A Novel Bacterium-Like Particle Vaccine Displaying the MERS-CoV Receptor-Binding Domain Induces Specific Mucosal and Systemic Immune Responses in Mice

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Construction and Expression of Recombinant Baculovirus
2.2. Immunofluorescence Assay (IFA) and Western Blotting Analyses of Recombinant Baculoviruses
2.3. Binding of the Fusion Proteins to Gem Particles
2.4. SDS-PAGE, Western Blotting and IFA Analysis of the Binding GEM Particles
2.5. Immunizations and Samples Collection
2.6. Pseudotyped Virus Neutralization Assay
2.7. Enzyme-Linked Immunosorbent Assay (ELISA) Measurement of RBD-Specific Antibodies
2.8. ELISpot IFN-γ and IL-4 Cytokine Assays
2.9. Splenocyte Proliferation Assay
2.10. Evaluation of B Cell and T Cell Activation by Flow Cytometry
2.11. ELISA Measurement of Cytokine Levels in Splenocyte Culture Supernatants
2.12. Data Analysis
2.13. Laboratory Facility and Ethics Statement
3. Results
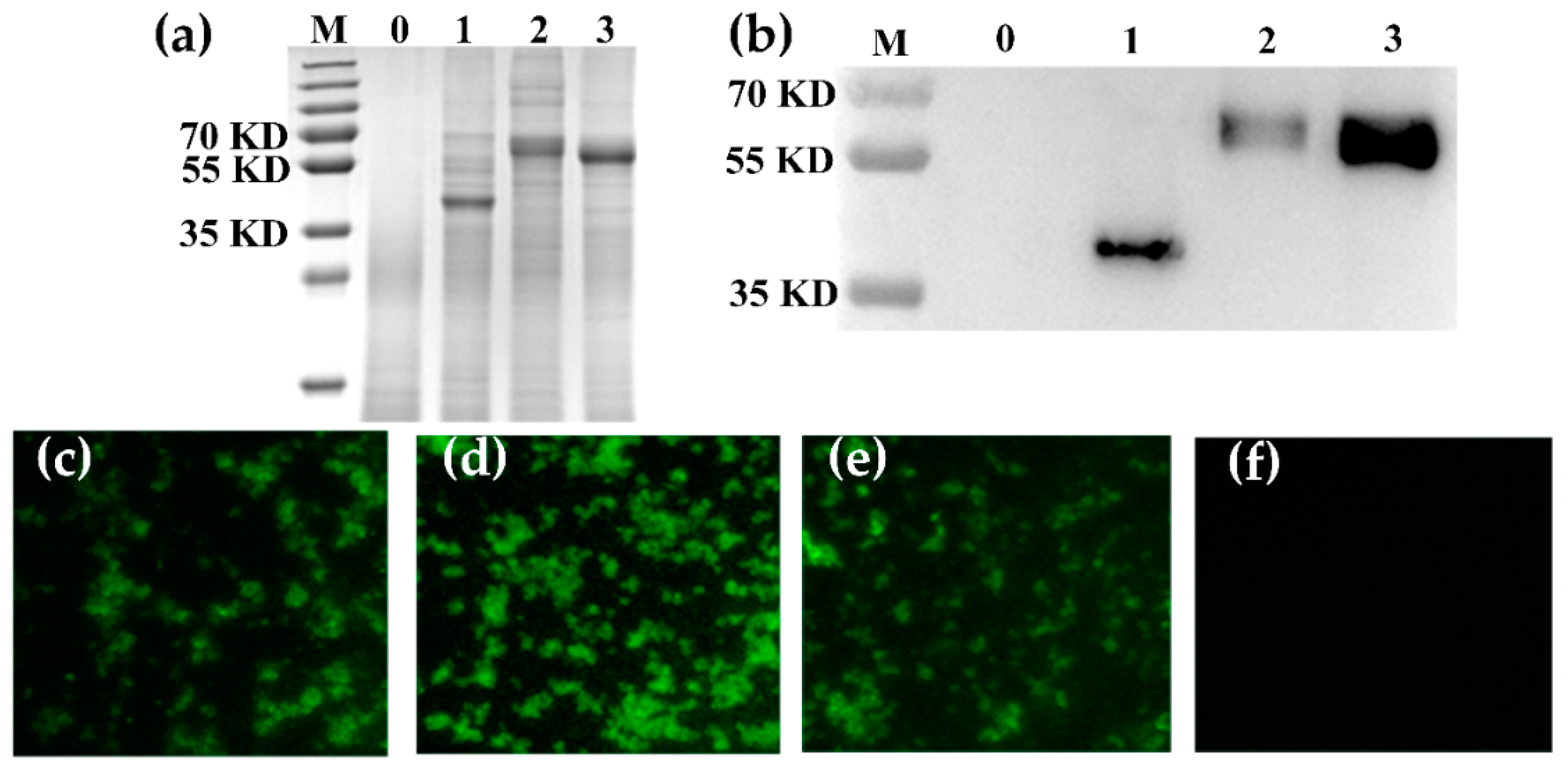
3.1. Expression of Fusion Proteins
3.2. Location of Fusion Proteins on GEM Particles
