Mayaro Virus Infects Human Chondrocytes and Induces the Expression of Arthritis-Related Genes Associated with Joint Degradation


 , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Virus and Cell Lines
2.2. Primary Human Cells
2.3. Infection of Human Fibroblast-Like Synoviocytes (HFLS), Human Osteoblasts (HOB) and Human Chondrocytes (HC)
2.4. Virus Titration
2.5. Quantification of Mayaro Virus (MAYV) by Real-Time Polymerase Chain Reaction (PCR)
2.6. RT2 Profiler PCR Array
2.7. Gene Expression Analysis by Real-Time PCR
2.8. Measurements of Cytokines
2.9. Western Blot
2.10. Cell Viability Assay
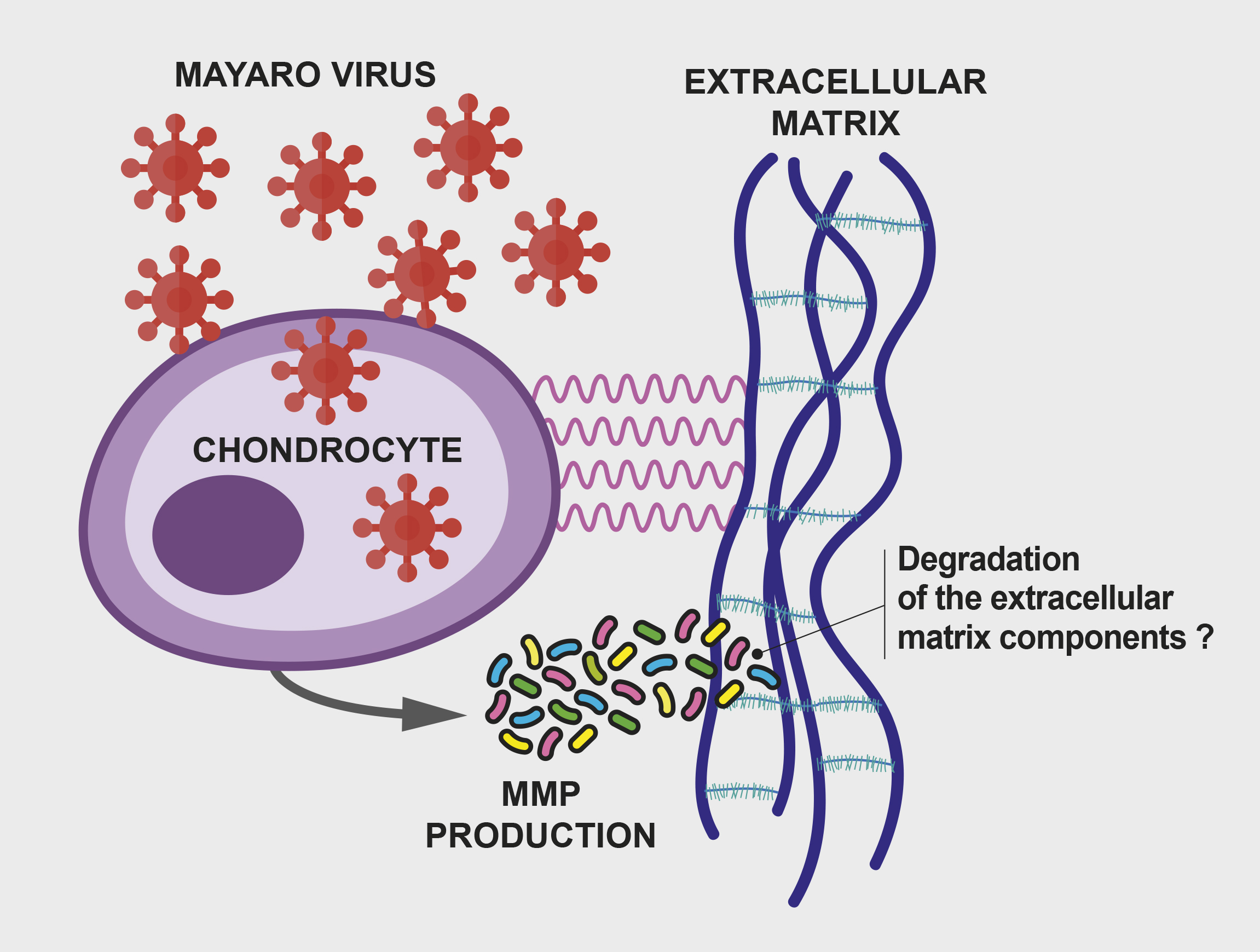
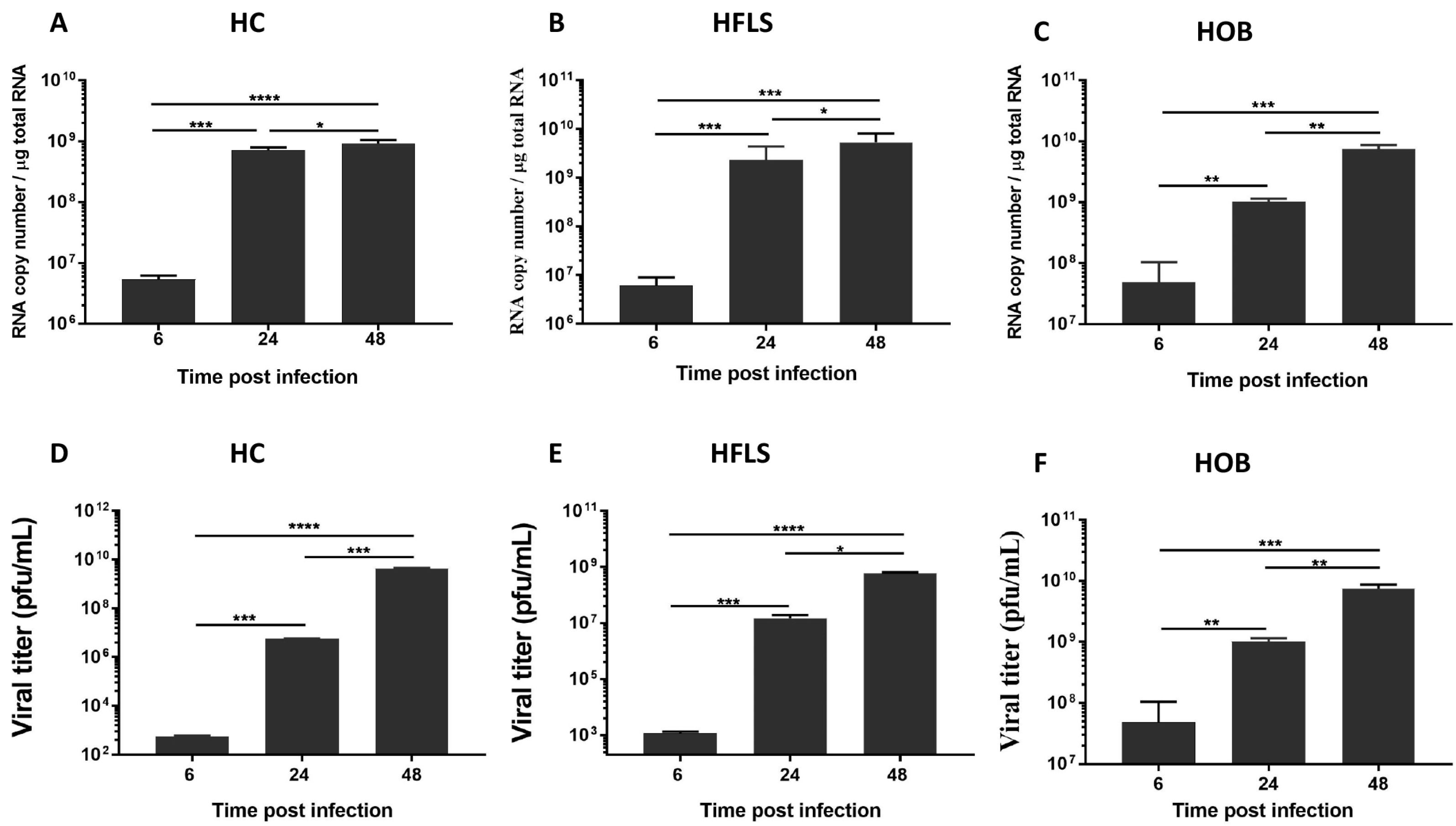
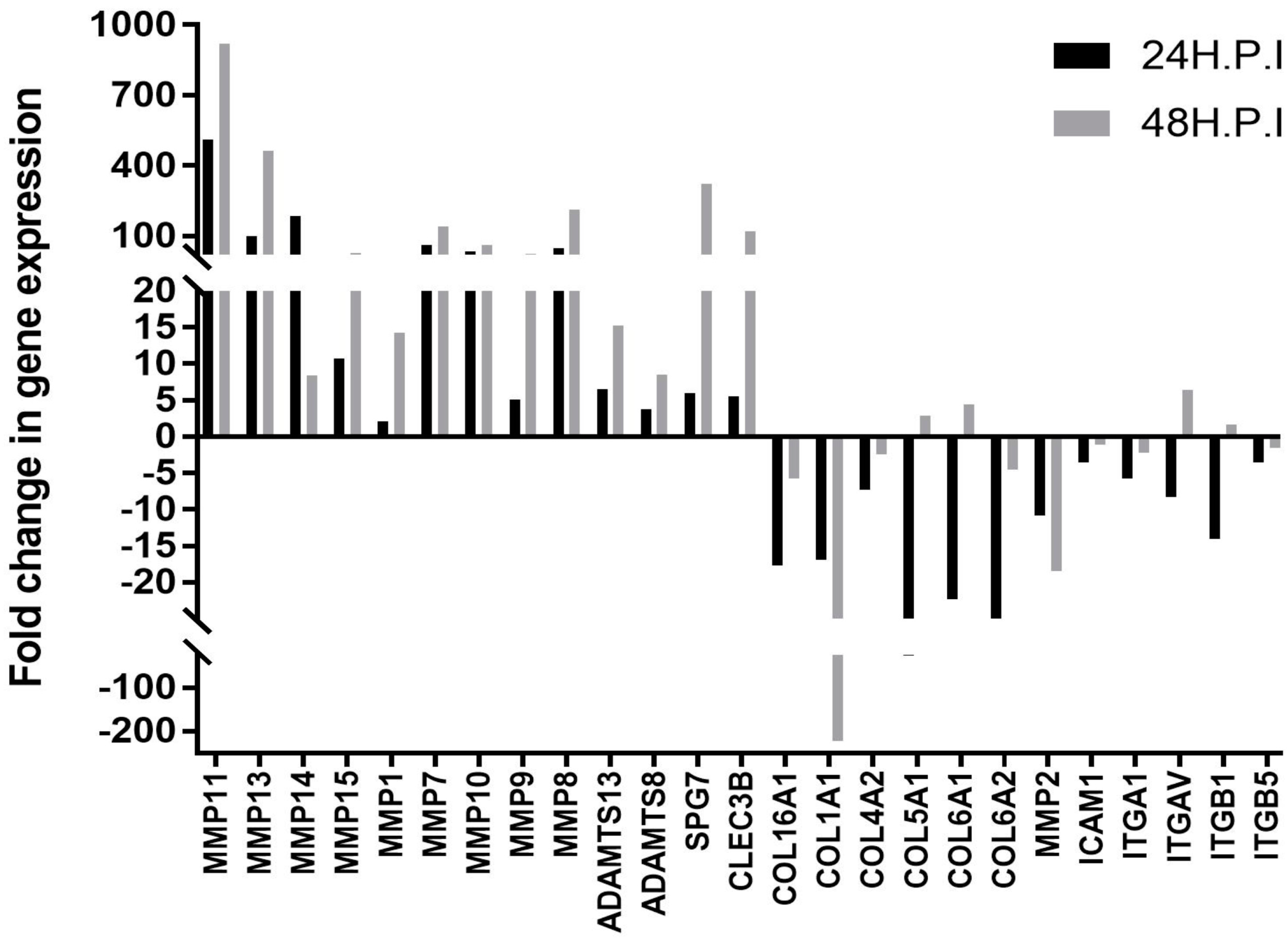
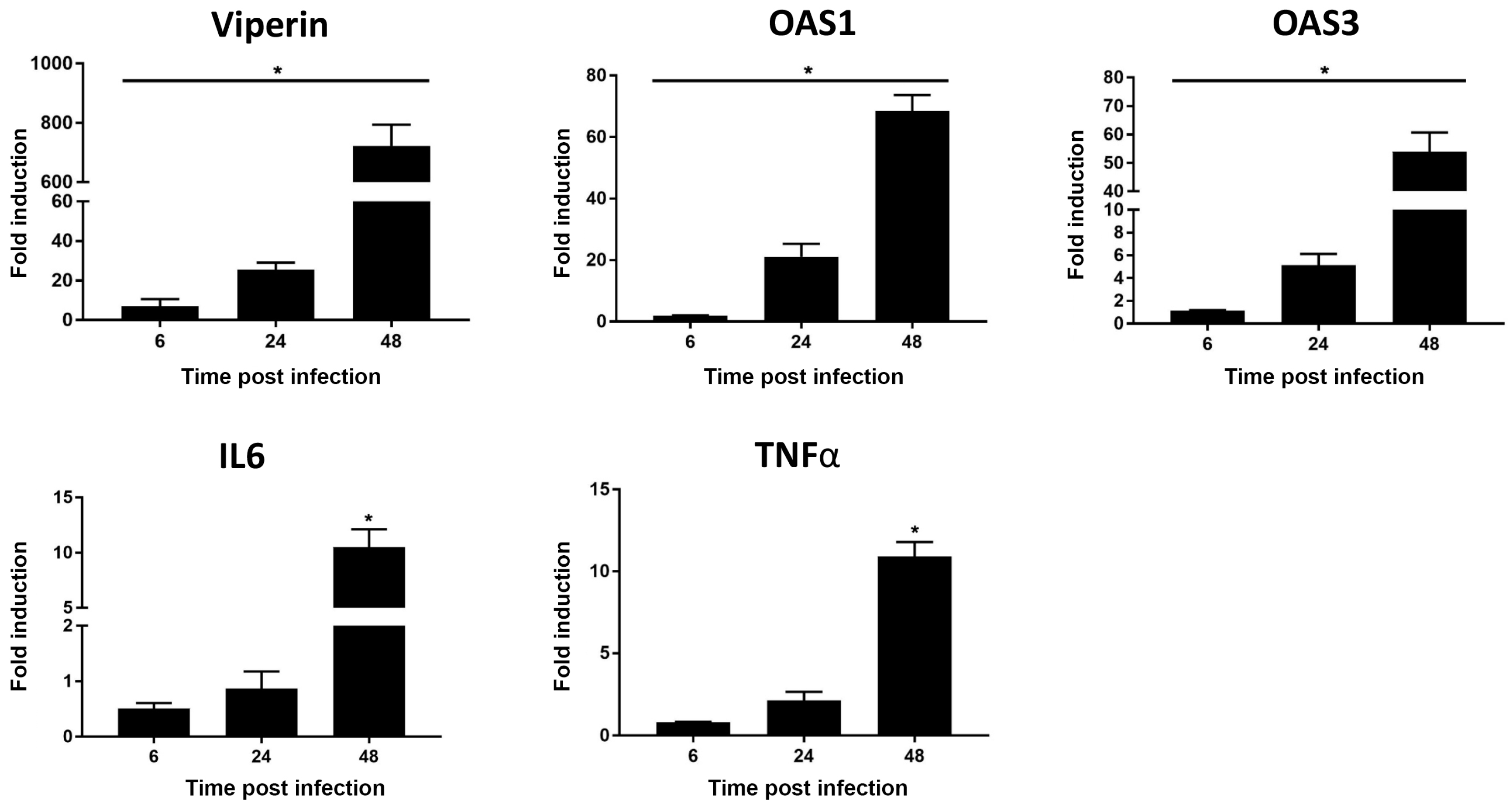
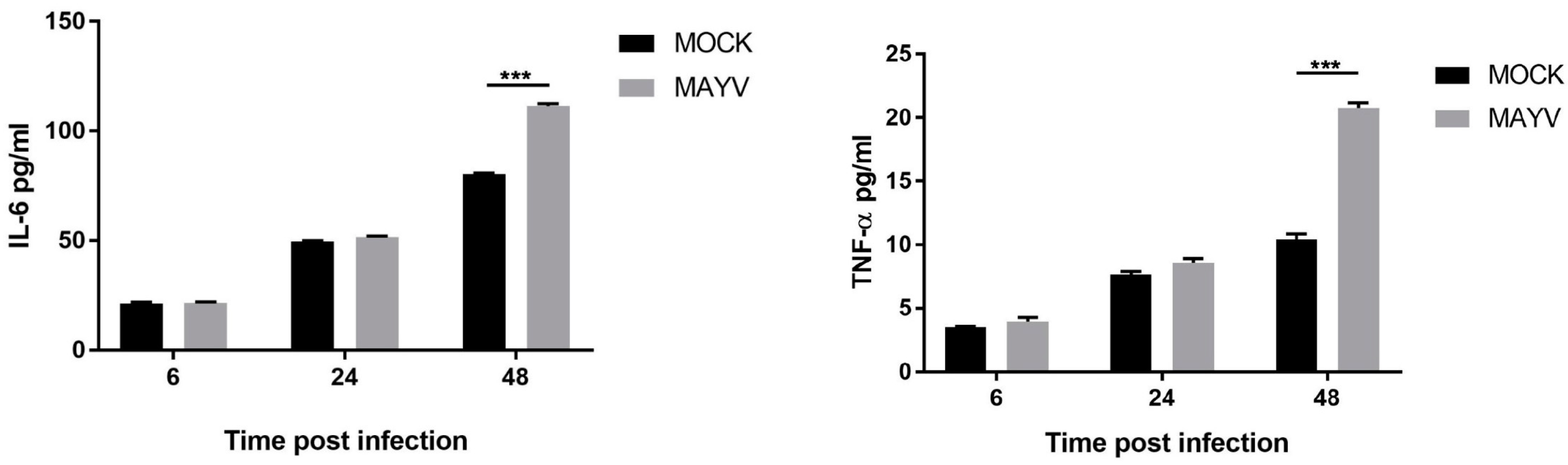
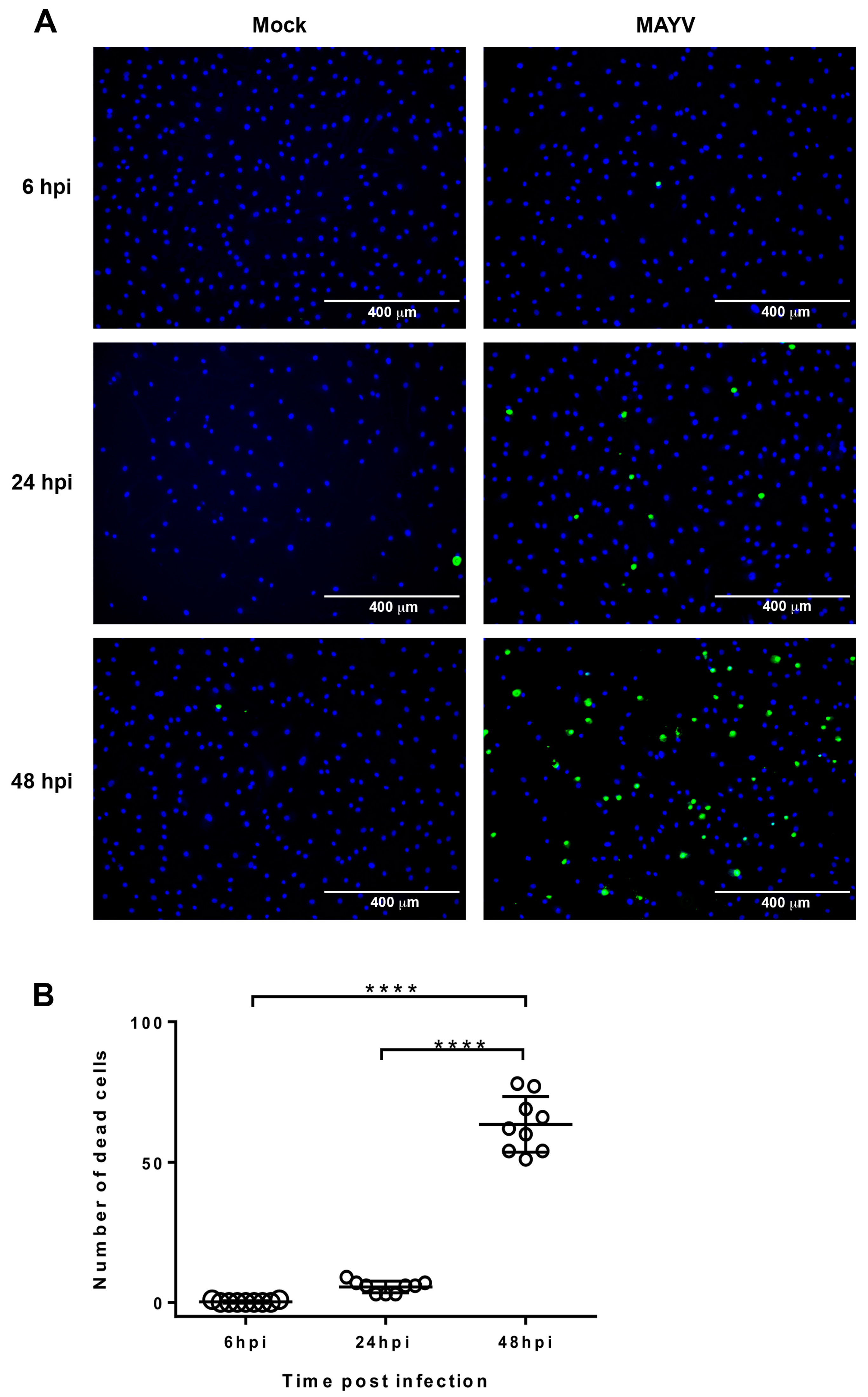
3. Results and Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Izurieta, R.O.; DeLacure, D.A.; Izurieta, A.; Hoare, I.A.; Ortiz, M.R. Mayaro virus: The jungle flu. Virus Adapt. Treat. 2018, 10, 9. [Google Scholar] [CrossRef]
- Auguste, A.J.; Liria, J.; Forrester, N.L.; Giambalvo, D.; Moncada, M.; Long, K.C.; Moron, D.; de Manzione, N.; Tesh, R.B.; Halsey, E.S.; et al. Evolutionary and ecological characterization of mayaro virus strains isolated during an outbreak, Venezuela, 2010. Emerg. Infect. Dis. 2015, 21, 1742–1750. [Google Scholar] [CrossRef] [PubMed]
- Mourao, M.P.; Bastos Mde, S.; de Figueiredo, R.P.; Gimaque, J.B.; Galusso Edos, S.; Kramer, V.M.; de Oliveira, C.M.; Naveca, F.G.; Figueiredo, L.T. Mayaro fever in the city of Manaus, Brazil, 2007–2008. Vector Borne Zoonotic Dis. 2012, 12, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Neumayr, A.; Gabriel, M.; Fritz, J.; Günther, S.; Hatz, C.; Schmidt-Chanasit, J.; Blum, J. Mayaro virus infection in traveler returning from Amazon basin, Northern Peru. Emerg. Infect. Dis. 2012, 18, 695. [Google Scholar] [CrossRef] [PubMed]
- Hotez, P.J.; Murray, K.O. Dengue, west nile virus, chikungunya, zika-and now mayaro? PLoS Negl. Trop. Dis. 2017, 11, e0005462. [Google Scholar] [CrossRef] [PubMed]
