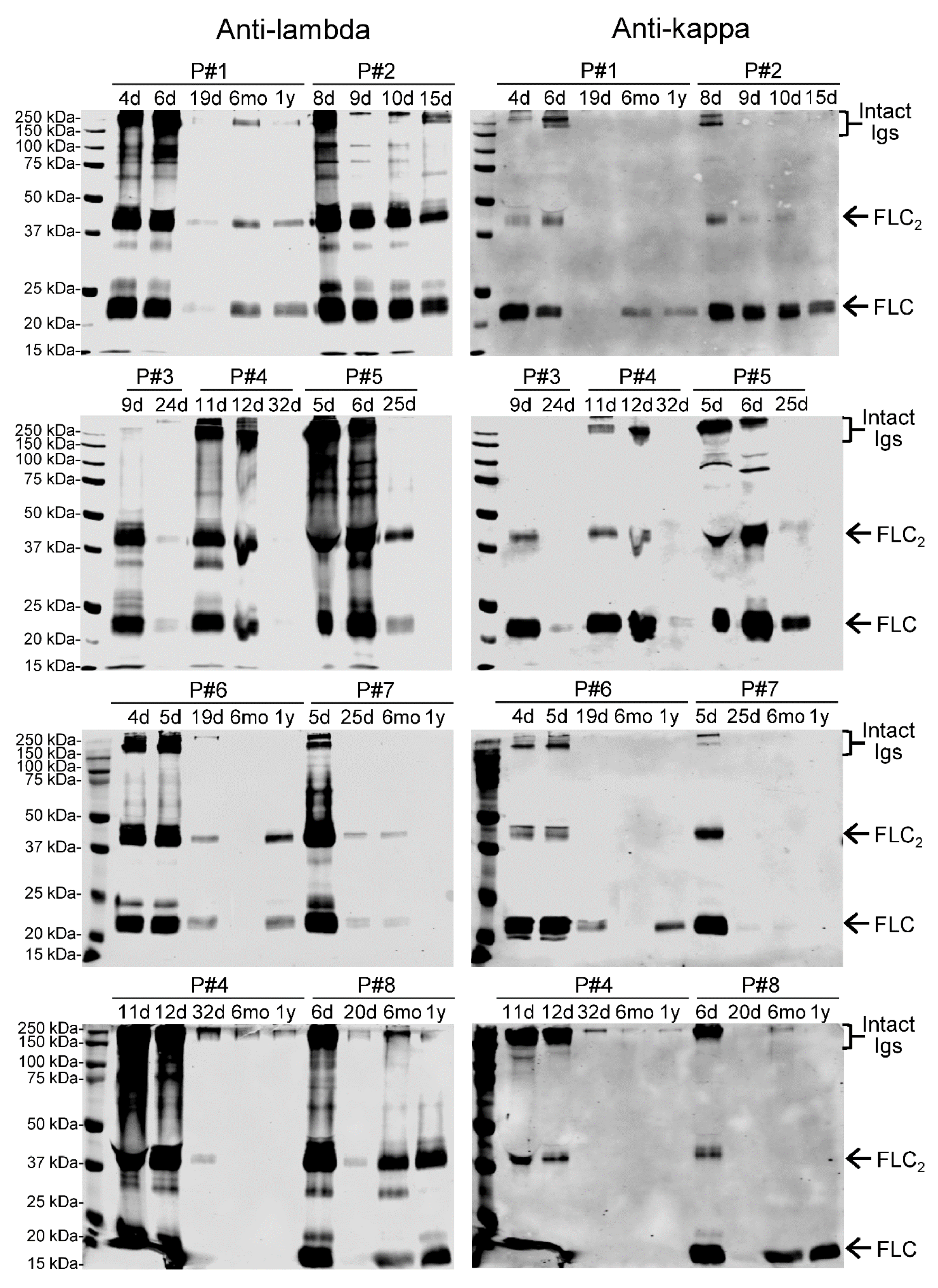
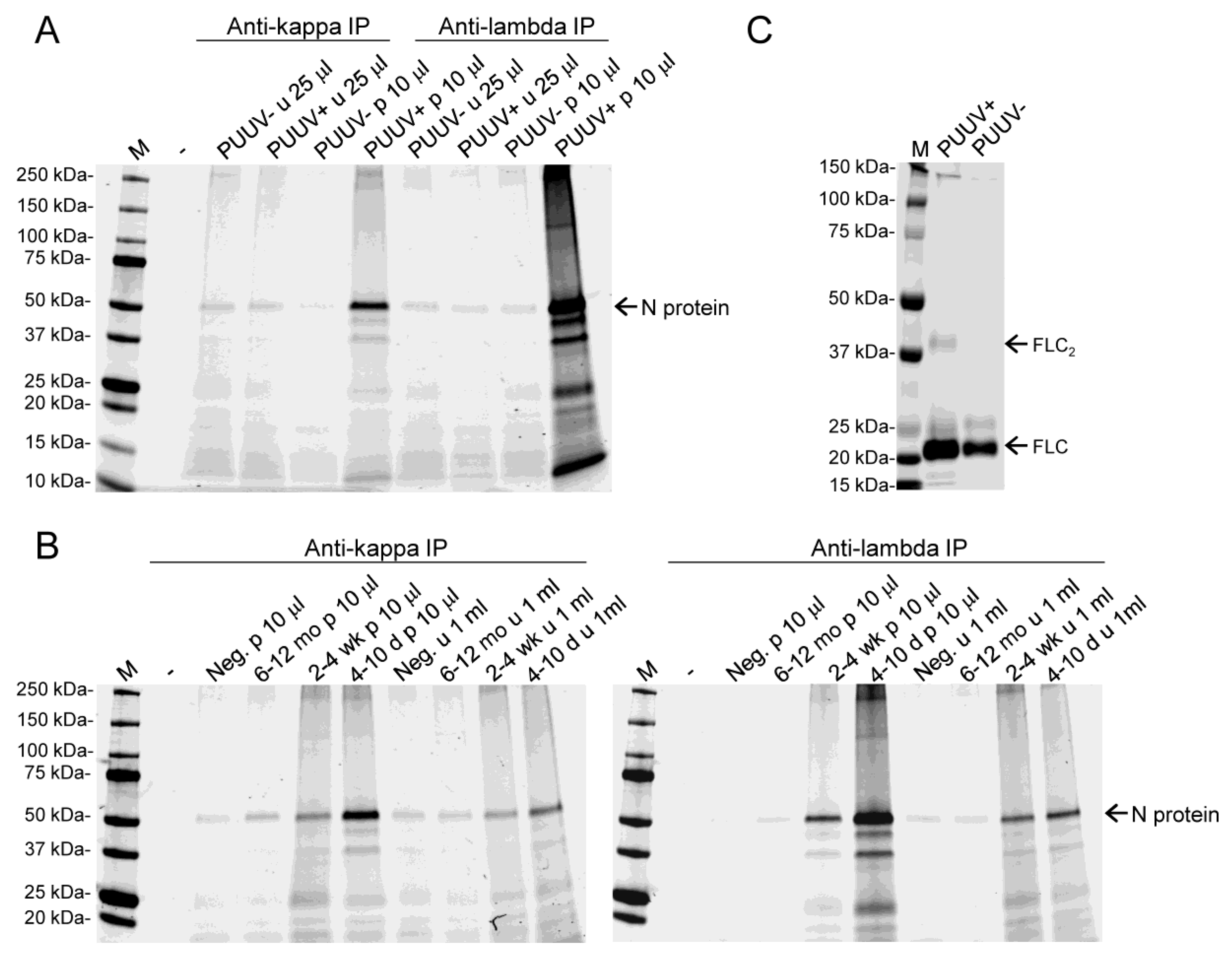
3.3. Both Kappa and Lambda FLCs Specifically Bind PUUV N Protein
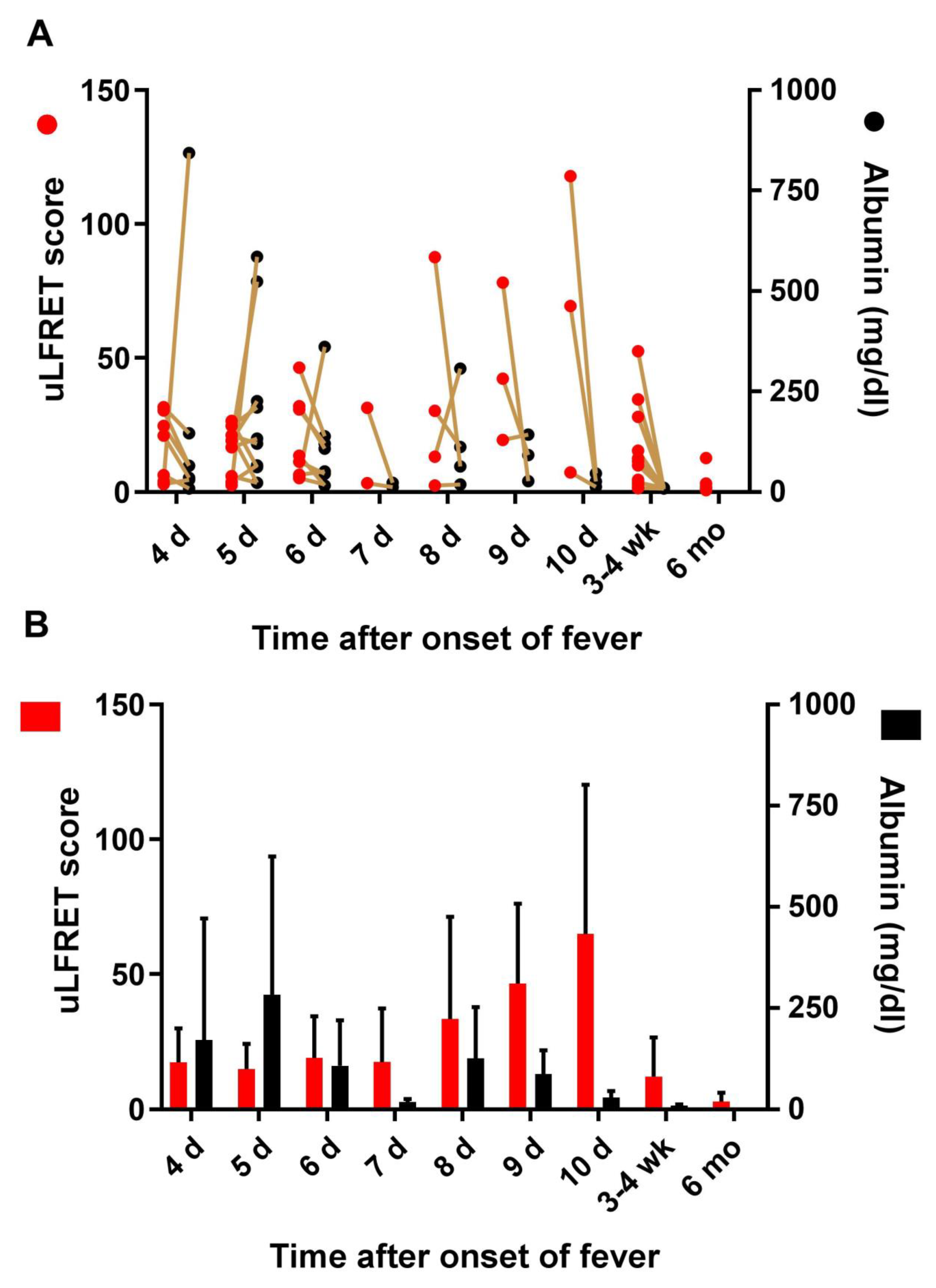
3.4. Lack of Correlation Between LFRET Signal and Proteinuria
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hedman, K.; Hepojoki, J.; Hepojoki, S.; Vaheri, A.; Vapalahti, O. Anonymous Protein L Based Bioassay Method for Determining Presence of Soluble Antibodies in a Sample and Kit Therefor. Finland Patent nr. WO2015128548A1, 30 September 2015. [Google Scholar]
- Hepojoki, S.; Rusanen, J.; Hepojoki, J.; Nurmi, V.; Vaheri, A.; Lundkvist, Å.; Hedman, K.; Vapalahti, O. Competitive Homogeneous Immunoassay for Rapid Serodiagnosis of Hantavirus Disease. J. Clin. Microbiol. 2015, 53, 2292–2297. [Google Scholar] [CrossRef] [Green Version]
- Hepojoki, S.; Hepojoki, J.; Hedman, K.; Vapalahti, O.; Vaheri, A. Rapid homogeneous immunoassay based on time-resolved forster resonance energy transfer for serodiagnosis of acute hantavirus infection. J. Clin. Microbiol. 2015, 53, 63–640. [Google Scholar] [CrossRef]
- Hepojoki, S.; Nurmi, V.; Vaheri, A.; Hedman, K.; Vapalahti, O.; Hepojoki, J. A protein L-based immunodiagnostic approach utilizing time-resolved forster resonance energy transfer. PLoS ONE 2014, 9, e106432. [Google Scholar] [CrossRef]
- Åkerstrom, B.; Björck, L. Protein L: An immunoglobulin light chain-binding bacterial protein. Characterization of binding and physicochemical properties. J. Biol. Chem. 1989, 264, 19740–19746. [Google Scholar]
