Field Evaluation of Deltamethrin and Ivermectin Applications to Cattle on Culicoides Host-Alighting, Blood-Feeding, and Emergence

 ,
,
Abstract
:
1. Introduction
2. Material and Methods
2.1. Trial 1—Five Week Study on the Effects of Deltamethrin and Ivermectin on Culicoides’ Landing and Emergence from Dung
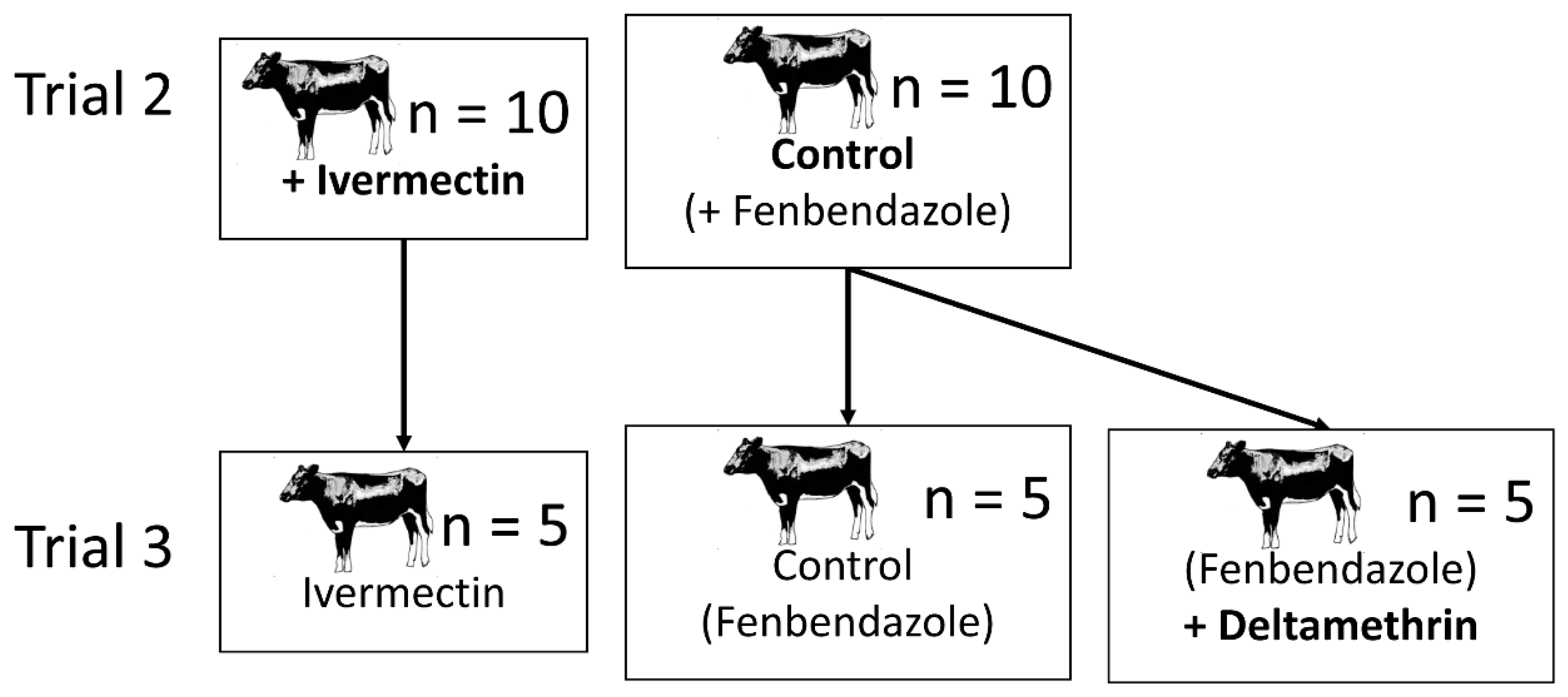
2.2. Trial 2—Culicoides’ Landing and Blood-Feeding on Friesian Heifers Treated with Ivermectin Pour-On
2.3. Trial 3—Culicoides’ Landing, Blood-Feeding and Mortality on Friesian Heifers Treated with Deltamethrin and Ivermectin Pour-Ons
2.4. Analyses
3. Results
3.1. Trial 1—Five Week Study on the Effects of Ivermectin and Deltamethrin on Culicoides’ Landing and Emergence from Dung
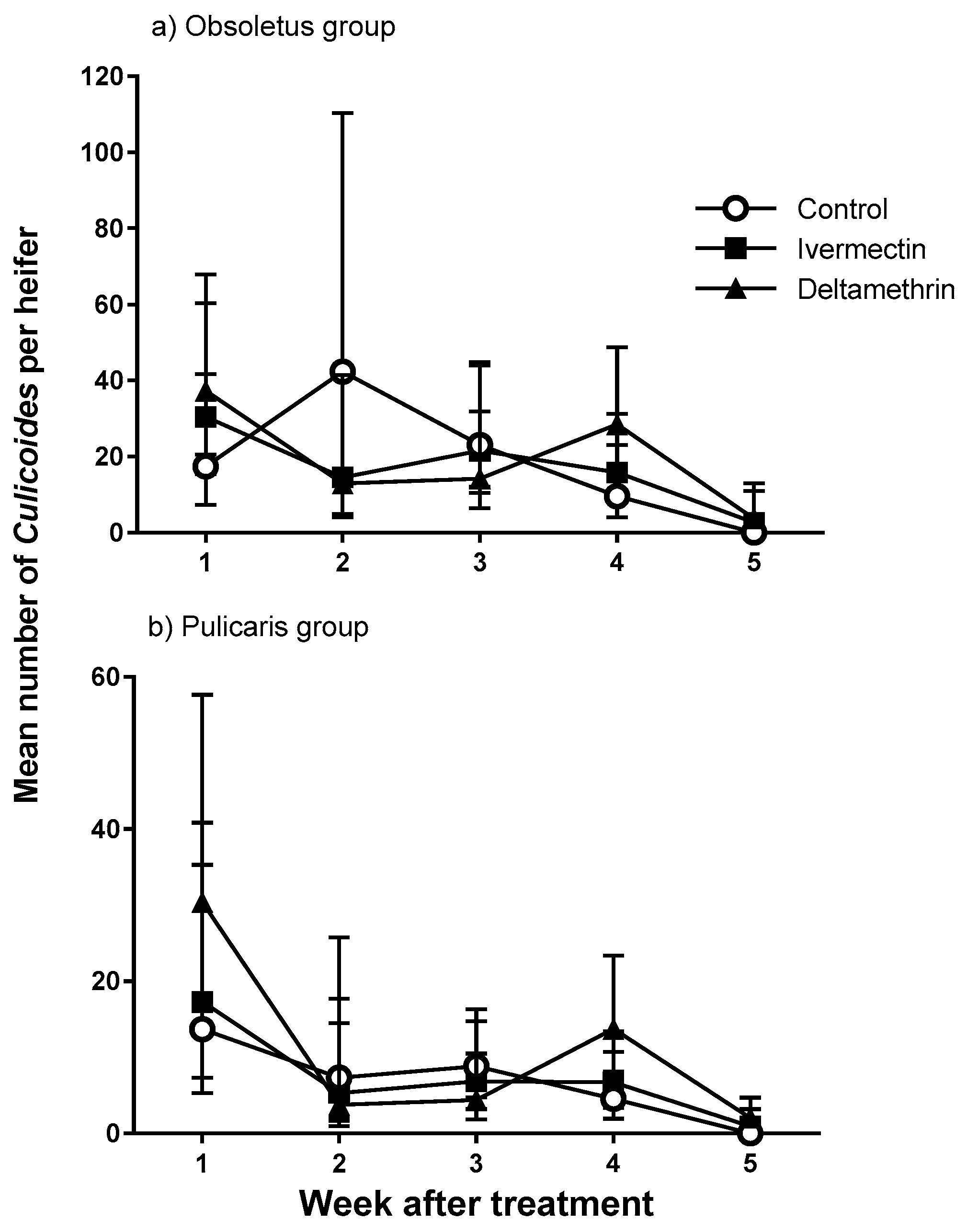
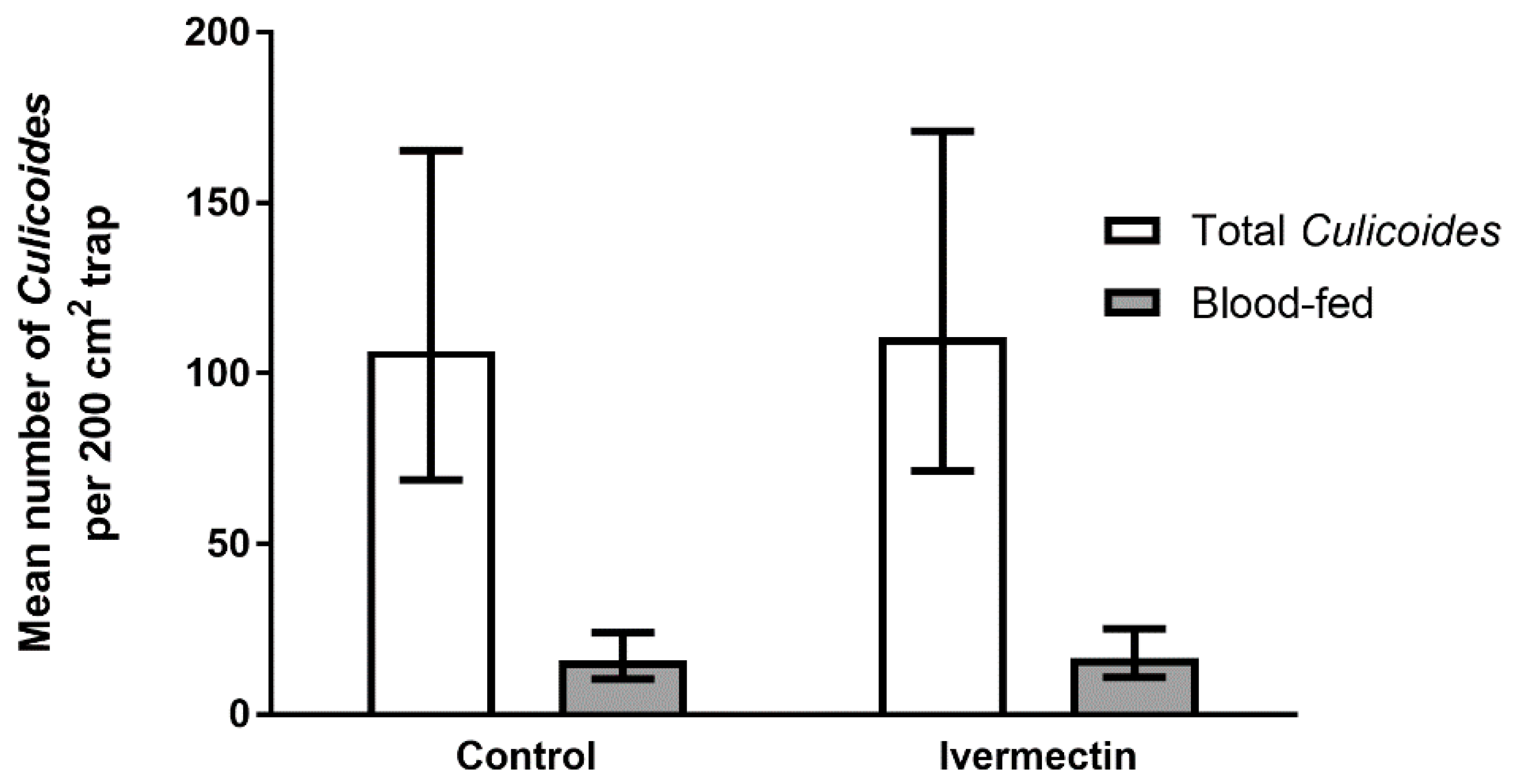
3.2. Trial 2—Culicoides’ Landing and Blood-Feeding on Friesian Heifers Treated with Ivermectin Pour-On
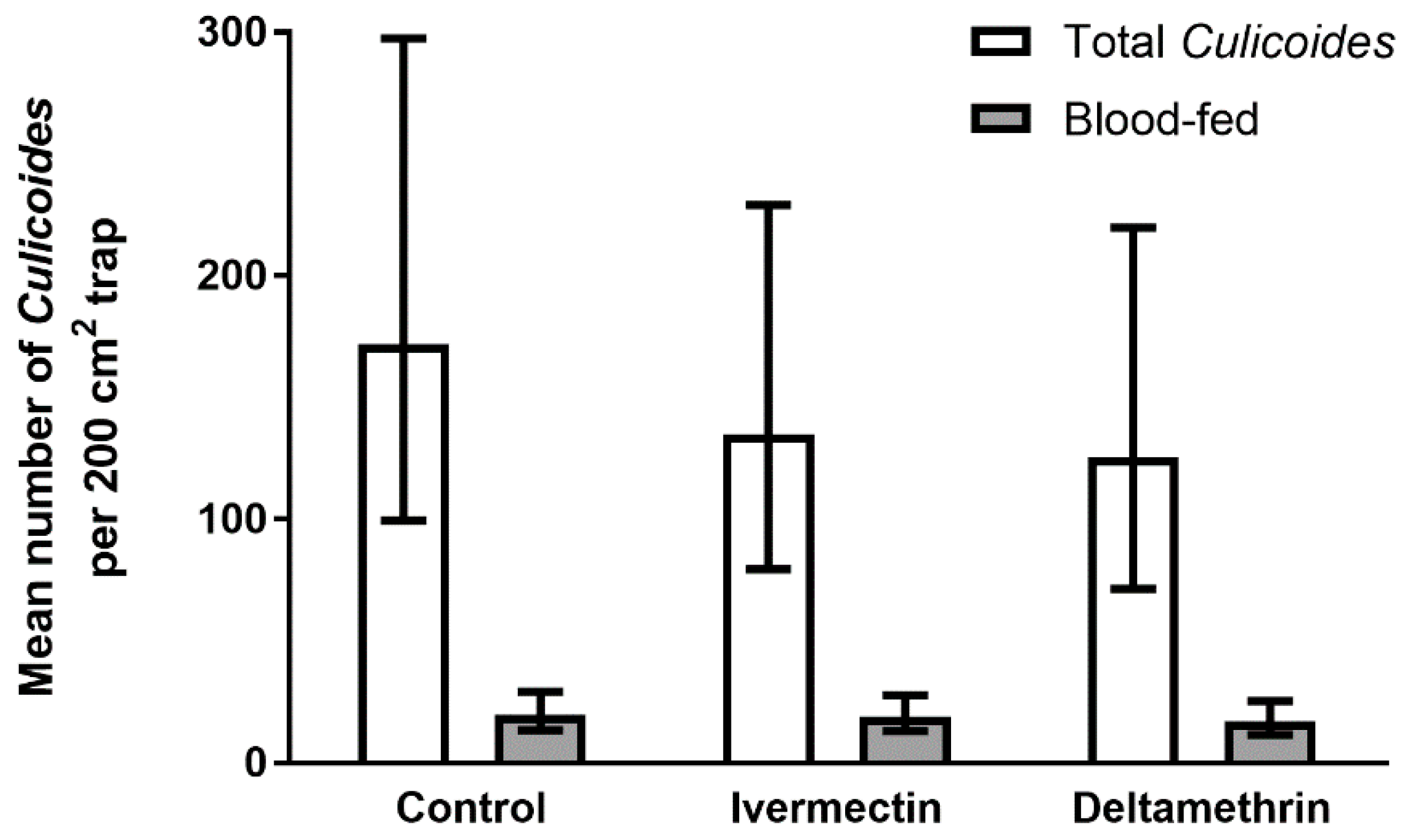
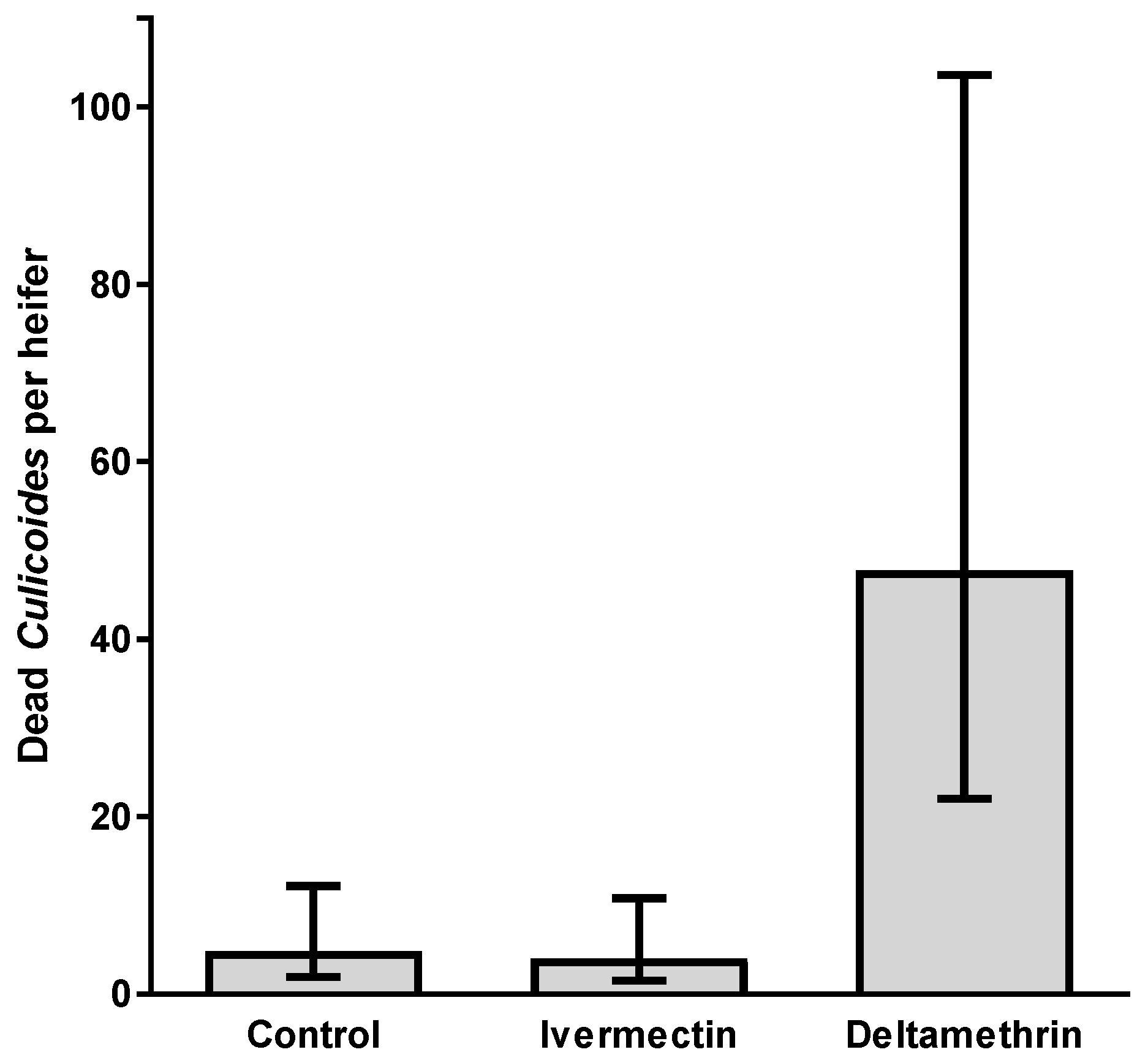
3.3. Trial 3—Culicoides’ Landing, Blood-Feeding and Mortality on Friesian Heifers Treated with Deltamethrin and Ivermectin Pour-Ons
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Harrup, L.E.; Miranda, M.A.; Carpenter, S. Advances in control techniques for Culicoides and future prospects. Vet. Ital. 2016, 52, 247–264. [Google Scholar] [PubMed]
- Menzies, F.D.; McCullough, S.J.; McKeown, I.M.; Forster, J.L.; Jess, S.; Batten, C.; Murchie, A.K.; Gloster, J.; Fallows, J.G.; Pelgrim, W.; et al. Evidence for transplacental and contact transmission of bluetongue virus in cattle. Vet. Rec. 2008, 163, 203–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DAERA. Bluetongue Detected in Imported Animal. Available online: https://www.daera-ni.gov.uk/news/bluetongue-detected-imported-animal (accessed on 6 December 2018).