- Mavian, C.; Rife, B.D.; Dollar, J.J.; Cella, E.; Ciccozzi, M.; Prosperi, M.C.F.; Lednicky, J.; Morris, J.G.; Capua, I.; Salemi, M. Emergence of recombinant mayaro virus strains from the Amazon basin. Sci. Rep. 2017, 7, 8718. [Google Scholar] [CrossRef] [PubMed]
- Halsey, E.S.; Siles, C.; Guevara, C.; Vilcarromero, S.; Jhonston, E.J.; Ramal, C.; Aguilar, P.V.; Ampuero, J.S. Mayaro virus infection, Amazon basin region, Peru, 2010–2013. Emerg. Infect. Dis. 2013, 19, 1839. [Google Scholar] [CrossRef] [PubMed]
- Morrison, T.E.; Whitmore, A.C.; Shabman, R.S.; Lidbury, B.A.; Mahalingam, S.; Heise, M.T. Characterization of ross river virus tropism and virus-induced inflammation in a mouse model of viral arthritis and myositis. J. Virol. 2006, 80, 737–749. [Google Scholar] [CrossRef]
- Morrison, T.E.; Oko, L.; Montgomery, S.A.; Whitmore, A.C.; Lotstein, A.R.; Gunn, B.M.; Elmore, S.A.; Heise, M.T. A mouse model of chikungunya virus–induced musculoskeletal inflammatory disease: Evidence of arthritis, tenosynovitis, myositis, and persistence. Am. J. Pathol. 2011, 178, 32–40. [Google Scholar] [CrossRef]
- Santos, F.M.; Dias, R.S.; de Oliveira, M.D.; Costa, I.C.T.A.; de Souza Fernandes, L.; Pessoa, C.R.; da Matta, S.L.P.; Costa, V.V.; Souza, D.G.; da Silva, C.C. Animal model of arthritis and myositis induced by the mayaro virus. PLoS Negl. Trop. Dis. 2019, 13, e0007375. [Google Scholar] [CrossRef]
- Ozden, S.; Huerre, M.; Riviere, J.-P.; Coffey, L.L.; Afonso, P.V.; Mouly, V.; De Monredon, J.; Roger, J.-C.; El Amrani, M.; Yvin, J.-L. Human muscle satellite cells as targets of chikungunya virus infection. PLoS ONE 2007, 2, e527. [Google Scholar] [CrossRef] [PubMed]
- Rohatgi, A.; Corbo, J.C.; Monte, K.; Higgs, S.; Vanlandingham, D.L.; Kardon, G.; Lenschow, D.J. Infection of myofibers contributes to increased pathogenicity during infection with an epidemic strain of chikungunya virus. J. Virol. 2014, 88, 2414–2425. [Google Scholar] [CrossRef] [PubMed]