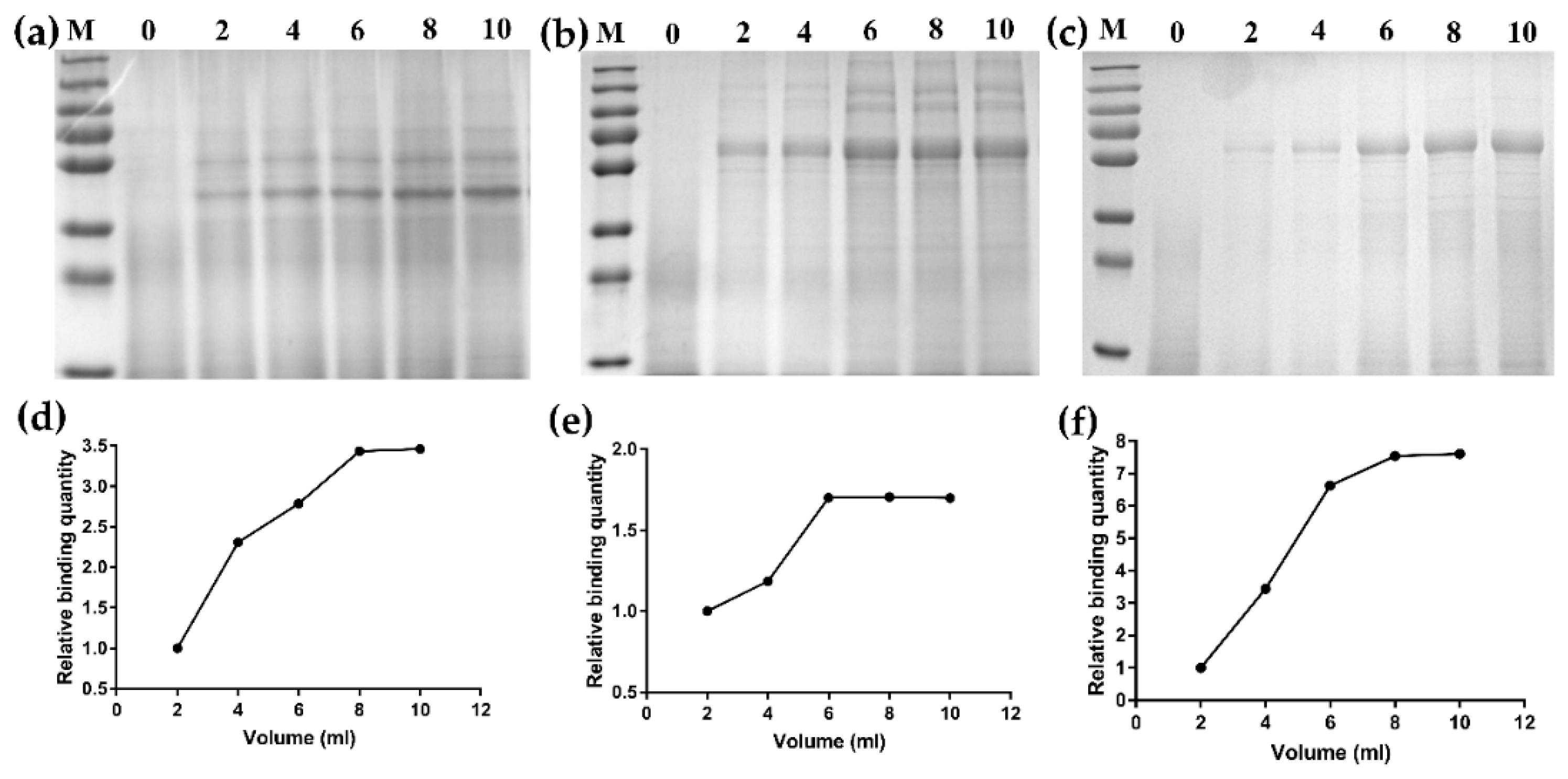
3.3. Binding Activity of Fusion Proteins on GEM Particles
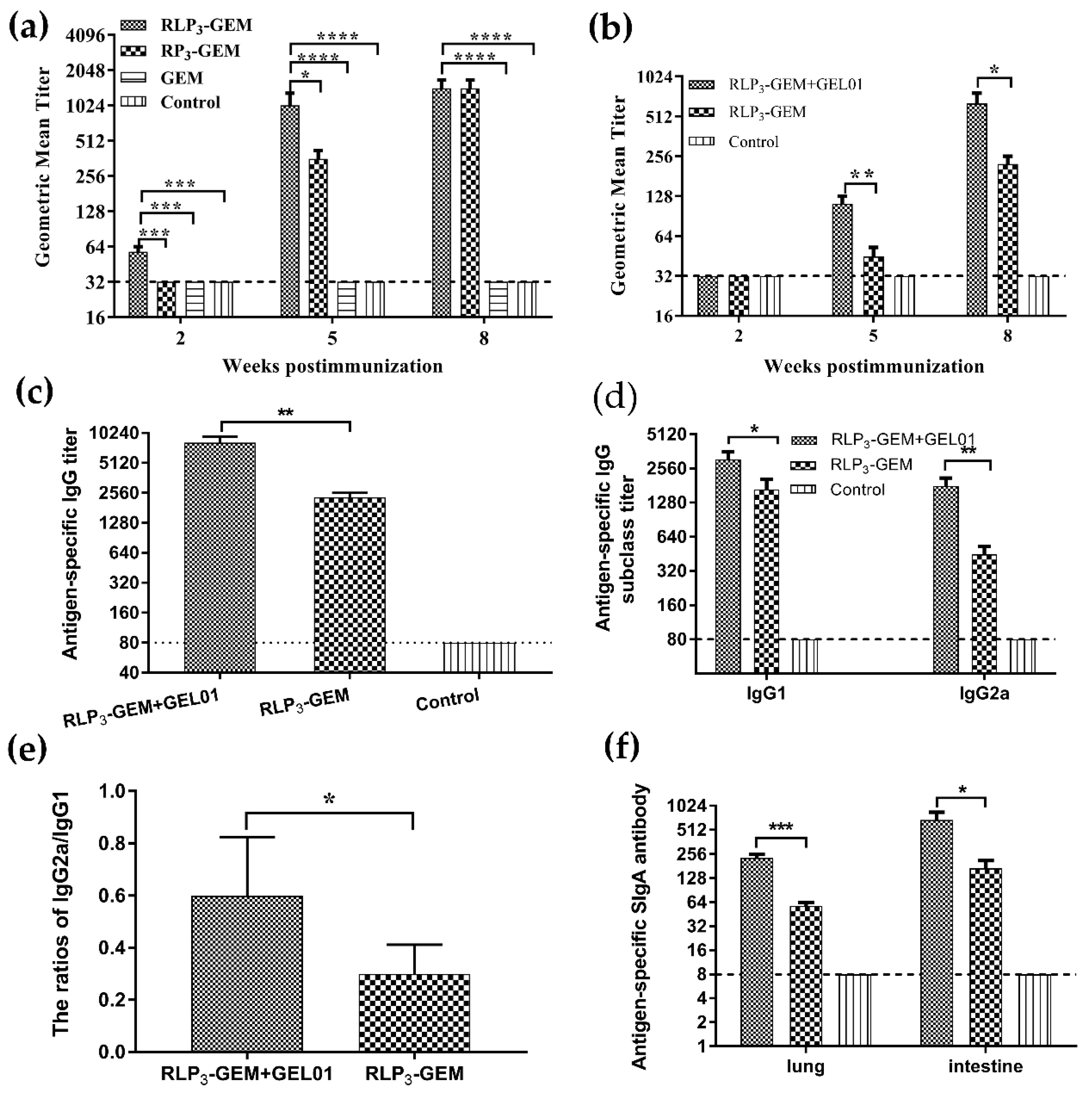
3.4. Virus Neutralizing Antibodies and ELISA Measurement of Subtype Antibodies in Serum
3.5. Mucosal IgA Levels in Lungs and Intestines
3.6. Splenocyte Proliferation by ex vivo Restimulation
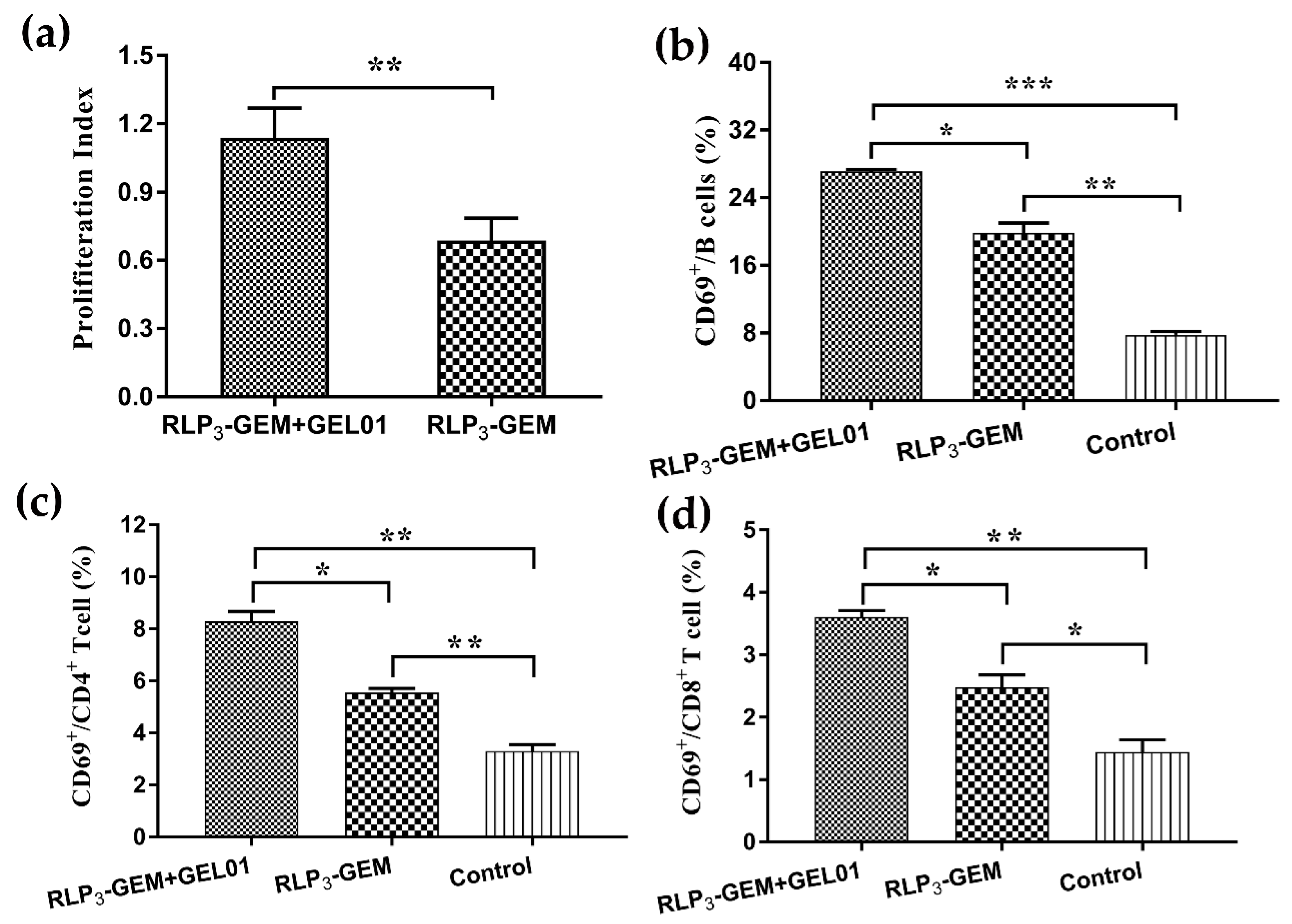
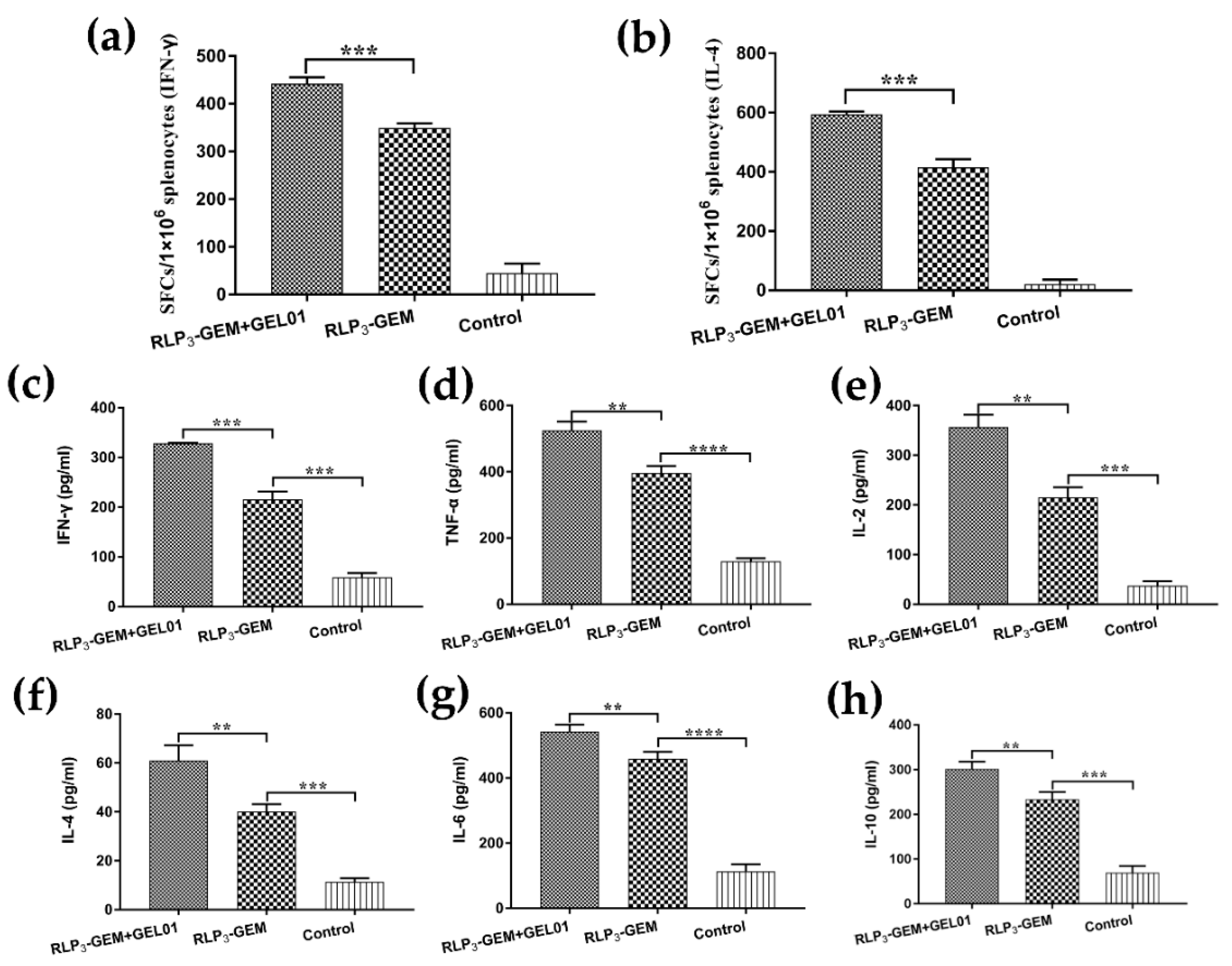
3.7. Splenocyte Activation Assays
3.8. Cytokine Secretion by Restimulated Splenocytes
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Milne-Price, S.; Miazgowicz, K.L.; Munster, V.J. The emergence of the Middle East respiratory syndrome coronavirus. Pathog. Dis. 2014, 71, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Wong, G.; Shi, W.F.; Liu, J.; Lai, A.C.K.; Zhou, J.Y.; Liu, W.J.; Bi, Y.H.; Gao, G.F. Epidemiology, genetic recombination, and pathogenesis of coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef] [PubMed]
- Middle East Respiratory Syndrome Coronavirus (Mers-Cov). Available online: https://www.Who.Int/emergencies/mers-cov/en/ (accessed on 19 July 2019).