- European Commission. Commission Regulation number 1266/2007 on implementation rules for Council Directive 2005/75/EC as regards the control, monitoring, surveillance and restrictions on movements of certain animals of susceptible species in relation to Bluetongue. OJEU 2007, 283, 37–52. [Google Scholar]
- Maclachlan, N.J.; Mayo, C.E. Potential strategies for control of bluetongue, a globally emerging, Culicoides-transmitted viral disease of ruminant livestock and wildlife. Antiviral Res. 2013, 99, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, S.; Mellor, P.S.; Torr, S.J. Control techniques for Culicoides biting midges and their application in the U.K. and northwestern Palaearctic. Med. Vet. Entomol. 2008, 22, 175–187. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific Opinion of the Scientific Panel on Animal Health and Welfare on request from the European Commission on bluetongue vectors and vaccines. EFSA J. 2007, 479, 1–29. [Google Scholar]
- EFSA. Bluetongue vectors and insecticides. Scientific Opinion of the Panel on Animal Health and Welfare on a request from the European Commission (DG SANCO) on Bluetongue. EFSA J. 2008, 735, 1–70. [Google Scholar]
- Mullens, B.A.; McDermott, E.G.; Gerry, A.C. Progress and knowledge gaps in Culicoides ecology and control. Vet. Ital. 2015, 51, 313–323. [Google Scholar]
- Doherty, W.M.; Johnson, S.J.; Reid, A.E. Suppression of Culicoides brevitarsis (Kieffer) (Diptera: Ceratopogonidae) on cattle in Queensland with deltamethrin and cypermethrin. Gen. Appl. Ent. 2001, 30, 45–47. [Google Scholar]
- Melville, L.; Hunt, N.; Bellis, G.; Pinch, D. Evaluation of chemical treatments to prevent Culicoides spp. (Diptera: Ceratopogonidae) feeding on cattle in the Northern Territory. Gen. Appl. Ent. 2001, 30, 41–44. [Google Scholar]
- Melville, L.V.; Hunt, N.T.; Bellis, G.; Pinch, D. An assessment of insecticides to minimize the transmission of arbovirus in cattle. Arbovirus Res. Aust. 2004, 8, 249–255. [Google Scholar]