- Santiago, F.W.; Halsey, E.S.; Siles, C.; Vilcarromero, S.; Guevara, C.; Silvas, J.A.; Ramal, C.; Ampuero, J.S.; Aguilar, P.V. Long-term arthralgia after mayaro virus infection correlates with sustained pro-inflammatory cytokine response. PLoS Negl. Trop. Dis. 2015, 9, e0004104. [Google Scholar] [CrossRef] [PubMed]
- Hoarau, J.-J.; Bandjee, M.-C.J.; Trotot, P.K.; Das, T.; Li-Pat-Yuen, G.; Dassa, B.; Denizot, M.; Guichard, E.; Ribera, A.; Henni, T. Persistent chronic inflammation and infection by chikungunya arthritogenic alphavirus in spite of a robust host immune response. J. Immunol. 2010, 184, 5914–5927. [Google Scholar] [CrossRef] [PubMed]
- Mylonas, A.D.; Brown, A.M.; Carthew, T.L.; Purdie, D.M.; Pandeya, N.; Collins, L.G.; Suhrbier, A.; McGrath, B.; Reymond, E.J.; Vecchio, P.C. Natural history of ross river virus-induced epidemic polyarthritis. Med. J. Aust. 2002, 177, 356–360. [Google Scholar] [PubMed]
- Tesh, R.B.; Watts, D.M.; Russell, K.L.; Damodaran, C.; Calampa, C.; Cabezas, C.; Ramirez, G.; Vasquez, B.; Hayes, C.G.; Rossi, C.A.; et al. Mayaro virus disease: An emerging mosquito-borne zoonosis in tropical South America. Clin. Infect. Dis. 1999, 28, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Kurkela, S.; Manni, T.; Myllynen, J.; Vaheri, A.; Vapalahti, O. Clinical and laboratory manifestations of sindbis virus infection: Prospective study, finland, 2002–2003. J. Infect. Dis. 2005, 191, 1820–1829. [Google Scholar] [CrossRef]
- Kurkela, S.; Helve, T.; Vaheri, A.; Vapalahti, O. Arthritis and arthralgia three years after sindbis virus infection: Clinical follow-up of a cohort of 49 patients. Scand. J. Infect. Dis. 2008, 40, 167–173. [Google Scholar] [CrossRef]
- Chen, W.; Foo, S.-S.; Rulli, N.E.; Taylor, A.; Sheng, K.-C.; Herrero, L.J.; Herring, B.L.; Lidbury, B.A.; Li, R.W.; Walsh, N.C. Arthritogenic alphaviral infection perturbs osteoblast function and triggers pathologic bone loss. Proc. Natl. Acad. Sci. USA 2014, 111, 6040–6045. [Google Scholar] [CrossRef] [Green Version]
- Lim, E.; Supramaniam, A.; Lui, H.; Coles, P.; Lee, W.; Liu, X.; Rudd, P.; Herrero, L. Chondrocytes contribute to alphaviral disease pathogenesis as a source of virus replication and soluble factor production. Viruses 2018, 10, 86. [Google Scholar] [CrossRef]