- Ki, M. 2015 mers outbreak in Korea: Hospital-to-hospital transmission. Epidemiol. Health 2015, 37, e2015033. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Wong, G.; Liu, Y.; Gao, G.F.; Li, S.; Bi, Y. Mers in South Korea and China: A potential outbreak threat? Lancet 2015, 385, 2349–2350. [Google Scholar] [CrossRef]
- Wang, C.; Zheng, X.X.; Gai, W.W.; Wong, G.; Wang, H.L.; Jin, H.L.; Feng, N.; Zhao, Y.K.; Zhang, W.J.; Li, N.; et al. Novel chimeric virus-like particles vaccine displaying mers-cov receptor-binding domain induce specific humoral and cellular immune response in mice. Antivir. Res. 2017, 140, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Du, L.Y.; Kou, Z.H.; Ma, C.Q.; Tao, X.R.; Wang, L.L.; Zhao, G.Y.; Chen, Y.Q.; Yu, F.; Tseng, C.T.K.; Zhou, Y.S.; et al. A truncated receptor-binding domain of mers-cov spike protein potently inhibits mers-cov infection and induces strong neutralizing antibody responses: Implication for developing therapeutics and vaccines. PLoS ONE 2013, 8, e81587. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, C.; Zhao, G.Y.; Chu, H.; Wang, D.; Yan, H.H.N.; Poon, V.K.M.; Wen, L.; Wong, B.H.Y.; Zhao, X.Y.; et al. Human intestinal tract serves as an alternative infection route for middle east respiratory syndrome coronavirus. Sci. Adv. 2017, 3, eaao4966. [Google Scholar] [CrossRef] [PubMed]
- Chi, H.; Zheng, X.; Wang, X.; Wang, C.; Wang, H.; Gai, W.; Perlman, S.; Yang, S.; Zhao, J.; Xia, X. DNA vaccine encoding middle east respiratory syndrome coronavirus s1 protein induces protective immune responses in mice. Vaccine 2017, 35, 2069–2075. [Google Scholar] [CrossRef]
- Al-Amri, S.S.; Abbas, A.T.; Siddiq, L.A.; Alghamdi, A.; Sanki, M.A.; Al-Muhanna, M.K.; Alhabbab, R.Y.; Azhar, E.I.; Li, X.; Hashem, A.M. Immunogenicity of candidate mers-cov DNA vaccines based on the spike protein. Sci. Rep. 2017, 7, 44875. [Google Scholar] [CrossRef]
- Muthumani, K.; Falzarano, D.; Reuschel, E.L.; Tingey, C.; Flingai, S.; Villarreal, D.O.; Wise, M.; Patel, A.; Izmirly, A.; Aljuaid, A.; et al. A synthetic consensus anti-spike protein DNA vaccine induces protective immunity against Middle East respiratory syndrome coronavirus in nonhuman primates. Sci. Transl. Med. 2015, 7. [Google Scholar] [CrossRef]
- Jia, W.X.; Channappanavar, R.; Zhang, C.; Li, M.X.; Zhou, H.X.; Zhang, S.Y.; Zhou, P.P.; Xu, J.Y.; Shan, S.S.; Shi, X.L.; et al. Single intranasal immunization with chimpanzee adenovirus-based vaccine induces sustained and protective immunity against mers-cov infection. Emerg. Microbes Infect. 2019, 8, 760–772. [Google Scholar] [CrossRef]
- Haagmans, B.L.; van den Brand, J.M.; Raj, V.S.; Volz, A.; Wohlsein, P.; Smits, S.L.; Schipper, D.; Bestebroer, T.M.; Okba, N.; Fux, R.; et al. An orthopoxvirus-based vaccine reduces virus excretion after mers-cov infection in dromedary camels. Science 2016, 351, 77–81. [Google Scholar] [CrossRef]
- Volz, A.; Kupke, A.; Song, F.; Jany, S.; Fux, R.; Shams-Eldin, H.; Schmidt, J.; Becker, C.; Eickmann, M.; Becker, S.; et al. Protective efficacy of recombinant modified vaccinia virus ankara delivering middle east respiratory syndrome coronavirus spike glycoprotein. J. Virol. 2015, 89, 8651–8656. [Google Scholar] [CrossRef]
- Guo, X.J.; Deng, Y.; Chen, H.; Lan, J.M.; Wang, W.; Zou, X.H.; Hung, T.; Lu, Z.Z.; Tan, W.J. Systemic and mucosal immunity in mice elicited by a single immunization with human adenovirus type 5 or 41 vector-based vaccines carrying the spike protein of middle east respiratory syndrome coronavirus. Immunology 2015, 145, 476–484. [Google Scholar] [CrossRef]
- Liu, R.Q.; Ge, J.Y.; Wang, J.L.; Shao, Y.; Zhang, H.L.; Wang, J.L.; Wen, Z.Y.; Bu, Z.G. Newcastle disease virus-based mers-cov candidate vaccine elicits high-level and lasting neutralizing antibodies in bactrian camels. J. Integr. Agric. 2017, 16, 2264–2273. [Google Scholar] [CrossRef]
