Investigation about the Occurrence of Transmission Cycles of Arbovirus in the Tropical Forest, Amazon Region

 , ,
, ,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
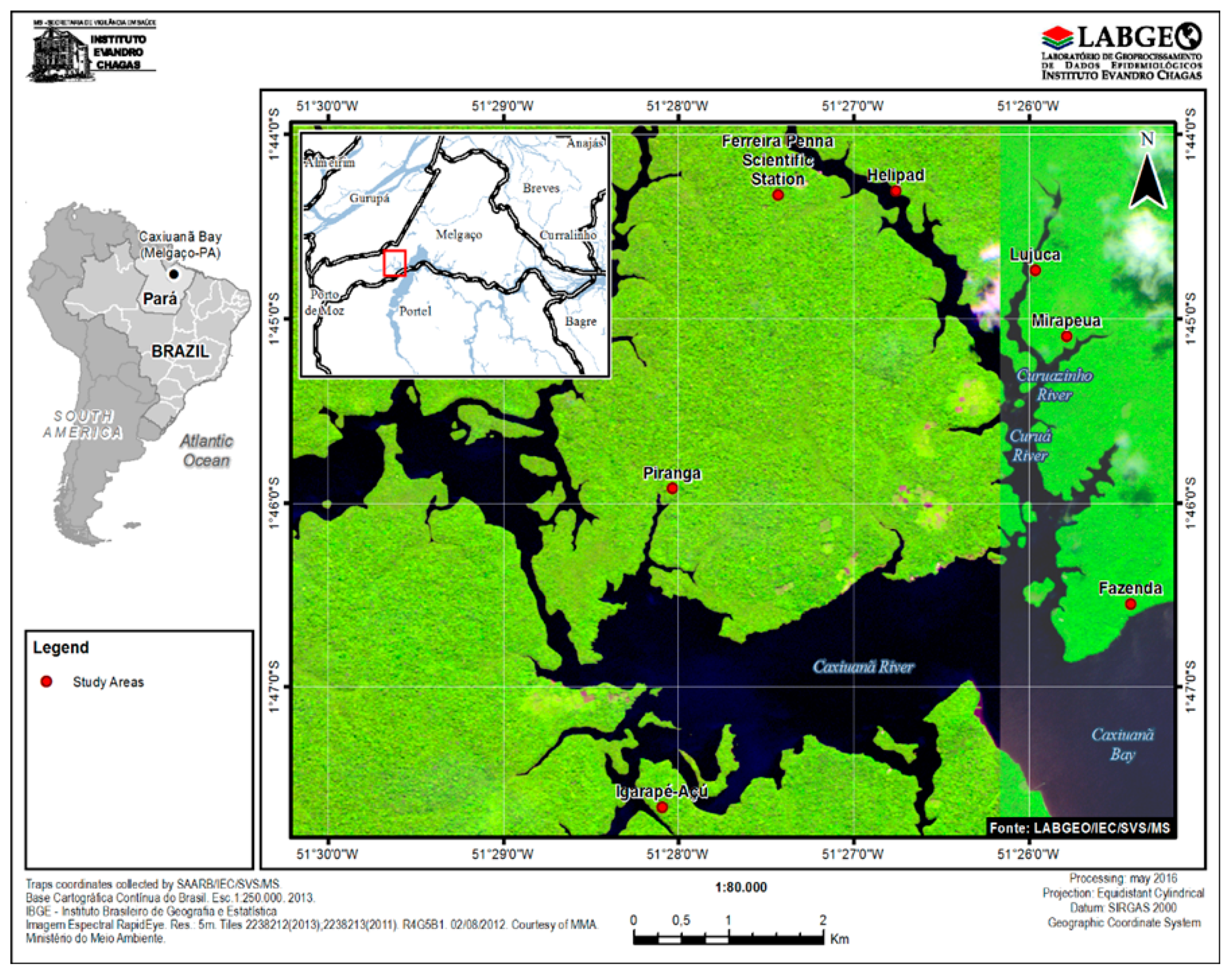
2.1. Study Areas
2.2. Collection Methods
2.3. Temperature and Humidity
2.4. Viral Isolation
2.5. Antibody Detection
2.6. Viral Genome Detection
2.7. Nucleotide Sequencing
2.8. Statistical Analysis
2.9. Legal Aspects
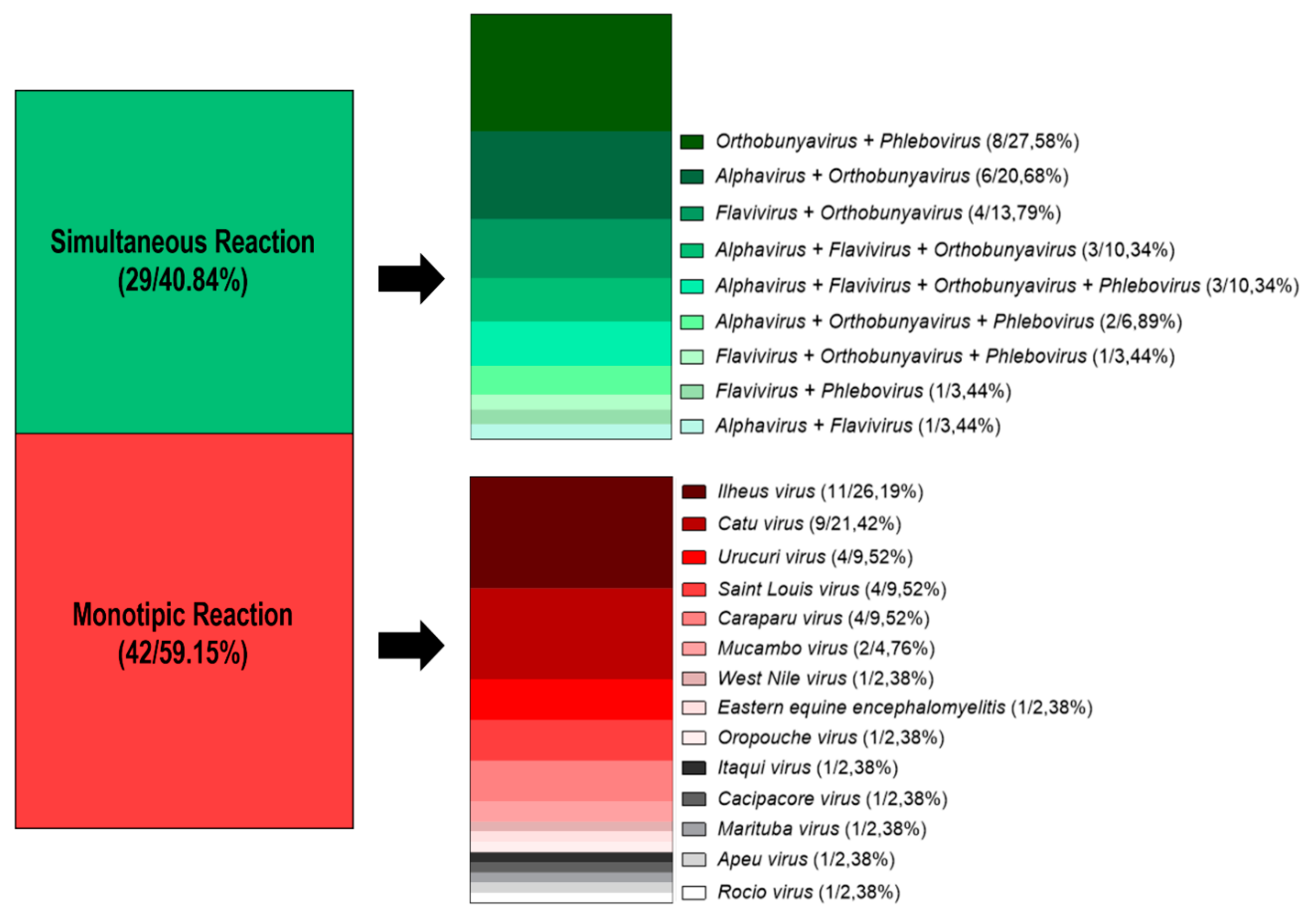
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Genus | Virus | Birds | Wild Vertebrates |
|---|---|---|---|
| Alphavirus | WEEV | X | X |
| EEEV | X | X | |
| MAYV | X | X | |
| MUCV | X | X | |
| AURAV | X | X | |
| PIXV | X | X | |
| Flavivirus | WNV | X | X |
| YFV | X | X | |
| ILHV | X | X | |
| SLEV | X | X | |
| ROCV | X | X | |
| CPCV | X | X | |
| BSQV | X | X | |
| NJLV | X | X | |
| Orthobunyavirus | MAGV | X | X |
| TACV | X | X | |
| CARV | X | X | |
| OROV | X | X | |
| CATV | X | X | |
| UTIV | - | X | |
| BLMV | X | - | |
| APEUV | X | X | |
| ITAV | X | X | |
| MARV | X | X | |
| MURV | X | X | |
| ORIV | X | X | |
| Phlebovirus | URUV | X | X |
| BUJV | X | X | |
| ICOV | - | X |
Appendix B
| Genus/Species | Viral Species | Sorological Titer | LNI |
|---|---|---|---|
| Proechimys sp. | ILHV | 1/320 | 0.8 |
| Proechimys sp. | ILHV | 1/640 | 1.0 |
| Proechimys sp. | ILHV | 1/640 | 1.0 |
| Chelonoidis denticulata | ILHV | 1/160 | 0.1 |
| Chelonoidis denticulata | ILHV | 1/160 | 0.2 |
| Proechimys sp. | MUCV | 1/640 | 1.9 |
| Oecomys sp. | MUCV | 1/320 | 1.9 |
| Oecomys sp. | MUCV | 1/40 | 2.1 |
| Proechimys sp. | MUCV | 1/40 | 1.3 |
| Proechimys sp. | MUCV | 1/160 | 2.2 |
| Oecomys sp. | MUCV | 1/80 | 2.0 |
| Oecomys sp. | MUCV | 1/80 | 0.4 |
| Metachirus nudicaudatus | MUCV | 1/1280 | 1.9 |