- Doherty, W.M.; Bishop, A.L.; Melville, L.F.; Johnson, S.J.; Bellis, G.A.; Hunt, N.T. Protection of cattle from Culicoides spp. in Australia by shelter and chemical treatments. Vet. Ital. 2004, 40, 321. [Google Scholar]
- Mullens, B.A.; Velten, R.K.; Gerry, A.C.; Braverman, Y.; Endris, R.G. Feeding and survival of Culicoides sonorensis on cattle treated with permethrin or pirimiphos-methyl. Med. Vet. Entomol. 2000, 14, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Bauer, B.; Jandowsky, A.; Schein, E.; Mehlitz, D.; Clausen, P.-H. An appraisal of current and new techniques intended to protect bulls against Culicoides and other haematophagous nematocera: the case of Schmergow, Brandenburg, Germany. Parasitol. Res. 2009, 105, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Mullens, B.A.; Gerry, A.C.; Monteys, V.S.I.; Pinna, M.; Gonzalez, A. Field studies on Culicoides (Diptera: Ceratopogonidae) activity and response to deltamethrin applications to sheep in northeastern Spain. J. Med. Entomol. 2010, 47, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Weiher, W.; Bauer, B.; Mehlitz, D.; Nijhof, A.M.; Clausen, P.-H. Field trials assessing deltamethrin (Butox®) treatments of sheep against Culicoides species. Parasitol. Res. 2014, 113, 2641–2645. [Google Scholar] [CrossRef] [PubMed]
- De Keyser, R.; Cassidy, C.; Laban, S.; Gopal, P.; Pickett, J.A.; Reddy, Y.K.; Prasad, M.; Prasad, G.; Chirukandoth, S.; Senthilven, K.; et al. Insecticidal effects of deltamethrin in laboratory and field populations of Culicoides species: how effective are host-contact reduction methods in India? Parasit. Vectors. 2017, 10, 54. [Google Scholar] [CrossRef] [PubMed]
- Robin, M.; Archer, D.; McGowan, C.; Garros, C.; Gardès, L.; Baylis, M. Repellent effect of topical deltamethrin on blood feeding by Culicoides on horses. Vet. Rec. 2015, 176, 574. [Google Scholar] [CrossRef]
