A DNA Prime Immuno-Potentiates a Modified Live Vaccine against the Porcine Reproductive and Respiratory Syndrome Virus but Does Not Improve Heterologous Protection

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Material and Methods
2.1. Antibodies (Abs)
2.2. Viruses
2.3. DNA Vectors
2.4. Pig Studies, Ethic and Authorizations
2.5. In Vivo Gene Transfer
2.6. Detection of pLuc Expression
2.7. Detection of pmCherry Expression
2.8. Detection of Inflammatory Cytokines in Pig Transfected Skin
2.9. Immunization of Pigs and Infectious Challenge
2.10. Nasal Swab and Serum Collections
2.11. Overlapping Peptides
2.12. IFNγ Detection by ELISPOTS
2.13. Detection of Anti-N IgG and Anti-Envelope IgG
2.14. Detection of Neutralizing Antibodies (NAbs)
2.15. Viral Detection by Specific qRT-PCR
2.16. Statistical and Correlation Analyses
3. Results
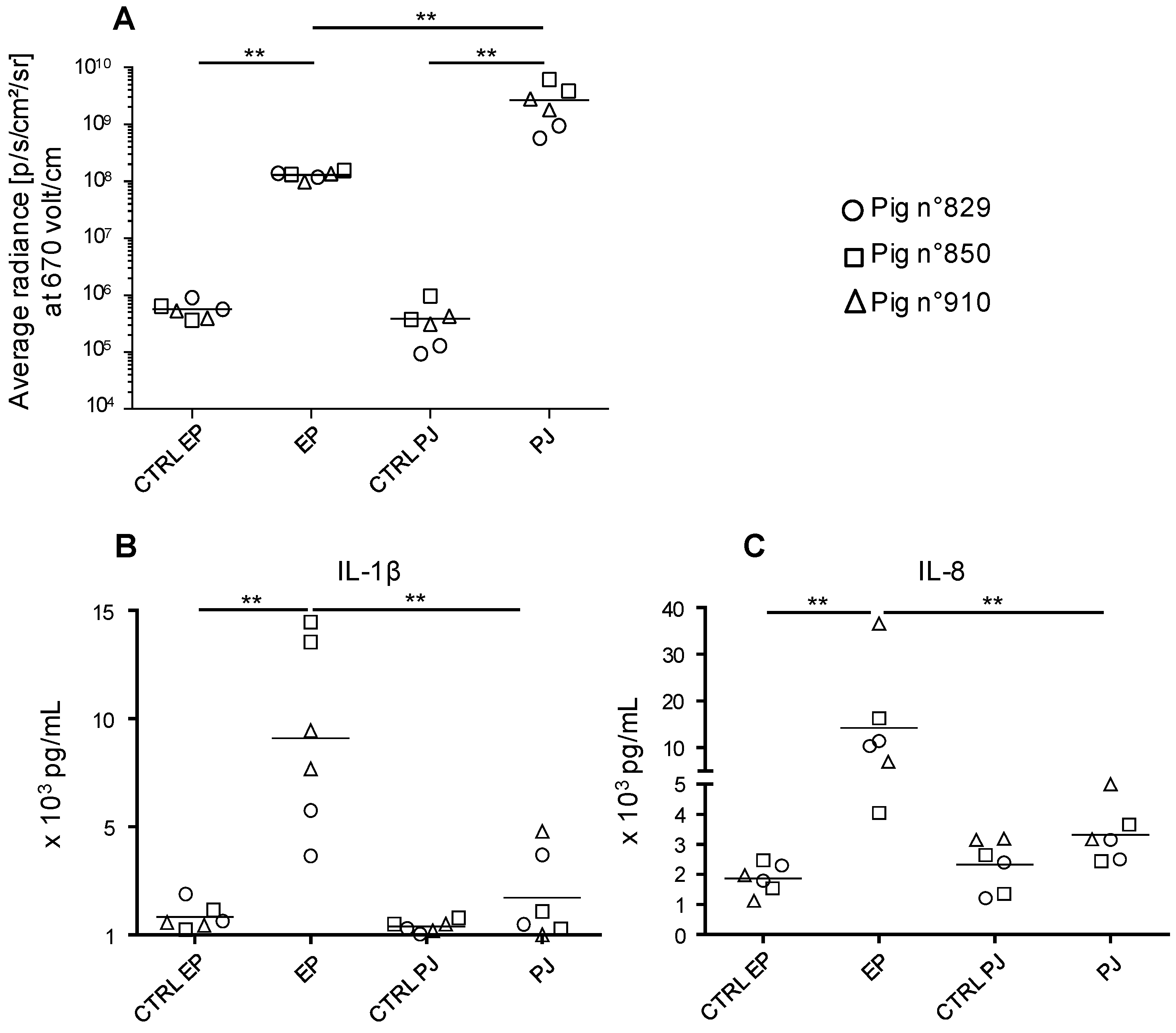
3.1. Transduction Efficacy of Pig Skin Cells upon DNA Delivery with a Needle-Free High Pressure Jet Injection (Pharmajet Tropis-ID device, PJ) Versus Surface EP