- Schroeder, H.W.; Cavacini, L. Structure and function of immunoglobulins. J. Allergy Clin. Immunol. 2010, 125, S41–S52. [Google Scholar] [CrossRef] [Green Version]
- Vaheri, A.; Strandin, T.; Hepojoki, J.; Sironen, T.; Henttonen, H.; Makela, S.; Mustonen, J. Uncovering the mysteries of hantavirus infections. Nat. Rev. Microbiol. 2013, 11, 539–550. [Google Scholar] [CrossRef]
- Vaheri, A.; Henttonen, H.; Voutilainen, L.; Mustonen, J.; Sironen, T.; Vapalahti, O. Hantavirus infections in Europe and their impact on public health. Rev. Med. Virol. 2013, 23, 35–49. [Google Scholar] [CrossRef]
- Vapalahti, O.; Mustonen, J.; Lundkvist, Å.; Henttonen, H.; Plyusnin, A.; Vaheri, A. Hantavirus infections in Europe. Lancet Infect. Dis. 2003, 3, 653–661. [Google Scholar] [CrossRef]
- Mustonen, J.; Helin, H.; Pietila, K.; Brummer-Korvenkontio, M.; Hedman, K.; Vaheri, A.; Pasternack, A. Renal biopsy findings and clinicopathologic correlations in nephropathia epidemica. Clin. Nephrol. 1994, 41, 121–126. [Google Scholar]
- Mustonen, J.; Outinen, T.; Laine, O.; Porsti, I.; Vaheri, A.; Makela, S. Kidney disease in Puumala hantavirus infection. Infect. Dis. (Lond.) 2017, 49, 321–332. [Google Scholar] [CrossRef]
- Temonen, M.; Mustonen, J.; Helin, H.; Pasternack, A.; Vaheri, A.; Holthofer, H. Cytokines, adhesion molecules, and cellular infiltration in nephropathia epidemica kidneys: An immunohistochemical study. Clin. Immunol. Immunopathol. 1996, 78, 47–55. [Google Scholar] [CrossRef]