- Standfast, H.A.; Muller, M.J.; Wilson, D.D. Mortality of Culicoides brevitarsis (Diptera: Ceratopogonidae) fed on cattle treated with Ivermectin. J. Econ. Entomol. 1984, 77, 419–421. [Google Scholar] [CrossRef]
- Holbrook, F.R. Survival, fecundity, and egg fertility of Culicoides variipennis (Diptera: Ceratopogonidae) fed on calves inoculated with ivermectin. J. Am. Mosq. Control. Assoc. 1994, 10, 7. [Google Scholar]
- Holbrook, F.R.; Mullens, B.A. Effects of ivermectin on survival, fecundity, and egg fertility in Culicoides variipennis (Diptera: Ceratopogonidae). J. Am. Mosq. Control. Assoc. 1994, 10, 70–73. [Google Scholar] [PubMed]
- Reeves, W.K.; Nol, P.; Miller, M.M.; Jones, G.Z. Effects of ivermectin on the susceptibility of Culicoides sonorensis (Diptera: Ceratopogonidae) to bluetongue and epizootic hemorrhagic disease viruses. J. Vector Ecol. 2009, 34, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Webster, W.R.; Gard, G.P.; St. George, T.D.; Kirkland, P.D. The Australian Bluetongue Control Strategy. In Proceedings of the Second International Symposium: Bluetongue, African Horse Sickness, and Related Orbiviruses, Paris, France, 17–21 June 1991; CRC Press Inc.: Boca Raton, FL, USA, 1992; pp. 843–850. [Google Scholar]
- Thompson, G.M.; Jess, S.; Gordon, A.W.; Murchie, A.K. Sticky-trapping biting midges (Culicoides spp.) alighting on cattle and sheep: Effects of trap colour and evidence for host preference. Parasitol. Res. 2014, 113, 3085–3094. [Google Scholar] [CrossRef] [PubMed]
- Ayllón, T.; Nijhof, A.M.; Weiher, W.; Bauer, B.; Allène, X.; Clausen, P.-H. Feeding behaviour of Culicoides spp. (Diptera: Ceratopogonidae) on cattle and sheep in northeast Germany. Parasit. Vectors 2014, 7, 34. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, B.O. Some observations on biting midges (Diptera: Ceratopogonidae) attacking grazing cattle in Denmark. Entomol. Scand. 1971, 2, 94–98. [Google Scholar] [CrossRef]