- Soden, M.; Vasudevan, H.; Roberts, B.; Coelen, R.; Hamlin, G.; Vasudevan, S.; La Brooy, J. Detection of viral ribonucleic acid and histologic analysis of inflamed synovium in ross river virus infection. Arthritis Rheum. J. Am. Coll. Rheumatol. 2000, 43, 365–369. [Google Scholar] [CrossRef]
- Lednicky, J.; De Rochars, V.M.; Elbadry, M.; Loeb, J.; Telisma, T.; Chavannes, S.; Anilis, G.; Cella, E.; Ciccozzi, M.; Okech, B.; et al. Mayaro virus in child with acute febrile illness, haiti, 2015. Emerg. Infect. Dis. 2016, 22, 2000–2002. [Google Scholar] [CrossRef] [PubMed]
- Atieh, T.; Baronti, C.; De Lamballerie, X.; Nougairède, A. Simple reverse genetics systems for asian and african zika viruses. Sci. Rep. 2016, 6, 39384. [Google Scholar] [CrossRef] [PubMed]
- Aubry, F.; Nougairede, A.; de Fabritus, L.; Querat, G.; Gould, E.A.; de Lamballerie, X. Single-stranded positive-sense rna viruses generated in days using infectious subgenomic amplicons. J. Gen. Virol. 2014, 95, 2462. [Google Scholar] [CrossRef] [PubMed]
- Levi, L.I.; Vignuzzi, M. Arthritogenic alphaviruses: A worldwide emerging threat? Microorganisms 2019, 7, 133. [Google Scholar] [CrossRef] [PubMed]
- Mostafavi, H.; Abeyratne, E.; Zaid, A.; Taylor, A. Arthritogenic alphavirus-induced immunopathology and targeting host inflammation as a therapeutic strategy for alphaviral disease. Viruses 2019, 11, 290. [Google Scholar] [CrossRef] [PubMed]
- Phuklia, W.; Kasisith, J.; Modhiran, N.; Rodpai, E.; Thannagith, M.; Thongsakulprasert, T.; Smith, D.R.; Ubol, S. Osteoclastogenesis induced by chikv-infected fibroblast-like synoviocytes: A possible interplay between synoviocytes and monocytes/macrophages in chikv-induced arthralgia/arthritis. Virus Res. 2013, 177, 179–188. [Google Scholar] [CrossRef]
- Mobasheri, A.; Bay-Jensen, A.-C.; Gualillo, O.; Larkin, J.; Levesque, M.C.; Henrotin, Y. Soluble biochemical markers of osteoarthritis: Are we close to using them in clinical practice? Best Pract. Res. Clin. Rheumatol. 2018, 19, 199–208. [Google Scholar] [CrossRef]
