Deletions in Genes Participating in Innate Immune Response Modify the Clinical Course of Andes Orthohantavirus Infection


Abstract
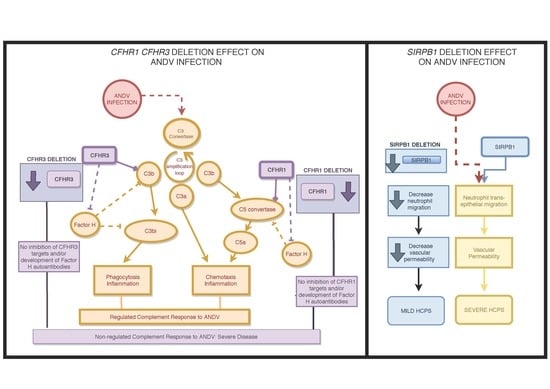
:
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Genotyping
2.2. Genotyping with Affymetrix SNP 6.0 Array
2.3. CNV Calling and Filters
2.4. Statistical Analysis
3. Results
3.1. ANDV Infected Patient Socio-Demographic Characterization
3.2. CNV Analysis
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Material Accesion Number
References
- Manigold, T.; Vial, P. Human hantavirus infections: Epidemiology, clinical features, pathogenesis and immunology. Swiss Med. Wkly. 2014, 144, w13937. [Google Scholar] [CrossRef] [PubMed]
- Vaheri, A.; Strandin, T.; Hepojoki, J.; Sironen, T.; Henttonen, H.; Mäkelä, S.; Mustonen, J. Uncovering the mysteries of hantavirus infections. Nat. Rev. Microbiol. 2013, 11, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Ferres, M.; Vial, P.; Marco, C.; Yanez, L.; Godoy, P.; Castillo, C.; Hjelle, B.; Delgado, I.; Lee, S.-J.; Mertz, G.J.; et al. Prospective evaluation of household contacts of persons with hantavirus cardiopulmonary syndrome in chile. J. Infect. Dis. 2007, 195, 1563–1571. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Valdebenito, C.; Calvo, M.; Vial, C.; Mansilla, R.; Marco, C.; Eduardo Palma, R.; Vial, P.A.; Valdivieso, F.; Mertz, G.; Ferrés, M. Person-to-Person Household and Nosocomial Transmission of Andes Hantavirus, Southern Chile, 2011. Emerg. Infect. Dis. 2014, 20, 1637–1644. [Google Scholar] [CrossRef] [PubMed]
- Vial, P.A.; Valdivieso, F.; Mertz, G.; Castillo, C.; Belmar, E.; Delgado, I.; Tapia, M.; Ferrés, M. Incubation period of hantavirus cardiopulmonary syndrome. Emerg. Infect. Dis. 2006, 12, 1271–1273. [Google Scholar] [CrossRef] [PubMed]
- Hallin, G.W.; Simpson, S.Q.; Crowell, R.E.; James, D.S.; Koster, F.T.; Mertz, G.J.; Levy, H. Cardiopulmonary manifestations of hantavirus pulmonary syndrome. Crit. Care Med. 1996, 24, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Duchin, J.S.; Koster, F.T.; Peters, C.J.; Simpson, G.L.; Tempest, B.; Zaki, S.R.; Ksiazek, T.G.; Rollin, P.E.; Nichol, S.; Umland, E.T.; et al. Hantavirus Pulmonary Syndrome: A Clinical Description of 17 Patients with a Newly Recognized Disease. New Engl. J. Med. 1994, 330, 949–955. [Google Scholar] [CrossRef] [PubMed]
- Täger, M.F.; Vial, P.C.; Castillo, C.H.; Godoy, P.M.; Hjelle, B.; Ferrés, M.G. Hantavirus Prevalence in the IX Region of Chile. Emerg. Infect. Dis. 2003, 9, 827–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toro, J.; Vega, J.D.; Khan, A.S.; Mills, J.N.; Padula, P.; Terry, W.; Yadón, Z.; Valderrama, R.; Ellis, B.A.; Pavletic, C.; et al. An outbreak of hantavirus pulmonary syndrome, Chile, 1997. Emerg. Infect. Dis. 1998, 4, 687–694. [Google Scholar] [CrossRef] [PubMed]
- Castillo, C.; Sanhueza, L.; Täger, M.; Muñoz, S.; Ossa, G.; Vial, P. Seroprevalence of antibodies against hantavirus in 10 communities of the IX Region of Chile where hantavirus infection were diagnosed. Rev. Med. Chil. 2002, 130, 251–258. [Google Scholar]