- Townley, P.; Baker, K.; Quinn, P. Preferential landing and engorging sites of Culicoides species landing on a horse in Ireland. Equine Vet. J. 1984, 16, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Venail, R.; Mathieu, B.; Setier-Rio, M.-L.; Borba, C.; Alexandre, M.; Viudes, G.; Garros, C.; Allene, X.; Carpenter, S.; Baldet, T. Laboratory and field-based tests of deltamethrin insecticides against adult Culicoides biting midges. J. Med. Entomol. 2011, 48, 351–357. [Google Scholar] [CrossRef]
- Gayrard, V.; Alvinerie, M.; Toutain, P. Comparison of pharmacokinetic profiles of doramectin and ivermectin pour-on formulations in cattle. Vet. Parasitol. 1999, 81, 47–55. [Google Scholar] [CrossRef]
- Gokbulut, C.; Cirak, V.Y.; Senlik, B.; Aksit, D.; Durmaz, M.; McKellar, Q.A. Comparative plasma disposition, bioavailability and efficacy of ivermectin following oral and pour-on administrations in horses. Vet. Parasitol. 2010, 170, 120–126. [Google Scholar] [CrossRef]
- Nielsen, S.A.; Kristensen, M. Morphological and molecular identification of species of the Obsoletus group (Diptera: Ceratopogonidae) in Scandinavia. Parasitol. Res. 2011, 109, 1133–1141. [Google Scholar] [CrossRef]
- Bartsch, S.; Bauer, B.; Wiemann, A.; Clausen, P.-H.; Steuber, S. Feeding patterns of biting midges of the Culicoides obsoletus and Culicoides pulicaris groups on selected farms in Brandenburg, Germany. Parasitol. Res. 2009, 105, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Boorman, J. A short key to British Culicoides of veterinary importance. Available online: http: www.culicoides.net/taxonomy/identification-keys/UK01–1 (accessed on 3 July 2012).
- Dyce, A.L. The recognition of nulliparous and parous Culicoides (Diptera: Ceratopogonidae) without dissection. J. Aust. Entomol. Soc. 1969, 8, 11–15. [Google Scholar] [CrossRef]
- Thompson, G.M.; Brunt, A.; Loye, S.; Jess, S.; Murchie, A.K. A low-cost bucket trap for newly emerged insects. Ir. Nat. J. 2013, 32, 117–120. [Google Scholar]