- Archer, C.W.; Francis-West, P. The chondrocyte. Int. J. Biochem. Cell Biol. 2003, 35, 401–404. [Google Scholar] [CrossRef]
- Callahan, L.A.S.; Ganios, A.M.; Childers, E.P.; Weiner, S.D.; Becker, M.L. Primary human chondrocyte extracellular matrix formation and phenotype maintenance using rgd-derivatized pegdm hydrogels possessing a continuous young’s modulus gradient. Acta Biomater. 2013, 9, 6095–6104. [Google Scholar] [CrossRef]
- Herrero, L.J.; Foo, S.-S.; Sheng, K.-C.; Chen, W.; Forwood, M.R.; Bucala, R.; Mahalingam, S. Pentosan polysulfate: A novel glycosaminoglycan-like molecule for effective treatment of alphavirus-induced cartilage destruction and inflammatory disease. J. Virol. 2015, 89, 8063–8076. [Google Scholar] [CrossRef] [PubMed]
- Murphy, G.; Knäuper, V.; Atkinson, S.; Butler, G.; English, W.; Hutton, M.; Stracke, J.; Clark, I. Matrix metalloproteinases in arthritic disease. Arthritis Res. Ther. 2002, 4, S39. [Google Scholar] [CrossRef] [PubMed]
- Hotary, K.; Allen, E.; Punturieri, A.; Yana, I.; Weiss, S.J. Regulation of cell invasion and morphogenesis in a three-dimensional type i collagen matrix by membrane-type matrix metalloproteinases 1, 2, and 3. J. Cell Biol. 2000, 149, 1309–1323. [Google Scholar] [CrossRef]
- Valdes, A.M.; Hart, D.J.; Jones, K.A.; Surdulescu, G.; Swarbrick, P.; Doyle, D.V.; Schafer, A.J.; Spector, T.D. Association study of candidate genes for the prevalence and progression of knee osteoarthritis. Arthritis Rheum. J. Am. Coll. Rheumatol. 2004, 50, 2497–2507. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, C.; Dehne, T.; Lindahl, A.; Brittberg, M.; Pruss, A.; Sittinger, M.; Ringe, J. Genome-wide expression profiling reveals new candidate genes associated with osteoarthritis. Osteoarthr. Cartil. 2010, 18, 581–592. [Google Scholar] [CrossRef] [Green Version]