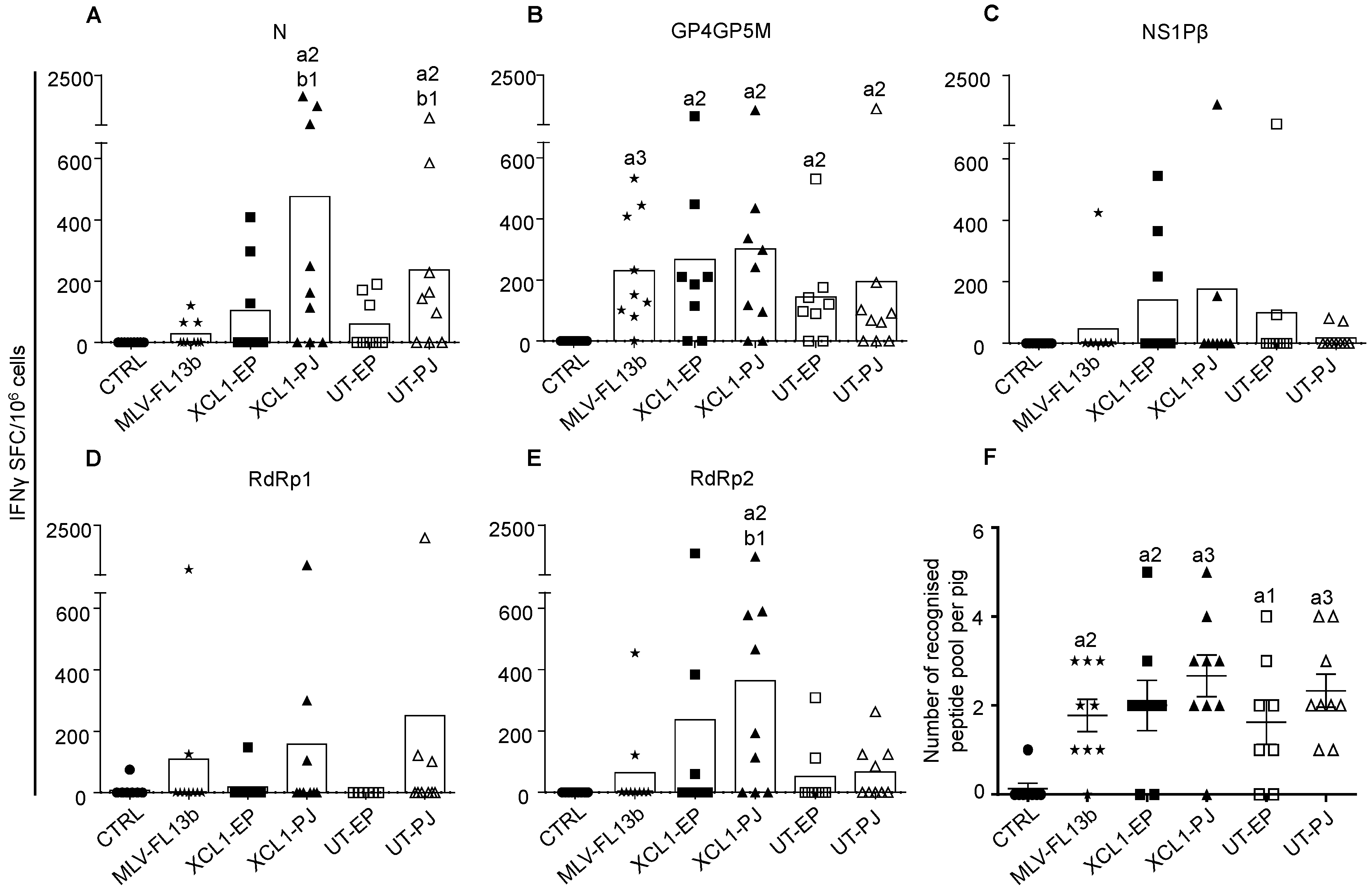
3.2. IFNγ T-Cell Responses Induced by a Prime-Boost DNA-MLV-FL13b Strategy with DNA Encoding PRRSV-AGs Targeted or Not to XCR1 and Delivered with EP or PJ
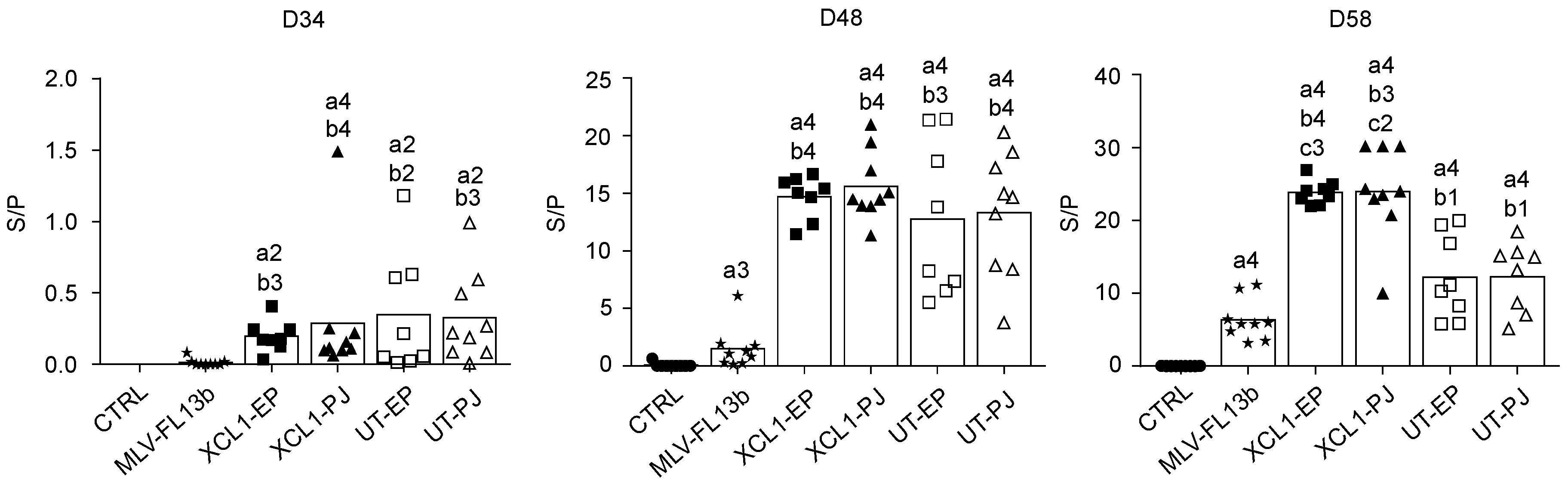
3.3. Antibody Responses Induced by the Prime-Boost DNA-MLV-FL13b Strategy
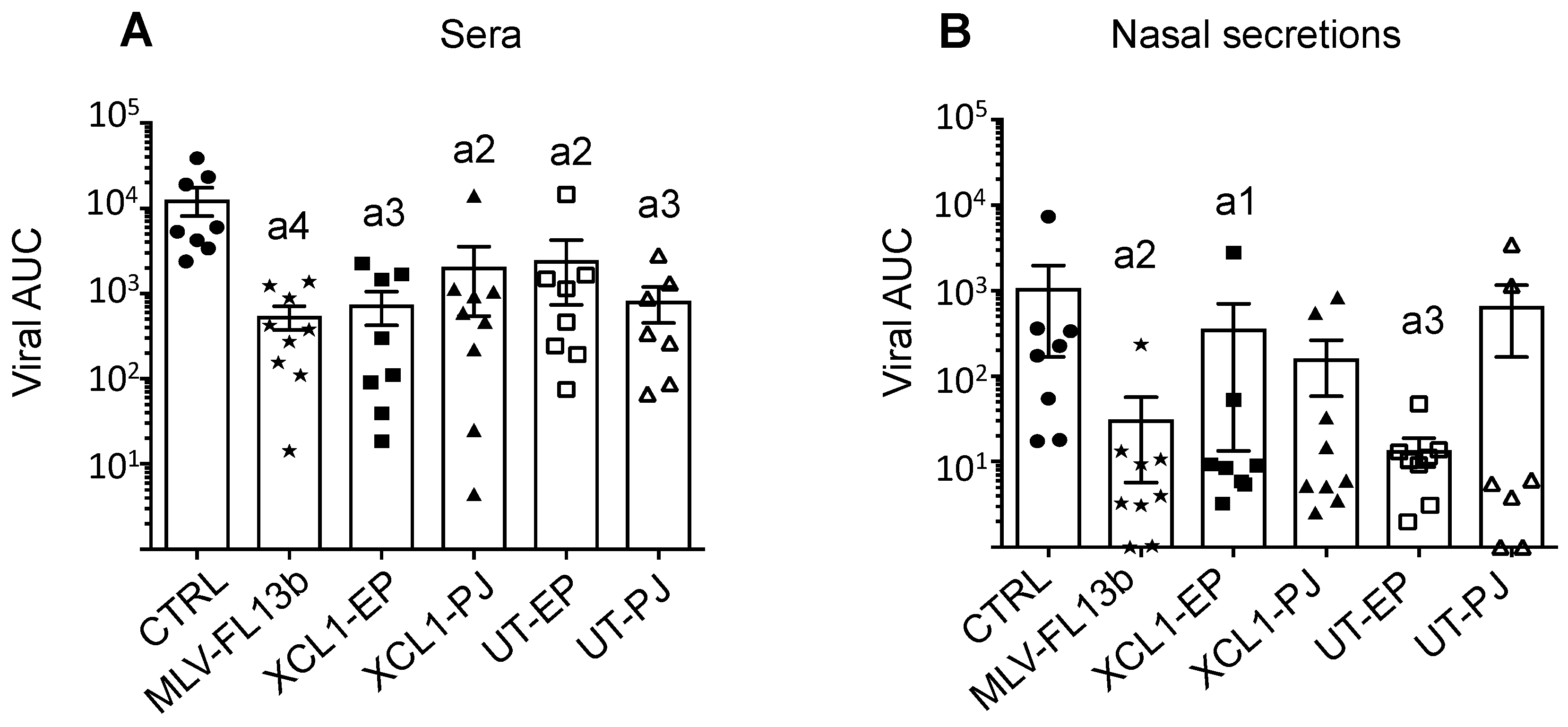
3.4. Protection Induced by the Prime-Boost DNA-MLV-FL13b Strategy against a Heterologous Challenge with FL07
3.5. Lack of Immune Correlates of Protection Induced by the MLV and Prime-Boost DNA-MLV-FL13b Strategies against the Heterologous FL07 Challenge
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Butler, J.E.; Lager, K.M.; Golde, W.; Faaberg, K.S.; Sinkora, M.; Loving, C.; Zhang, Y.I. Porcine reproductive and respiratory syndrome (PRRS): An immune dysregulatory pandemic. Immunol. Res. 2014, 59, 81–108. [Google Scholar] [CrossRef] [PubMed]