- Achee, N.L.; Sardelis, M.S.; Dusfour, I.; Chauhan, K.R.; Grieco, J.P. Characterisation of spatial repellent, contact irritant, and toxicant chemical actions of standard vector control compounds. J. Am. Mosq. Control. Assoc. 2009, 25, 156–157. [Google Scholar] [CrossRef] [PubMed]
- Mehlhorn, H.; Al-Rasheid, K.A.S.; Abdel-Ghaffar, F.; Klimpel, S.; Pohle, H. Life cycle and attacks of ectoparasites on ruminants during the year in Central Europe: recommendations for treatment with insecticides (e.g., Butox®). Parasitol. Res. 2010, 107, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Sollai, G.; Solari, P.; Masala, C.; Crnjar, R.; Liscia, A. Effects of avermectins on olfactory responses of Culicoides imicola (Diptera: Ceratopogonidae). J. Med. Entomol. 2007, 44, 656–659. [Google Scholar] [CrossRef]
- Del Río, R.; Venail, R.; Calvete, C.; Barceló, C.; Baldet, T.; Lucientes, J.; Miranda, M.A. Sensitivity of Culicoides obsoletus (Meigen) (Diptera: Ceratopogonidae) to deltamethrin determined by an adapted WHO standard susceptibility test. Parasitology 2014, 141, 542–546. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, S.; Mellor, P.; Torr, S. Bluetongue and midge control. Vet. Rec. 2007, 161, 633. [Google Scholar] [CrossRef]
- Schmahl, G.; Klimpel, S.; Walldorf, V.; Al-Quraishy, S.; Schumacher, B.; Jatzlau, A.; Mehlhorn, H. Pilot study on deltamethrin treatment (Butox® 7.5, Versatrine®) of cattle and sheep against midges (Culicoides species, Ceratopogonidae). Parasitol. Res. 2009, 104, 809–813. [Google Scholar] [CrossRef]
- Mehlhorn, H.; Schmahl, G.; D’Haese, J.; Schumacher, B. Butox 7.5 pour on: A deltamethrin treatment of sheep and cattle: pilot study of killing effects on Culicoides species (Ceratopogonidae). Parasitol. Res. 2008, 102, 515–518. [Google Scholar] [CrossRef]
- Murchie, A.K.; Clawson, S.; Rea, I.; Forsythe, I.W.N.; Gordon, A.W.; Jess, S. DEET (N,N-diethyl-meta-toluamide)/PMD (para-menthane-3,8-diol) repellent-treated mesh increases Culicoides catches in light traps. Parasitol. Res. 2016, 115, 3543–3549. [Google Scholar] [CrossRef] [PubMed]