- Mantula, P.S.; Outinen, T.K.; Clement, J.P.G.; Huhtala, H.S.A.; Porsti, I.H.; Vaheri, A.; Mustonen, J.T.; Makela, S.M. Glomerular Proteinuria Predicts the Severity of Acute Kidney Injury in Puumala Hantavirus-Induced Tubulointerstitial Nephritis. Nephron 2017, 136, 193–201. [Google Scholar] [CrossRef]
- Settergren, B. Clinical aspects of nephropathia epidemica (Puumala virus infection) in Europe: A review. Scand. J. Infect. Dis. 2000, 32, 125–132. [Google Scholar] [CrossRef]
- Settergren, B.; Juto, P.; Trollfors, B.; Wadell, G.; Norrby, S.R. Hemorrhagic complications and other clinical findings in nephropathia epidemica in Sweden: A study of 355 serologically verified cases. J. Infect. Dis. 1988, 157, 380–382. [Google Scholar] [CrossRef]
- Kramski, M.; Achazi, K.; Klempa, B.; Kruger, D.H. Nephropathia epidemica with a 6-week incubation period after occupational exposure to Puumala hantavirus. J. Clin. Virol. 2009, 44, 99–101. [Google Scholar] [CrossRef]
- Kallio-Kokko, H.; Leveelahti, R.; Brummer-Korvenkontio, M.; Lundkvist, A.; Vaheri, A.; Vapalahti, O. Human immune response to Puumala virus glycoproteins and nucleocapsid protein expressed in mammalian cells. J. Med. Virol. 2001, 65, 605–613. [Google Scholar] [CrossRef]
- Hedman, K.; Vaheri, A.; Brummer-Korvenkontio, M. Rapid diagnosis of hantavirus disease with an IgG-avidity assay. Lancet 1991, 338, 1353–1356. [Google Scholar] [CrossRef]
- Hujakka, H.; Koistinen, V.; Eerikainen, P.; Kuronen, I.; Mononen, I.; Parviainen, M.; Lundkvist, A.; Vaheri, A.; Narvanen, A.; Vapalahti, O. New immunochromatographic rapid test for diagnosis of acute Puumala virus infection. J. Clin. Microbiol. 2001, 39, 2146–2150. [Google Scholar] [CrossRef]