| Xiphorhynchus spixii | OROV | 1/80 | 1.8 |
References
- World Health Organization. Arbovirus and Human Diseases: Report of a WHO Scientific Group; World Health Organization: Genebra, Switzerland, 1967. [Google Scholar]
- Eldridge, B.F.; Scott, T.W.; Day, J.F.; Tabachnick, W.J. Arbovirus Diseases. In Medical Entomology; Springer: Dordrecht, The Netherlands, 2004; pp. 415–460. [Google Scholar]
- Lopes, N.; Nozawa, C.; Linhares, R.E.C. Características gerais e epidemiologia dos arbovírus emergentes no Brasil. Rev. Pan-Amazônica Saúde 2014, 5, 55–64. [Google Scholar] [CrossRef]
- Weaver, S.C.; Barrett, A.D.T. Transmission cycles, host range, evolution and emergence of arboviral disease. Nat. Rev. Microbiol. 2004, 2, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Dégallier, N.; Travassos, A.P.A.; Vasconcelos, P.F.C.; Travassos, J.F.S.; Figueiredo, L.T.M.; Rodrigues, S.G.; Travassos, E.S. La dengue et ses vecteurs au Brésil. Bull. Soc. Pathol. Exot. 1996, 89, 128–136. [Google Scholar] [PubMed]
- Mondet, B.; de Freitas, N.M.; Travassos da Rosa, A.P.D.A.; Vasconcelos, P.F.D.C.; Vieira, C.D.M.A. Biologia dos culicídeos vetores da febre amarela. In Caxiuanã: Populações tradicionais, meio físico e diversidade biológica; Lisboa, P.L.B., Ed.; Museu Paraense Emílio Goeldi: Belém, Brazil, 2002; p. 734. ISBN 8570980612. [Google Scholar]
- Cruz, A.C.R.; Prazeres, A.D.S.C.D.; Gama, E.C.; de Lima, M.F.; Azevedo, R.D.S.S.; Casseb, L.M.N.; Nunes Neto, J.P.; Martins, L.C.; Chiang, J.O.; Rodrigues, S.G.; et al. Vigilância sorológica para arbovírus em Juruti, Pará, Brasil. Cad. Saude Publica 2009, 25, 2517–2523. [Google Scholar] [CrossRef] [PubMed]