- Wall, R.; Strong, L. Environmental consequences of treating cattle with the antiparasitic drug ivermectin. Nature 1987, 327, 418–421. [Google Scholar] [CrossRef] [PubMed]
- Strong, L. Avermectins: A review of their impact on insects of cattle dung. Bull. Entomol. Res. 1992, 82, 265–274. [Google Scholar] [CrossRef]
- Wall, R.; Beynon, S. Area-wide impact of macrocyclic lactone parasiticides in cattle dung. Med. Vet. Entomol. 2012, 26, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Floate, K.D.; Spooner, R.W.; Colwell, D.D. Larvicidal activity of endectocides against pest flies in the dung of treated cattle. Med. Vet. Entomol. 2001, 15, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Strong, L.; Brown, T.A. Avermectins in insect control and biology: A review. Bull. Entomol. Res. 1987, 77, 357–389. [Google Scholar] [CrossRef]
- Mann, C.M.; Barnes, S.; Offer, B.; Wall, R. Lethal and sub-lethal effects of faecal deltamethrin residues on dung-feeding insects. Med. Vet. Entomol. 2015, 29, 189–195. [Google Scholar] [CrossRef]
- Gilbert, G.; MacGillivray, F.S.; Robertson, H.L.; Jonsson, N.N. Adverse effects of routine bovine health treatments containing triclabendazole and synthetic pyrethroids on the abundance of dipteran larvae in bovine faeces. Sci. Rep. 2019, 9, 4315. [Google Scholar] [CrossRef]
- Jess, S.; Thompson, G.M.; Clawson, S.; Forsythe, I.W.N.; Rea, I.; Gordon, A.W.; Murchie, A.K. Surveillance of biting midges (Culicoides spp.) in Northern Ireland: influence of seasonality, surrounding habitat and livestock housing. Med. Vet. Entomol. 2018, 32, 48–60. [Google Scholar] [CrossRef]
- Viennet, E.; Garros, C.; Gardès, L.; Rakotoarivony, I.; Allène, X.; Lancelot, R.; Crochet, D.; Moulia, C.; Baldet, T.; Balenghien, T. Host preferences of Palaearctic Culicoides biting midges: implications for transmission of orbiviruses. Med. Vet. Entomol. 2013, 27, 255–266. [Google Scholar] [CrossRef]