- Hopper, J.E.; Papagiannes, E. Evidence by radioimmunoassay that mitogen-activated human blood mononuclear cells secrete significant amounts of light chain Ig unassociated with heavy chain. Cell. Immunol. 1986, 101, 122–131. [Google Scholar] [CrossRef]
- Redegeld, F.A.; van der Heijden, M.W.; Kool, M.; Heijdra, B.M.; Garssen, J.; Kraneveld, A.D.; van Loveren, H.; Roholl, P.; Saito, T.; Verbeek, J.S.; et al. Immunoglobulin-free light chains elicit immediate hypersensitivity-like responses. Nat. Med. 2002, 8, 694–701. [Google Scholar] [CrossRef]
- Hannam-Harris, A.C.; Smith, J.L. Free immunoglobulin light chain synthesis by human foetal liver and cord blood lymphocytes. Immunology 1981, 43, 417–423. [Google Scholar]
- Skvortsov, V.T.; Gurvich, A.E. Relative rates of synthesis of immunoglobulins and light chains in rabbit spleen cells during secondary response. Nature 1968, 218, 377–378. [Google Scholar] [CrossRef]
- Shapiro, A.L.; Scharff, M.D.; Maizel, J.V.; Uhr, J.W. Synthesis of excess light chains of gamma globulin by rabbit lymph node cells. Nature 1966, 211, 243–245. [Google Scholar] [CrossRef]
- Abe, M.; Goto, T.; Kosaka, M.; Wolfenbarger, D.; Weiss, D.T.; Solomon, A. Differences in kappa to lambda (kappa:lambda) ratios of serum and urinary free light chains. Clin. Exp. Immunol. 1998, 111, 457–462. [Google Scholar] [CrossRef]
- Saraheimo, S.; Hepojoki, J.; Nurmi, V.; Lahtinen, A.; Hemmilä, I.; Vaheri, A.; Vapalahti, O.; Hedman, K.A. Time-resolved FRET-based approach for antibody detection—A new serodiagnostic concept. PLoS ONE 2013, 8, e62739. [Google Scholar] [CrossRef]