- Mertz, G.J.; Hjelle, B.; Crowley, M.; Iwamoto, G.; Tomicic, V.; Vial, P.A. Diagnosis and treatment of new world hantavirus infections. Curr. Opin. Infect. Dis. 2006, 19, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Rothman, A.L.; Kurane, I.; Montoya, J.M.; Nolte, K.B.; Norman, J.E.; Waite, D.C.; Koster, F.T.; Ennis, F.A. High levels of cytokine-producing cells in the lung tissues of patients with fatal hantavirus pulmonary syndrome. J. Infect. Dis. 1999, 179, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Perdomo-Celis, F.; Salvato, M.S.; Medina-Moreno, S.; Zapata, J.C. T-Cell Response to Viral Hemorrhagic Fevers. Vaccines 2019, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Klingström, J.; Smed-Sörensen, A.; Maleki, K.T.; Solà-Riera, C.; Ahlm, C.; Björkström, N.K.; Ljunggren, H.G. Innate and adaptive immune responses against human Puumala virus infection: Immunopathogenesis and suggestions for novel treatment strategies for severe hantavirus-associated syndromes. J. Intern. Med. 2019, 285, 510–523. [Google Scholar] [CrossRef] [PubMed]
- Maleki, K.T.; García, M.; Iglesias, A.; Alonso, D.; Ciancaglini, M.; Hammar, U.; Ljunggren, H.-G.; Schierloh, P.; Martínez, V.P.; Klingström, J. Serum Markers Associated with Severity and Outcome of Hantavirus Pulmonary Syndrome. J. Infect. Dis. 2019, 219, 1832–1840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonsson, C.B.; Hooper, J.; Mertz, G. Treatment of hantavirus pulmonary syndrome. Antivir. Res. 2008, 78, 162–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angulo, J.; Pino, K.; Echeverría-Chagas, N.; Marco, C.; Martínez-Valdebenito, C.; Galeno, H.; Villagra, E.; Vera, L.; Lagos, N.; Becerra, N.; et al. Association of Single-Nucleotide Polymorphisms inIL28B, but NotTNF-α, with Severity of Disease Caused by Andes Virus. Clin. Infect. Dis. 2015, 61, e62–e69. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.L.; Lai, J.H.; Zhu, Y.; Zhang, H.B.; Li, C.; Wang, J.P.; Li, Y.M.; Yang, A.G.; Jin, B.Q. Genetic susceptibility to haemorrhagic fever with renal syndrome caused by Hantaan virus in Chinese Han population. Int. J. Immunogenet. 2009, 36, 227–229. [Google Scholar] [CrossRef] [PubMed]
- Korva, M.; Saksida, A.; Kunilo, S.; Vidan Jeras, B.; Avsic-Zupanc, T. HLA-associated hemorrhagic fever with renal syndrome disease progression in slovenian patients. Clin. Vaccine Immunol. 2011, 18, 1435–1440. [Google Scholar] [CrossRef] [PubMed]
- Laine, O.; Joutsi-Korhonen, L.; Mäkelä, S.; Mikkelsson, J.; Pessi, T.; Tuomisto, S.; Huhtala, H.; Libraty, D.; Vaheri, A.; Karhunen, P.; et al. Polymorphisms of PAI-1 and platelet GP Ia may associate with impairment of renal function and thrombocytopenia in Puumala hantavirus infection. Thromb. Res. 2012, 129, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.A.; Donadi, E.A.; Campos, G.M.; de Figueiredo, G.G.; Saggioro, F.P.; Badra, S.J.; Deghaide, N.H.S.; Figueiredo, L.T.M. Polymorphisms in human leukocyte antigens, human platelet antigens, and cytokine genes in hantavirus cardiopulmonary syndrome patients from Ribeirão Preto, Brazil. J. Med. Virol. 2014, 86, 1962–1970. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, C.P.; Vial, C.P.A.; Ferrés, G.M.; Godoy, M.P.; Culza, A.V.; Marco, C.C.; Castillo, H.C.; Umaña, C.M.E.; Rothhammer, E.F.; Llop, R.E. Genetic susceptibility to Andes Hantavirus: Association between severity of disease and HLA alíeles in Chilean patients. Rev. Chilena Infectol. 2007, 24, 351–359. [Google Scholar]
- McCarroll, S.A.; Altshuler, D.M. Copy-number variation and association studies of human disease. Nat. Genet. 2007, 39, S37–S42. [Google Scholar] [CrossRef] [PubMed]
- Hollox, E.J.; Hoh, B.-P. Human gene copy number variation and infectious disease. Hum. Genet. 2014, 133, 1217–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Copy Number Variation. Available online: https://www.nature.com/scitable/topicpage/copy-number-variation-445 (accessed on 12 July 2019).