- Ekchariyawat, P.; Hamel, R.; Bernard, E.; Wichit, S.; Surasombatpattana, P.; Talignani, L.; Thomas, F.; Choumet, V.; Yssel, H.; Despres, P.; et al. Inflammasome signaling pathways exert antiviral effect against chikungunya virus in human dermal fibroblasts. Infect. Genet. Evol. 2015, 32, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Hamel, R.; Dejarnac, O.; Wichit, S.; Ekchariyawat, P.; Neyret, A.; Luplertlop, N.; Perera-Lecoin, M.; Surasombatpattana, P.; Talignani, L.; Thomas, F.; et al. Biology of zika virus infection in human skin cells. J. Virol. 2015, 89, 8880–8896. [Google Scholar] [CrossRef]
- Carissimo, G.; Teo, T.-H.; Chan, Y.-H.; Lee, C.Y.-P.; Lee, B.; Torres-Ruesta, A.; Tan, J.J.; Chua, T.-K.; Fong, S.-W.; Lum, F.-M. Viperin controls chikungunya virus–specific pathogenic t cell ifnγ th1 stimulation in mice. Life Sci. Alliance 2019, 2, e201900298. [Google Scholar] [CrossRef]
- Bréhin, A.-C.; Casadémont, I.; Frenkiel, M.-P.; Julier, C.; Sakuntabhai, A.; Desprès, P. The large form of human 2′, 5′-oligoadenylate synthetase (oas3) exerts antiviral effect against chikungunya virus. Virology 2009, 384, 216–222. [Google Scholar] [CrossRef]
- Teng, T.-S.; Foo, S.-S.; Simamarta, D.; Lum, F.-M.; Teo, T.-H.; Lulla, A.; Yeo, N.K.; Koh, E.G.; Chow, A.; Leo, Y.-S. Viperin restricts chikungunya virus replication and pathology. J. Clin. Investig. 2012, 122, 4447–4460. [Google Scholar] [CrossRef] [Green Version]
- Malemud, C.J. Matrix metalloproteinases and synovial joint pathology. In Progress in Molecular Biology and Translational Science; Elsevier: Amsterdam, The Netherlands, 2017; Volume 148, pp. 305–325. [Google Scholar]
- Charlier, E.; Relic, B.; Deroyer, C.; Malaise, O.; Neuville, S.; Collée, J.; Malaise, M.; De Seny, D. Insights on molecular mechanisms of chondrocytes death in osteoarthritis. Int. J. Mol. Sci. 2016, 17, 2146. [Google Scholar] [CrossRef] [PubMed]
- Goldring, M.B.; Otero, M.; Plumb, D.A.; Dragomir, C.; Favero, M.; El Hachem, K.; Hashimoto, K.; Roach, H.I.; Olivotto, E.; Borzì, R.M. Roles of inflammatory and anabolic cytokines in cartilage metabolism: Signals and multiple effectors converge upon mmp-13 regulation in osteoarthritis. Eur. Cells Mater. 2011, 21, 202. [Google Scholar] [CrossRef]