- Vapalahti, O.; Lundkvist, Å.; Kallio-Kokko, H.; Paukku, K.; Julkunen, I.; Lankinen, H.; Vaheri, A. Antigenic properties and diagnostic potential of puumala virus nucleocapsid protein expressed in insect cells. J. Clin. Microbiol. 1996, 34, 119–125. [Google Scholar] [Green Version]
- Libraty, D.H.; Makela, S.; Vlk, J.; Hurme, M.; Vaheri, A.; Ennis, F.A.; Mustonen, J. The degree of leukocytosis and urine GATA-3 mRNA levels are risk factors for severe acute kidney injury in Puumala virus nephropathia epidemica. PLoS ONE 2012, 7, e35402. [Google Scholar] [CrossRef]
- Makela, S.; Ala-Houhala, I.; Mustonen, J.; Koivisto, A.M.; Kouri, T.; Turjanmaa, V.; Vapalahti, O.; Vaheri, A.; Pasternack, A. Renal function and blood pressure five years after puumala virus-induced nephropathy. Kidney Int. 2000, 58, 1711–1718. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time after Onset of Fever | 4–5 d | 6–7 d | 1–2 wk | 2–4 wk | 6 mo | 1 y |
|---|---|---|---|---|---|---|
| Number of urine samples | 24 | 30 | 20 | 34 | 27 | 26 |
| uLFRET positive | 23 | 30 | 20 | 29 | 11 | 11 |
| Sensitivity | 95.8% | 100.0% | 100.0% | 85.3% | 40.7% | 42.3% |
| Number of plasma samples | 23 | 29 | 20 | 34 | 22 | 25 |
| pLFRET positive | 23 | 29 | 20 | 34 | 21 | 25 |
| Sensitivity | 100.0% | 100.0% | 100.0% | 100.0% | 95.5% | 100.0% |
| Patient 1 | IFA IgG | IFA IgM | IFA IgA | uLFRET |
|---|---|---|---|---|
| 5d * | neg | neg | neg | pos (19) |
| 6d | pos (>10) | neg | neg | pos (46) |
| 7d | pos (5) | neg | neg | pos (32) |