- Araújo, F.A.A. Inquéritos sorológicos em equídeos e aves silvestres para detecção de anticorpos anti-arbovírus de importância em saúde pública no Brasil. Master’s Thesis, Universidade Federal de Goiás, Goiânia, Braizl, 6 December 2011. [Google Scholar]
- Igarashi, A. Isolation of a Singh’s Aedes albopictus Cell Clone Sensitive to Dengue and Chikungunya Viruses. J. Gen. Virol. 1978, 40, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Travassos da Rosa, J.F.S.; Travassos da Rosa, A.P.A.; Vasconcelos, P.F.C.; Pinheiro, F.P.; Rodrigues, S.G.; Travassos, E.S.; Dias, L.B.; Cruz, A.C. Arboviruses isolated in the Evandro Chagas Institute, including some described for the first time in the Brazilian Arnazon region, their known hosts, and their pathology for man. In An Overview of Arbovirology in Brazil and Neighbouring Countries; Travassos da Rosa, A.P.A., Vasconcelos, P.F.C., Travassos da Rosa, J.F.S., Eds.; Instituto Evandro Chagas: Belém, Brzail, 1998; pp. 186–192. ISBN 85-86784-01-X. [Google Scholar]
- Gubler, D.J.; Kuno, G.; Sather, G.E.; Velez, M.; Oliver, A. Mosquito Cell Cultures and Specific Monoclonal Antibodies in Surveillance for Dengue Viruses. Am. J. Trop. Med. Hyg. 1984, 33, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Clarke, D.H.; Casals, J. Techniques for hemagglutination and hemagglutination-inhibition with arthropod-borne viruses. Am. J. Trop. Med. Hyg. 1958, 7, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Shope, R.E. The use of a micro hemagglutination- inhibition test to follow antibody response after arthropod-borne virus infection in a community of forest animals. An. Microbiol. (Rio. J.) 1963, 11, 167–169. [Google Scholar]
- Casals, J. Antigenic characterization of tick-borne arboviruses not belonging to group B. Jpn J. Med. Sci. Biol. 1967, 20, 119–129. [Google Scholar]