- May, J.; Evans, J.A.; Timmann, C.; Ehmen, C.; Busch, W.; Thye, T.; Agbenyega, T.; Horstmann, R.D. Hemoglobin variants and disease manifestations in severe falciparum malaria. JAMA 2007, 297, 2220–2226. [Google Scholar] [CrossRef]
- Gonzalez, E.; Kulkarni, H.; Bolivar, H.; Mangano, A.; Sanchez, R.; Catano, G.; Nibbs, R.J.; Freedman, B.I.; Quinones, M.P.; Bamshad, M.J.; et al. The influence of CCL3L1 gene-containing segmental duplications on HIV-1/AIDS susceptibility. Science 2005, 307, 1434–1440. [Google Scholar] [CrossRef]
- Wang, K.; Li, M.; Hadley, D.; Liu, R.; Glessner, J.; Grant, S.F.A.; Hakonarson, H.; Bucan, M. PennCNV: An integrated hidden Markov model designed for high-resolution copy number variation detection in whole-genome SNP genotyping data. Genome Res. 2007, 17, 1665–1674. [Google Scholar] [CrossRef] [Green Version]
- Diskin, S.J.; Li, M.; Hou, C.; Yang, S.; Glessner, J.; Hakonarson, H.; Bucan, M.; Maris, J.M.; Wang, K. Adjustment of genomic waves in signal intensities from whole-genome SNP genotyping platforms. Nucleic Acids Res. 2008, 36, e126. [Google Scholar] [CrossRef]
- León, L.E.; Benavides, F.; Espinoza, K.; Vial, C.; Alvarez, P.; Palomares, M.; Lay-Son, G.; Miranda, M.; Repetto, G.M. Partial microduplication in the histone acetyltransferase complex member KANSL1 is associated with congenital heart defects in 22q11.2 microdeletion syndrome patients. Sci. Rep. 2017, 7, 1795. [Google Scholar] [CrossRef]
- Glessner, J.T.; Li, J.; Hakonarson, H. ParseCNV integrative copy number variation association software with quality tracking. Nucleic Acids Res. 2013, 41, e64. [Google Scholar] [CrossRef]
- Hinrichs, A.S.; Karolchik, D.; Baertsch, R.; Barber, G.P.; Bejerano, G.; Clawson, H.; Diekhans, M.; Furey, T.S.; Harte, R.A.; Hsu, F.; et al. The UCSC Genome Browser Database: Update 2006. Nucleic Acids Res. 2006, 34, D590–D598. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Young, J.M.; Endicott, R.M.; Parghi, S.S.; Walker, M.; Kidd, J.M.; Trask, B.J. Extensive copy-number variation of the human olfactory receptor gene family. Am. J. Hum. Genet. 2008, 83, 228–242. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, P.F.; Skerka, C. Complement regulators and inhibitory proteins. Nat. Rev. Immunol. 2009, 9, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Huik, K.; Sadam, M.; Karki, T.; Avi, R.; Krispin, T.; Paap, P.; Rüütel, K.; Uusküla, A.; Talu, A.; Abel-Ollo, K.; et al. CCL3L1 copy number is a strong genetic determinant of HIV seropositivity in Caucasian intravenous drug users. J. Infect. Dis. 2010, 201, 730–739. [Google Scholar] [CrossRef] [PubMed]