- Malczyk, A.H.; Kupke, A.; Prufer, S.; Scheuplein, V.A.; Hutzler, S.; Kreuz, D.; Beissert, T.; Bauer, S.; Hubich-Rau, S.; Tondera, C.; et al. A highly immunogenic and protective middle east respiratory syndrome coronavirus vaccine based on a recombinant measles virus vaccine platform. J. Virol. 2015, 89, 11654–11667. [Google Scholar] [CrossRef]
- Wirblich, C.; Coleman, C.M.; Kurup, D.; Abraham, T.S.; Bernbaum, J.G.; Jahrling, P.B.; Hensley, L.E.; Johnson, R.F.; Frieman, M.B.; Schnell, M.J. One-health: A safe, efficient, dual-use vaccine for humans and animals against middle east respiratory syndrome coronavirus and rabies virus. J. Virol. 2017, 91. [Google Scholar] [CrossRef]
- Wang, C.; Zheng, X.; Gai, W.; Zhao, Y.; Wang, H.; Wang, H.; Feng, N.; Chi, H.; Qiu, B.; Li, N.; et al. Mers-cov virus-like particles produced in insect cells induce specific humoural and cellular imminity in rhesus macaques. Oncotarget 2017, 8, 12686–12694. [Google Scholar] [CrossRef]
- Maslow, J.N. Vaccines for emerging infectious diseases: Lessons from mers coronavirus and zika virus. Hum. Vaccines Immunother. 2017, 13, 2918–2930. [Google Scholar] [CrossRef]
- Zhang, N.R.; Jiang, S.B.; Du, L.Y. Current advancements and potential strategies in the development of mers-cov vaccines. Expert Rev. Vaccines 2014, 13, 761–774. [Google Scholar] [CrossRef]
- Du, L.Y.; Zhao, G.Y.; Kou, Z.H.; Ma, C.Q.; Sun, S.H.; Poon, V.K.M.; Lu, L.; Wang, L.L.; Debnath, A.K.; Zheng, B.J.; et al. Identification of a receptor-binding domain in the s protein of the novel human coronavirus middle east respiratory syndrome coronavirus as an essential target for vaccine development. J. Virol. 2013, 87, 11963. [Google Scholar] [CrossRef]
- Zhang, N.R.; Tang, J.; Lu, L.; Jiang, S.B.; Du, L.Y. Receptor-binding domain-based subunit vaccines against mers-cov. Virus Res. 2015, 202, 151–159. [Google Scholar] [CrossRef]
- Lan, J.M.; Deng, Y.; Chen, H.; Lu, G.W.; Wang, W.; Guo, X.J.; Lu, Z.Z.; Gao, G.F.; Tan, W.J. Tailoring subunit vaccine immunity with adjuvant combinations and delivery routes using the middle east respiratory coronavirus (mers-cov) receptor-binding domain as an antigen. PLoS ONE 2014, 9, e112602. [Google Scholar] [CrossRef]
- Raj, V.S.; Mou, H.; Smits, S.L.; Dekkers, D.H.; Muller, M.A.; Dijkman, R.; Muth, D.; Demmers, J.A.; Zaki, A.; Fouchier, R.A.; et al. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-emc. Nature 2013, 495, 251–254. [Google Scholar] [CrossRef]
- Mou, H.; Raj, V.S.; van Kuppeveld, F.J.; Rottier, P.J.; Haagmans, B.L.; Bosch, B.J. The receptor binding domain of the new middle east respiratory syndrome coronavirus maps to a 231-residue region in the spike protein that efficiently elicits neutralizing antibodies. J. Virol. 2013, 87, 9379–9383. [Google Scholar] [CrossRef]
- Ma, C.Q.; Wang, L.L.; Tao, X.R.; Zhang, N.R.; Yang, Y.; Tseng, C.T.K.; Li, F.; Zhou, Y.S.; Jiang, S.B.; Du, L.Y. Searching for an ideal vaccine candidate among different mers coronavirus receptor-binding fragments-the importance of immunofocusing in subunit vaccine design. Vaccine 2014, 32, 6170–6176. [Google Scholar] [CrossRef]
- Van Roosmalen, M.L.; Kanninga, R.; El Khattabi, M.; Neef, J.; Audouy, S.; Bosma, T.; Kuipers, A.; Post, E.; Steen, A.; Kok, J.; et al. Mucosal vaccine delivery of antigens tightly bound to an adjuvant particle made from food-grade bacteria. Methods 2006, 38, 144–149. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Choi, J.H.; Xu, Z.H. Microbial cell-surface display. Trends Biotechnol. 2003, 21, 45–52. [Google Scholar] [CrossRef]
- Bosma, T.; Kanninga, R.; Neef, J.; Audouy, S.A.L.; van Roosmalen, M.L.; Steen, A.; Buist, G.; Kok, J.; Kuipers, O.P.; Robillard, G.; et al. Novel surface display system for proteins on non-genetically modified Gram-positive bacteria. Appl. Environ. Microb. 2006, 72, 880–889. [Google Scholar] [CrossRef]
- Ramasamy, R.; Yasawardena, S.; Zomer, A.; Venema, G.; Kok, J.; Leenhouts, K. Immunogenicity of a malaria parasite antigen displayed by lactococcus lactis in oral immunisations. Vaccine 2006, 24, 3900–3908. [Google Scholar] [CrossRef]