- Chow, A.; Her, Z.; Ong, E.K.; Chen, J.-M.; Dimatatac, F.; Kwek, D.J.; Barkham, T.; Yang, H.; Rénia, L.; Leo, Y.-S. Persistent arthralgia induced by chikungunya virus infection is associated with interleukin-6 and granulocyte macrophage colony-stimulating factor. J. Infect. Dis. 2011, 203, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.F.; Chow, A.; Sun, Y.-J.; Kwek, D.J.; Lim, P.-L.; Dimatatac, F.; Ng, L.-C.; Ooi, E.-E.; Choo, K.-H.; Her, Z. Il-1β, il-6, and rantes as biomarkers of chikungunya severity. PLoS ONE 2009, 4, e4261. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.A.; Lee, Y.-J.; Seong, S.-C.; Choe, K.W.; Song, Y.W. Apoptotic chondrocyte death in human osteoarthritis. J. Rheumatol. 2000, 27, 455–462. [Google Scholar] [PubMed]
- Sandell, L.J.; Aigner, T. Articular cartilage and changes in arthritis: Cell biology of osteoarthritis. Arthritis Res. Ther. 2001, 3, 107. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fold Change in Gene Expression at Indicated Time Postinfection a | Fold Change in Gene Expression at Indicated Time Postinfection a | |||||
|---|---|---|---|---|---|---|
| Gene | 24 h | 48 h | Gene | 24 h | 48 h | |
| AIM2 | −1.11 | 15.85 | IRF3 | −1.41 | −1.14 | |
| APOBEC3G | 1.37 | 5.35 | IRF5 | −1.05 | −1.66 | |
| ATG5 | −1.44 | −1.58 | IRF7 | 9.04 | 24.08 | |
| AZI2 | −1.93 | −2.00 | ISG15 | 30.84 | 176.88 | |
| CARD9 | 1.38 | −1.00 | JUN | −1.15 | 1.11 | |
| CASP1 | 1.16 | 1.90 | MAP2K1 | 1.10 | 1.08 | |
| CASP10 | 2.31 | −1.07 | MAP2K3 | −1.61 | −1.79 | |
| CASP8 | −1.50 | −1.51 | MAP3K1 | −1.66 | −2.44 | |
| CCL3 | 1.87 | 4.42 | MAP3K7 | −1.80 | −1.78 | |
| CCL5 | 3.79 | 44.84 | MAPK1 | −1.64 | −1.54 | |
| CD40 | −1.54 | −1.09 | MAPK14 | −1.68 | −1.49 | |
| CD80 | −1.32 | −1.73 | MAPK3 | −1.16 | −1.94 | |
| CD86 | −1.29 | −1.00 | MAPK8 | −1.24 | −1.84 | |
| CHUK | −1.10 | 1.05 | MAVS | −1.99 | −1.22 | |
| CTSB | 1.06 | −3.50 | MEFV | 1.34 | −1.52 | |
| CTSL1 | −1.53 | 1.06 | MX1 | 36.17 | 43.51 | |
| CTSS | −1.3 | 2.91 | MYD88 | 1.43 | 2.71 | |
| CXCL10 | 25.28 | 304.44 | NFKB1 | 1.17 | 1.39 | |
| CXCL11 | 18,25 | 242.19 | NFKBIA | −1.11 | 2.17 | |
| CXCL9 | 1.25 | 1.77 | NLRP3 | 2.73 | 1.03 | |
| CYLD | −1.49 | −1.01 | NOD2 | 1.25 | −1.76 | |