- Lunney, J.K.; Fang, Y.; Ladinig, A.; Chen, N.; Li, Y.; Rowland, B.; Renukaradhya, G.J. Porcine reproductive and respiratory syndrome virus (PRRSV): Pathogenesis and interaction with the immune system. Annu. Rev. Anim. Biosci. 2016, 4, 129–154. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, J.H.; Lauck, M.; Bailey, A.L.; Shchetinin, A.M.; Vishnevskaya, T.V.; Bao, Y.; Ng, T.F.; LeBreton, M.; Schneider, B.S.; Gillis, A.; et al. Reorganization and expansion of the nidoviral family arteriviridae. Arch. Virol. 2016, 161, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lam, T.T.; Hon, C.C.; Hui, R.K.; Faaberg, K.S.; Wennblom, T.; Murtaugh, M.P.; Stadejek, T.; Leung, F.C. Molecular epidemiology of prrsv: A phylogenetic perspective. Virus Res. 2010, 154, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Renukaradhya, G.J.; Meng, X.J.; Calvert, J.G.; Roof, M.; Lager, K.M. Live porcine reproductive and respiratory syndrome virus vaccines: Current status and future direction. Vaccine 2015, 33, 4069–4080. [Google Scholar] [CrossRef] [PubMed]
- Lyoo, Y.S. Porcine reproductive and respiratory syndrome virus vaccine does not fit in classical vaccinology. Clin. Exp. Vaccine Res. 2015, 4, 159–165. [Google Scholar] [CrossRef]
- Martelli, P.; Gozio, S.; Ferrari, L.; Rosina, S.; De Angelis, E.; Quintavalla, C.; Bottarelli, E.; Borghetti, P. Efficacy of a modified live porcine reproductive and respiratory syndrome virus (PRRSV) vaccine in pigs naturally exposed to a heterologous european (Italian cluster) field strain: Clinical protection and cell-mediated immunity. Vaccine 2009, 27, 3788–3799. [Google Scholar] [CrossRef]
- Mokhtar, H.; Eck, M.; Morgan, S.B.; Essler, S.E.; Frossard, J.P.; Ruggli, N.; Graham, S.P. Proteome-wide screening of the european porcine reproductive and respiratory syndrome virus reveals a broad range of t cell antigen reactivity. Vaccine 2014, 32, 6828–6837. [Google Scholar] [CrossRef]