- Elbers, A.; Koenraadt, C.; Meiswinkel, R. Mosquitoes and Culicoides biting midges: vector range and the influence of climate change. Rev. Sci. Tech. 2015, 34, 123–137. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial Number | Date | Days after Ivermectin Treatment | Days after Deltamethrin Treatment | Activity |
|---|---|---|---|---|
| Trial 2 | 28 Aug | 0 | - | Ivermectin applied |
| 2 Sep | 5 | - | Sticky traps attached | |
| 3 Sep | 6 | - | Sticky traps collected and attached | |
| 4 Sep | 7 | - | Sticky traps collected | |
| Trial 3 | 4 Sep | 7 | 0 | Deltamethrin applied |
| 8 Sep | 11 | 4 | Sticky traps attached | |
| 9 Sep | 12 | 5 | Sticky traps collected and attached | |
| 10 Sep | 13 | 6 | Sticky traps collected and attached | |
| Flanks combed | ||||
| 11 Sep | 14 | 7 | Sticky traps collected and attached | |
| Flanks combed | ||||
| 12 Sep | 15 | 8 | Sticky traps collected | |
| Flanks combed |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murchie, A.K.; Thompson, G.M.; Clawson, S.; Brown, A.; Gordon, A.W.; Jess, S. Field Evaluation of Deltamethrin and Ivermectin Applications to Cattle on Culicoides Host-Alighting, Blood-Feeding, and Emergence. Viruses 2019, 11, 731. https://doi.org/10.3390/v11080731
Murchie AK, Thompson GM, Clawson S, Brown A, Gordon AW, Jess S. Field Evaluation of Deltamethrin and Ivermectin Applications to Cattle on Culicoides Host-Alighting, Blood-Feeding, and Emergence. Viruses. 2019; 11(8):731. https://doi.org/10.3390/v11080731
Chicago/Turabian StyleMurchie, Archie K., Geoff M. Thompson, Sam Clawson, Andrew Brown, Alan W. Gordon, and Stephen Jess. 2019. "Field Evaluation of Deltamethrin and Ivermectin Applications to Cattle on Culicoides Host-Alighting, Blood-Feeding, and Emergence" Viruses 11, no. 8: 731. https://doi.org/10.3390/v11080731