| DAK | −1.12 | −1.53 | OAS2 | 13.06 | 28.18 | |
| DDX3X | −1.57 | −2.05 | PIN1 | 1.24 | 1.27 | |
| DDX58 (RIG–I) | 6.68 | 13.12 | PSTPIP1 | −1.65 | −2.44 | |
| DHX58 (LGP2) | 2.22 | 6.42 | PYCARD | −1.73 | −1.58 | |
| FADD | −1.21 | −1.68 | PYDC1 | 1.37 | 1.22 | |
| FOS | 15.42 | 9.00 | RELA | −1.01 | 1.41 | |
| HSP90AA1 | −1.38 | −1.03 | RIPK1 | 1.44 | 2.21 | |
| IFIH1 (MDA5) | 6.51 | 16.34 | SPP1 | −1.49 | 2.13 | |
| IFNA1 | 1.12 | 1.55 | STAT1 | 1.84 | 2.15 | |
| IFNA2 | 3.59 | 4.79 | SUGT1 | −1.84 | −1.61 | |
| IFNAR1 | −1.54 | −2.11 | TBK1 | −2.36 | −1.65 | |
| IFNB1 | 45.68 | 570.73 | TICAM1 | −1.30 | 1.52 | |
| IKBKB | −1.26 | −1.14 | TLR3 | 1.14 | 2.51 | |
| IL12A | −1.64 | −1.51 | TLR7 | 1.16 | 1.03 | |
| IL12B | −1.24 | 1.12 | TLR8 | 1.04 | 1.52 | |
| IL15 | 1.15 | 1.59 | TLR9 | −1.39 | −1.10 | |
| IL18 | −2.09 | −2.78 | TNF | ND | ND | |
| IL1B | 2.02 | 6.71 | TRADD | 1.27 | 1.30 | |
| IL6 | −2.24 | 2.24 | TRAF3 | −1.13 | −1.63 | |
| IL8 | −1.37 | 6.02 | TRAF6 | −2.66 | −1.77 | |
| IRAK1 | 1.84 | −1.16 | TRIM25 | 2.08 | 1.78 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bengue, M.; Ferraris, P.; Baronti, C.; Diagne, C.T.; Talignani, L.; Wichit, S.; Liegeois, F.; Bisbal, C.; Nougairède, A.; Missé, D. Mayaro Virus Infects Human Chondrocytes and Induces the Expression of Arthritis-Related Genes Associated with Joint Degradation. Viruses 2019, 11, 797. https://doi.org/10.3390/v11090797
Bengue M, Ferraris P, Baronti C, Diagne CT, Talignani L, Wichit S, Liegeois F, Bisbal C, Nougairède A, Missé D. Mayaro Virus Infects Human Chondrocytes and Induces the Expression of Arthritis-Related Genes Associated with Joint Degradation. Viruses. 2019; 11(9):797. https://doi.org/10.3390/v11090797
Chicago/Turabian StyleBengue, Michèle, Pauline Ferraris, Cécile Baronti, Cheikh Tidiane Diagne, Loïc Talignani, Sineewanlaya Wichit, Florian Liegeois, Catherine Bisbal, Antoine Nougairède, and Dorothée Missé. 2019. "Mayaro Virus Infects Human Chondrocytes and Induces the Expression of Arthritis-Related Genes Associated with Joint Degradation" Viruses 11, no. 9: 797. https://doi.org/10.3390/v11090797