- Grünhage, F.; Nattermann, J.; Gressner, O.A.; Wasmuth, H.E.; Hellerbrand, C.; Sauerbruch, T.; Spengler, U.; Lammert, F. Lower copy numbers of the chemokine CCL3L1 gene in patients with chronic hepatitis C. J. Hepatol. 2010, 52, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, X.; Zou, G.-Z.; Gao, Y.-F.; Ye, J. Association between TLR7 copy number variations and hepatitis B virus infection outcome in Chinese. World J. Gastroenterol. 2017, 23, 1602. [Google Scholar] [CrossRef] [PubMed]
- Skerka, C.; Chen, Q.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement factor H related proteins (CFHRs). Mol. Immunol. 2013, 56, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Ward, P.A. Functions of C5a receptors. J. Mol. Med. 2009, 87, 375–378. [Google Scholar] [CrossRef]
- Díaz-Guillén, M.A.; Rodríguez de Córdoba, S.; Heine-Suñer, D. A radiation hybrid map of complement factor H and factor H-related genes. Immunogenetics 1999, 49, 549–552. [Google Scholar] [CrossRef]
- Heinen, S.; Hartmann, A.; Lauer, N.; Wiehl, U.; Dahse, H.-M.; Schirmer, S.; Gropp, K.; Enghardt, T.; Wallich, R.; Hälbich, S.; et al. Factor H-related protein 1 (CFHR-1) inhibits complement C5 convertase activity and terminal complex formation. Blood 2009, 114, 2439–2447. [Google Scholar] [CrossRef] [PubMed]
- Józsi, M.; Schneider, A.E.; Kárpáti, É.; Sándor, N. Complement factor H family proteins in their non-canonical role as modulators of cellular functions. Semin. Cell Dev. Biol. 2019, 85, 122–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zipfel, P.F.; Edey, M.; Heinen, S.; Józsi, M.; Richter, H.; Misselwitz, J.; Hoppe, B.; Routledge, D.; Strain, L.; Hughes, A.E.; et al. Deletion of Complement Factor H–Related Genes CFHR1 and CFHR3 Is Associated with Atypical Hemolytic Uremic Syndrome. PLoS Genet. 2007, 3, e41. [Google Scholar] [CrossRef] [PubMed]
- Józsi, M.; Licht, C.; Strobel, S.; Zipfel, S.L.H.; Richter, H.; Heinen, S.; Zipfel, P.F.; Skerka, C. Factor H autoantibodies in atypical hemolytic uremic syndrome correlate with CFHR1/CFHR3 deficiency. Blood 2008, 111, 1512–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sane, J.; Laine, O.; Mäkelä, S.; Paakkala, A.; Jarva, H.; Mustonen, J.; Vapalahti, O.; Meri, S.; Vaheri, A. Complement activation in Puumala hantavirus infection correlates with disease severity. Ann. Med. 2012, 44, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Paakkala, A.; Mustonen, J.; Viander, M.; Huhtala, H.; Pasternack, A. Complement activation in nephropathia epidemica caused by Puumala hantavirus. Clin. Nephrol. 2000, 53, 424–431. [Google Scholar] [PubMed]
- Liu, Y.; Soto, I.; Tong, Q.; Chin, A.; Bühring, H.-J.; Wu, T.; Zen, K.; Parkos, C.A. SIRPβ1 Is Expressed as a Disulfide-linked Homodimer in Leukocytes and Positively Regulates Neutrophil Transepithelial Migration. J. Biol. Chem. 2005, 280, 36132–36140. [Google Scholar] [CrossRef] [PubMed]