- Reed, L.J.; Muench, H. A Simple Method Of Estimating Fifty Per Cent Endpoints12. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Pfeffer, M.; Kinney, R.M.; Kaaden, O.-R. The Alphavirus 3′-Nontranslated Region: Size Heterogeneity and Arrangement of Repeated Sequence Elements. Virology 1998, 240, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Kuno, G.; Chang, G.J.; Tsuchiya, K.R.; Karabatsos, N.; Cropp, C.B. Phylogeny of the genus Flavivirus. J. Virol. 1998, 72, 73–83. [Google Scholar] [PubMed]
- Peng, Y.; Leung, H.C.M.; Yiu, S.M.; Chin, F.Y.L. IDBA-UD: A de novo assembler for single-cell and metagenomic sequencing data with highly uneven depth. Bioinformatics 2012, 28, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Degallier, N.; Travassos da Rosa, A.P.A.; da Silva, J.M.C.; Rodrigues, S.G.; Vasconcelos, P.F.D.C.; Travassos da Rosa, J.F.S.; da Silva, G.P.; Pereira da Silva, R. As aves como hospedeiras de arbovírus na Amazônia brasileira. Bol. Mus. Para. Emilio Goeldi Sér. Zool. 1992, 8, 69–111. [Google Scholar]
- Pereira, L.E.; Suzuki, A.; Coimbra, T.L.; de Souza, R.P.; Chamelet, E.L. Arbovírus Ilheus em aves silvestres (Sporophila caerulescens e Molothrus bonariensis). Rev. Saude Publica 2001, 35, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Monath, T.; Kemp, G.E.; Cropp, C.B.; Bowen, S. Experimental infection of House Sparrows (Passer domesticus) with Rocio virus. Am. Soc. Trop. Med. Hyg. 1978, 27, 1251–1254. [Google Scholar] [CrossRef]
- Rappole, J.H.; Derrickson, S.R.; Hubálek, Z. Migratory birds and spread of West Nile virus in the Western Hemisphere. Emerg. Infect. Dis. 2000, 6, 319–328. [Google Scholar] [CrossRef]
- Freitas, M.N.O. Investigação sorológica para arbovírus em vertebrados silvestres oriundos da região de Carajás, no período de 2012 a 2015. In preparation.