| 21d | pos (2) | neg | neg | pos (4) |
| Patient 2 | ||||
| 8d | pos (>10) | neg | neg | pos (88) |
| 9d | pos (>10) | neg | neg | pos (78) |
| 10d | pos (2) | neg | neg | pos (7) |
| 24d | pos (2) | neg | neg | pos (11) |
| Patient 3 | ||||
| 7d | pos (>10) | neg | pos (>10) | pos (13) |
| 8d | pos (5) | neg | pos (2) | pos (42) |
| 9d | pos (5) | neg | neg | pos (69) |
| 24d | pos (2) | neg | neg | pos (52) |
| Patient 4 | ||||
| 10d | pos (>10) | neg | pos (2) | pos (118) |
| 11d | pos (5) | neg | pos (2) | pos (109) |
| 12d | pos (>10) | neg | pos (2) | pos (105) |
| 32d | neg | neg | neg | pos (70) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hepojoki, S.; Kareinen, L.; Strandin, T.; Vaheri, A.; Holthöfer, H.; Mustonen, J.; Mäkelä, S.; Hedman, K.; Vapalahti, O.; Hepojoki, J. Urine and Free Immunoglobulin Light Chains as Analytes for Serodiagnosis of Hantavirus Infection. Viruses 2019, 11, 809. https://doi.org/10.3390/v11090809
Hepojoki S, Kareinen L, Strandin T, Vaheri A, Holthöfer H, Mustonen J, Mäkelä S, Hedman K, Vapalahti O, Hepojoki J. Urine and Free Immunoglobulin Light Chains as Analytes for Serodiagnosis of Hantavirus Infection. Viruses. 2019; 11(9):809. https://doi.org/10.3390/v11090809
Chicago/Turabian StyleHepojoki, Satu, Lauri Kareinen, Tomas Strandin, Antti Vaheri, Harry Holthöfer, Jukka Mustonen, Satu Mäkelä, Klaus Hedman, Olli Vapalahti, and Jussi Hepojoki. 2019. "Urine and Free Immunoglobulin Light Chains as Analytes for Serodiagnosis of Hantavirus Infection" Viruses 11, no. 9: 809. https://doi.org/10.3390/v11090809