- Audouy, S.A.L.; van Selm, S.; van Roosmalen, M.L.; Post, E.; Kanninga, R.; Neef, J.; Estevao, S.; Nieuwenhuis, E.E.S.; Adrian, P.V.; Leenhouts, K.; et al. Development of lactococcal gem-based pneumococcal vaccines. Vaccine 2007, 25, 2497–2506. [Google Scholar] [CrossRef]
- Li, P.C.; Qiao, X.W.; Zheng, Q.S.; Hou, J.B. Immunogenicity and immunoprotection of porcine circovirus type 2 (pcv2) cap protein displayed by lactococcus lactis. Vaccine 2016, 34, 696–702. [Google Scholar] [CrossRef]
- Buist, G.; Steen, A.; Kok, J.; Kuipers, O.R. Lysm, a widely distributed protein motif for binding to (peptido)glycans. Mol. Microbiol. 2008, 68, 838–847. [Google Scholar] [CrossRef]
- Michon, C.; Langella, P.; Eijsink, V.G.H.; Mathiesen, G.; Chatel, J.M. Display of recombinant proteins at the surface of lactic acid bacteria: Strategies and applications. Microb. Cell Fact. 2016, 15, 70. [Google Scholar] [CrossRef]
- Steen, A.; Buist, G.; Horsburgh, G.J.; Venema, G.; Kuipers, O.P.; Foster, S.J.; Kok, J. Acma of lactococcus lactis is an n-acetylglucosaminidase with an optimal number of lysm domains for proper functioning. FEBS J. 2005, 272, 2854–2868. [Google Scholar] [CrossRef]
- Qiao, X.; Li, P.; Zheng, Q.; Chen, J.; Yu, X.; Hou, L.; Wu, N.; Hou, J. Comparison of the binding activity of lactococcus lactis peptidoglycan protein anchor with different number of motifs. Wei Sheng Wu Xue Bao = Acta Microbiol. Sin. 2015, 55, 193–197. [Google Scholar]
- Li, Y.; Wan, Y.H.; Liu, P.P.; Zhao, J.C.; Lu, G.W.; Qi, J.X.; Wang, Q.H.; Lu, X.C.; Wu, Y.; Liu, W.J.; et al. A humanized neutralizing antibody against mers-cov targeting the receptor-binding domain of the spike protein. Cell Res. 2015, 25, 1237–1249. [Google Scholar] [CrossRef]
- Zhang, W.F.; Wang, L.Y.; Liu, Y.; Chen, X.M.; Liu, Q.; Jia, J.L.; Yang, T.Y.; Qiu, S.H.; Ma, G.H. Immune responses to vaccines involving a combined antigen-nanoparticle mixture and nanoparticle-encapsulated antigen formulation. Biomaterials 2014, 35, 6086–6097. [Google Scholar] [CrossRef]
- Lindsey, W.B.; Lowdell, M.W.; Marti, G.E.; Abbasi, F.; Zenger, V.; King, K.M.; Lamb, L.S., Jr. Cd69 expression as an index of t-cell function: Assay standardization, validation and use in monitoring immune recovery. Cytotherapy 2007, 9, 123–132. [Google Scholar] [CrossRef]
- Assiri, A. Hospital outbreak of middle east respiratory syndrome coronavirus. N. Engl. J. Med. 2013, 369, 886. [Google Scholar] [CrossRef]
- Ma, C.Q.; Li, Y.; Wang, L.L.; Zhao, G.Y.; Tao, X.R.; Tseng, C.T.K.; Zhou, Y.S.; Du, L.Y.; Jiang, S.B. Intranasal vaccination with recombinant receptor-binding domain of mers-cov spike protein induces much stronger local mucosal immune responses than subcutaneous immunization: Implication for designing novel mucosal mers vaccines. Vaccine 2014, 32, 2100–2108. [Google Scholar] [CrossRef]
- Salminen, S.; von Wright, A.; Morelli, L.; Marteau, P.; Brassart, D.; de Vos, W.M.; Fonden, R.; Saxelin, M.; Collins, K.; Mogensen, G.; et al. Demonstration of safety of probiotics—A review. Int. J. Food Microbiol. 1998, 44, 93–106. [Google Scholar] [CrossRef]
- Nganou-Makamdop, K.; van Roosmalen, M.L.; Audouy, S.A.L.; van Gemert, G.J.; Leenhouts, K.; Hermsen, C.C.; Sauerwein, R.W. Bacterium-like particles as multi-epitope delivery platform for plasmodium berghei circumsporozoite protein induce complete protection against malaria in mice. Malar. J. 2012, 11, 50. [Google Scholar] [CrossRef]
- Li, B.; Chen, X.R.; Yu, J.F.; Zhang, Y.; Mo, Z.J.; Gu, T.J.; Kong, W.; Wu, Y.G. Protection elicited by nasal immunization with pneumococcal surface protein a (pspa) adjuvanted with bacterium-like particles against streptococcus pneumoniae infection in mice. Microb. Pathog. 2018, 123, 115–119. [Google Scholar] [CrossRef]
- Heine, S.J.; Franco-Mahecha, O.L.; Chen, X.T.; Choudhari, S.; Blackwelder, W.C.; van Roosmalen, M.L.; Leenhouts, K.; Picking, W.L.; Pasetti, M.F. Shigella ipab and ipad displayed on l-lactis bacterium-like particles induce protective immunity in adult and infant mice. Immunol. Cell Biol. 2015, 93, 641–652. [Google Scholar] [CrossRef]