- dos S. Moraes, J.R. Avaliação da diversidade de aves e roedores e a circulação de arbovírus em áreas da província mineral de Carajás-Estado do Pará de 2012 a 2015. Master’s Thesis, Universidade do Estado do Pará, Belém do Pará, Braizl, 20 April 2016. [Google Scholar]
- Causey, O.R.; Chusey, C.E.; Maroja, O.H.; Macedo, D.G. The isolation of arthropod-borne viruses, including members of two hitherto undescribed serological groups, in the Amazon region of Brazil. Am. Soc. Trop. Med. Hyg. 1961, 10, 227–249. [Google Scholar] [CrossRef]
- Jonkers, A.H.; Spence, L.; Downs, W.G.; Aitken, T.H.G.; Worth, C. Rodent-Associated Viruses (VEE and Agents of Groups C and Guamá): Isolations and Further Studies*. Am. Soc. Trop. Med. Hyg. 1968, 17, 285–298. [Google Scholar] [CrossRef]
- Aitken, T.H.G.; Spence, L.; Jonkers, A.H.; Downs, W.G. A 10-Year Survey Of Trinidadian Arthropods For Natural Virus Infections (1953–1963). J. Med. Entomol. 1969, 6, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Serié, C. Rapport sur le fonctionnement technique de l’Institut Pasteur de la Guyane fransaise pendant l’annde 1969. Arch. Inst. Pasteur Guyane Inini. Publ. 1970, 21, 518–527. [Google Scholar]
- Digoutte, J.P. Rapport annuel dractivites du groupe de Rècherche U79, I.N.S.E.R.M. Inst. Pasteur la Guyane Fr. 1976. [Google Scholar]
- Reeves, W.C. Ecology of Mosquitoes in Relation to Arboviruses. Annu. Rev. Entomol. 1965, 10, 25–46. [Google Scholar] [CrossRef]
- Hoshino, K.; Isawa, H.; Tsuda, Y.; Yano, K.; Sasaki, T.; Yuda, M.; Takasaki, T.; Kobayashi, M.; Sawabe, K. Genetic characterization of a new insect flavivirus isolated from Culex pipiens mosquito in Japan. Virology 2007, 359, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Evangelista, J.; Cruz, C.; Guevara, C.; Astete, H.; Carey, C.; Kochel, T.J.; Morrison, A.C.; Williams, M.; Halsey, E.S.; Forshey, B.M. Characterization of a novel flavivirus isolated from Culex (Melanoconion) ocossa mosquitoes from Iquitos, Peru. J. Gen. Virol. 2013, 94, 1266–1272. [Google Scholar] [CrossRef] [PubMed]
- Go, Y.Y.; Balasuriya, U.B.R.; Lee, C. Zoonotic encephalitides caused by arboviruses: Transmission and epidemiology of alphaviruses and flaviviruses. Clin. Exp. Vaccine Res. 2014, 3, 58. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, F.P.; Travassos da Rosa, A.P.A.; Travassos da Rosa, J.F.S.; Bensabath, G. An outbreak of Oropouche virus diease in the vicinity of santarem, para, barzil. Tropenmed. Parasitol. 1976, 27, 213–223. [Google Scholar] [PubMed]
- Araújo, F.A.A.; Vianna, R.S.T.; Wada, M.Y.; Silva, E.V.; Cavalcante, G.C.; Magalhães, V.S.; Andrade Filho, G.V.; Rodrigues, S.G.; Martins, L.C.; Fedrizzi, C.E.; et al. Primeiro Inquérito Sorológico em Aves Migratórias e Nativas do Parque Nacional da Lagoa do Peixe para detecção do Vírus da febre do Nilo Ocidental. Bol. Eletrônico 2004, 1, 1–8. [Google Scholar]
- Araujo, F.A.A.; Lima, P.C.; Andrade, M.A.; Jayme, V.D.S.; Ramos, D.G.; Da Silveira, S.L. Soroprevalência de anticorpos “anti-arbovírus” de importância em saúde pública em aves selvagens, brasil—2007 e 2008. Cienc. Anim. Bras. 2012, 13, 115–123. [Google Scholar]
- Roberts, D.R.; Pinheiro, F.D.P.; Hoch, A.L.; Leduc, J.W.; Peterson, N.E.; Santos, M.A.V.; Pinheiro, F.D.P.; Western, K.A. Vectors and Natural Reservoirs of Oropouche Virus in the Amazon Region; PN (1977): Washington, DC, USA, 1977. [Google Scholar]