- Barclay, A.N.; Brown, M.H. The SIRP family of receptors and immune regulation. Nat. Rev. Immunol. 2006, 6, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Strandin, T.; Mäkelä, S.; Mustonen, J.; Vaheri, A. Neutrophil Activation in Acute Hemorrhagic Fever with Renal Syndrome Is Mediated by Hantavirus-Infected Microvascular Endothelial Cells. Front. Immunol. 2018, 9, 2098. [Google Scholar] [CrossRef]
- Koma, T.; Yoshimatsu, K.; Nagata, N.; Sato, Y.; Shimizu, K.; Yasuda, S.P.; Amada, T.; Nishio, S.; Hasegawa, H.; Arikawa, J. Neutrophil Depletion Suppresses Pulmonary Vascular Hyperpermeability and Occurrence of Pulmonary Edema Caused by Hantavirus Infection in C.B-17 SCID Mice. J. Virol. 2014, 88, 7178–7188. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Severe (n = 107) | Mild (n = 88) | p-Value | ||
|---|---|---|---|---|
| Age (years), Mean | 38.12 | 35.33 | 0.99 a | |
| Gender, n (%) | Man | 71 (66.4) | 62 (70.5) | 0.64 b |
| Woman | 36 (33.6) | 26 (29.5) | ||
| Total | 107 (100) | 88 (100) |
| CNV Number | Genome Coordinates (GRCh38/hg38) | OR (95% CI) | p-Value | Gain/Loss | Severe (n = 60) | Mild (n = 51) | * Distance to Closest Gene (bp) | Putative Affected Gene | Cytoband |
|---|---|---|---|---|---|---|---|---|---|
| CNVs enriched in severe patients | |||||||||
| CNV1 | chr11: 18940271–18940271 | 4.37 (1.10–25.44) | 0.028 | Loss | 13 | 3 | 5269 | MRGPRX1 | 11p15.1 |
| CNV2 | chr1: 196769493–196789029 | 3.31 (1.04–12.55) | 0.029 | Loss | 16 | 5 | 0 | CFHR1, CFHR2, CFHR3 | 1q31.3 |
| CNV3 | chr11: 4950775–4955384 | 3.31 (1.04–12.55) | 0.029 | Gain | 16 | 5 | 0 | OR51A2 | 11p15.4 |
| CNVs enriched in mild patients | |||||||||
| CNV4 | chr20: 1599742–1601561 | 0.3 (0.12–0.76) | 0.007 | Loss | 12 | 23 | 0 | SIRPB1 | 20p13 |
| CNV5 | chr14: 19742838–19939483 | 0.19 (0.03–0.8) | 0.010 | Gain | 3 | 11 | 0 | OR4K1, OR4K2, OR4K5, OR4M1, OR4N2, OR4Q3 | 14q11.2 |
| CNV6 | chr2: 146106836–146109366 | 0.16 (0.01–0.85) | 0.022 | Gain | 2 | 9 | 478,078 | LOC728773 | 2q22.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro, G.E.; Leon, L.E.; Perez, R.; Cuiza, A.; Vial, P.A.; Ferres, M.; Mertz, G.J.; Vial, C. Deletions in Genes Participating in Innate Immune Response Modify the Clinical Course of Andes Orthohantavirus Infection. Viruses 2019, 11, 680. https://doi.org/10.3390/v11080680
Ribeiro GE, Leon LE, Perez R, Cuiza A, Vial PA, Ferres M, Mertz GJ, Vial C. Deletions in Genes Participating in Innate Immune Response Modify the Clinical Course of Andes Orthohantavirus Infection. Viruses. 2019; 11(8):680. https://doi.org/10.3390/v11080680
Chicago/Turabian StyleRibeiro, Grazielle Esteves, Luis Edgardo Leon, Ruth Perez, Analia Cuiza, Pablo Agustin Vial, Marcela Ferres, Gregory J. Mertz, and Cecilia Vial. 2019. "Deletions in Genes Participating in Innate Immune Response Modify the Clinical Course of Andes Orthohantavirus Infection" Viruses 11, no. 8: 680. https://doi.org/10.3390/v11080680