- Ramirez, K.; Ditamo, Y.; Rodriguez, L.; Picking, W.L.; van Roosmalen, M.L.; Leenhouts, K.; Pasetti, M.F. Neonatal mucosal immunization with a non-living, non-genetically modified lactococcus lactis vaccine carrier induces systemic and local th1-type immunity and protects against lethal bacterial infection. Mucosal. Immunol. 2010, 3, 159–171. [Google Scholar] [CrossRef]
- Rigter, A.; Widjaja, I.; Versantvoort, H.; Coenjaerts, F.E.J.; Van Roosmalen, M.; Leenhouts, K.; Rottier, P.J.M.; Haijema, B.J.; de Haan, C.A.M. A protective and safe intranasal rsv vaccine based on a recombinant prefusion-like form of the f protein bound to bacterium-like particles. PLoS ONE 2013, 8, e71072. [Google Scholar] [CrossRef]
- Bateman, A.; Bycroft, M. The structure of a lysm domain from e-coli membrane-bound lytic murein transglycosylase d (mltd). J. Mol. Biol. 2000, 299, 1113–1119. [Google Scholar] [CrossRef]
- Okano, K.; Zhang, Q.; Kimura, S.; Narita, J.; Tanaka, T.; Fukuda, H.; Kondo, A. System using tandem repeats of the ca peptidoglycan-binding domain from lactococcus lactis for display of both n- and c-terminal fusions on cell surfaces of lactic acid bacteria. Appl. Environ. Microb. 2008, 74, 1117–1123. [Google Scholar] [CrossRef]
- Chen, X.Y.; Zaro, J.L.; Shen, W.C. Fusion protein linkers: Property, design and functionality. Adv. Drug Deliv. Rev. 2013, 65, 1357–1369. [Google Scholar] [CrossRef]
- Deville, S.; Ben Arous, J.; Bertrand, F.; Borisov, V.; Dupuis, L. Efficacy of intranasal and spray delivery of adjuvanted live vaccine against infectious bronchitis virus in experimentally infected poultry. Procedia Vaccinol. 2012, 6, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Nimmerjahn, F.; Ravetch, J.V. Divergent immunoglobulin g subclass activity through selective fc receptor binding. Science 2005, 310, 1510–1512. [Google Scholar] [CrossRef] [PubMed]
- Sancho, D.; Gomez, M.; Sanchez-Madrid, F. Cd69 is an immunoregulatory molecule induced following activation. Trends Immunol. 2005, 26, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Vialle, R.; Dupuis, L.; Deville, S.; Bertrand, F.; Gaucheron, J.; Aucouturier, J. Microgel particulate adjuvant: Characterisation and mechanisms of action. Procedia Vaccinol. 2010, 2, 12–16. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligonucleotide Primers | Sequences |
|---|---|
| RBD-F1,3 | 5′-TGCTCTAGACATCACCATCACCATCACCAAGCCGAAGGAGTTGAA-3′ (XbaI) |
| Linker-RBD-R2 | 5′-ACCAGAACCACCACCAGAACCACCCAACTTAGGGCAGACGCT-3′ |
| RBD-R | 5′-GTTACCAGCTGAAGAAGCACCATCCAACTTAGGGCAGACGCT-3′ |
| PA3-F | 5′-ACCAATAGCGTCTGCCCTAAGTTGGATGGTGCTTCTTCAGCTGG-3′ |
| Linker-PA2-F2 | 5′-GGTGGTTCTGGTGGTGGTTCTGGTACTACCGTTAAGGTGAAGTC-3′ |
| Linker-PA3-F2 | 5′-GGTGGTTCTGGTGGTGGTTCTGGTGATGGTGCTTCTTCAGCTGG-3′ |
| PA3-R1 | 5′-CGGGGTACCTTACTTGATACGCAGGTATTGAC-3′ (KpnI) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, E.; Chi, H.; Huang, P.; Yan, F.; Zhang, Y.; Liu, C.; Wang, Z.; Li, G.; Zhang, S.; Mo, R.; et al. A Novel Bacterium-Like Particle Vaccine Displaying the MERS-CoV Receptor-Binding Domain Induces Specific Mucosal and Systemic Immune Responses in Mice. Viruses 2019, 11, 799. https://doi.org/10.3390/v11090799
Li E, Chi H, Huang P, Yan F, Zhang Y, Liu C, Wang Z, Li G, Zhang S, Mo R, et al. A Novel Bacterium-Like Particle Vaccine Displaying the MERS-CoV Receptor-Binding Domain Induces Specific Mucosal and Systemic Immune Responses in Mice. Viruses. 2019; 11(9):799. https://doi.org/10.3390/v11090799
Chicago/Turabian StyleLi, Entao, Hang Chi, Pei Huang, Feihu Yan, Ying Zhang, Chuanyu Liu, Zhenshan Wang, Guohua Li, Shengnan Zhang, Ruo Mo, and et al. 2019. "A Novel Bacterium-Like Particle Vaccine Displaying the MERS-CoV Receptor-Binding Domain Induces Specific Mucosal and Systemic Immune Responses in Mice" Viruses 11, no. 9: 799. https://doi.org/10.3390/v11090799