- De Souza-Lopes, O.; Sacchetta, L.A. Isolation of Mucambo virus, a member of the Venezuelan equine encephalitis virus complex in the State of São Paulo, Brazil. Rev. Inst. Med. Trop. Sao Paulo 1978, 20, 82–86. [Google Scholar] [PubMed]
- Pinheiro, F.P.; Pinheiro, M.; Bensabath, G.; Causey, O.R.; Shope, R.E. Oropouche virus epidemic in Bélem. Rev. Serv. Esp. Saude Pub. 1962, 12, 15–23. [Google Scholar]
- Vasconcelos, P.F.; Da Rosa, J.F.; Da Rosa, A.P.; Dégallier, N.; Pinheiro, F.d.P.; Sá Filho, G.C. Epidemiology of encephalitis caused by arbovirus in the Brazilian Amazonia. Rev. Inst. Med. Trop. Sao Paulo 1991, 33, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.A. Estudo sobre a circulação de arbovírus na população humana nas áreas de influência na Floresta Nacional de Caxiuanã, Melgaço, Estado do Pará. Master’s Thesis, Instituto Evandro Chagas, Ananindeua, Brazil, 5 May 2017. [Google Scholar]
- Forattini, O.P. Some data on the domesticity of Aedes scapularis (Rondani) in São Paulo, Brazil. Mosq. News 1961, 21, 295–296. [Google Scholar]
- Forattini, O.P.; Gomes, A.D.C.; Galati, E.A.B.; Rabello, E.X.; Iversson, L.B. Ecológicos Sobre Mosquitos Culicidae no sistema da Serra do Mar, Brasil. Rev. Saude Publica 1978, 12, 476–496. [Google Scholar] [CrossRef] [PubMed][Green Version]


| Pool of Species | Season | Collection Area | Method of Capture | Virus | Homology |
|---|---|---|---|---|---|
| Ur. (Ura.) geometrica | Dry | ECFPn | CDC (Solo) | MUCV | 83% |
| Cx. (Mel.) portesi | Dry | ECFPn | CDC (Solo) | ROCV | 92% |
| Cx. (Mel.) portesi | Dry | ECFPn | CDC (Solo) | ILHV | 94% |
| Cx. (Mel.) portesi | Dry | ECFPn | CDC (Solo) | BSQV | 94% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Araújo, P.A.; Freitas, M.O.; Chiang, J.O.; Silva, F.A.; Chagas, L.L.; Casseb, S.M.; Silva, S.P.; Nunes-Neto, J.P.; Rosa-Júnior, J.W.; Nascimento, B.S.; et al. Investigation about the Occurrence of Transmission Cycles of Arbovirus in the Tropical Forest, Amazon Region. Viruses 2019, 11, 774. https://doi.org/10.3390/v11090774
Araújo PA, Freitas MO, Chiang JO, Silva FA, Chagas LL, Casseb SM, Silva SP, Nunes-Neto JP, Rosa-Júnior JW, Nascimento BS, et al. Investigation about the Occurrence of Transmission Cycles of Arbovirus in the Tropical Forest, Amazon Region. Viruses. 2019; 11(9):774. https://doi.org/10.3390/v11090774
Chicago/Turabian StyleAraújo, Pedro A., Maria O. Freitas, Jannifer Oliveira Chiang, Franko Arruda Silva, Liliane Leal Chagas, Samir Mansour Casseb, Sandro Patroca Silva, Joaquim Pinto Nunes-Neto, José Wilson Rosa-Júnior, Bruna Sena Nascimento, and et al. 2019. "Investigation about the Occurrence of Transmission Cycles of Arbovirus in the Tropical Forest, Amazon Region" Viruses 11, no. 9: 774. https://doi.org/10.3390/v11090774
APA StyleAraújo, P. A., Freitas, M. O., Chiang, J. O., Silva, F. A., Chagas, L. L., Casseb, S. M., Silva, S. P., Nunes-Neto, J. P., Rosa-Júnior, J. W., Nascimento, B. S., Hernández, L. A., Paz, T. B., Barros, L. L., Silva, E. P., Azevedo, R. S., Martins, L. C., Ferreira, M. S., & Vasconcelos, P. F. (2019). Investigation about the Occurrence of Transmission Cycles of Arbovirus in the Tropical Forest, Amazon Region. Viruses, 11(9), 774. https://doi.org/10.3390/v11090774