- Bernelin-Cottet, C.; Urien, C.; Studsrub, E.; Jakob, V.; Bouguyon, E.; Bordet, E.; Barc, C.; Boulesteix, O.; Contreras, V.; Barnier-Quer, C.; et al. A DNA-modified live vaccine prime–boost strategy broadens the t-cell response and enhances the antibody response against the porcine reproductive and respiratory syndrome virus. Viruses 2019, 11, 551. [Google Scholar] [CrossRef]
- Crozat, K.; Tamoutounour, S.; Vu Manh, T.P.; Fossum, E.; Luche, H.; Ardouin, L.; Guilliams, M.; Azukizawa, H.; Bogen, B.; Malissen, B.; et al. Cutting edge: Expression of xcr1 defines mouse lymphoid-tissue resident and migratory dendritic cells of the cd8α+ type. J. Immunol. 2011, 187, 4411–4415. [Google Scholar] [CrossRef]
- Grodeland, G.; Fredriksen, A.B.; Loset, G.A.; Vikse, E.; Fugger, L.; Bogen, B. Antigen targeting to human hla class ii molecules increases efficacy of DNA vaccination. J. Immunol. 2016, 197, 3575–3585. [Google Scholar] [CrossRef] [PubMed]
- Frydas, I.S.; Trus, I.; Kvisgaard, L.K.; Bonckaert, C.; Reddy, V.R.; Li, Y.; Larsen, L.E.; Nauwynck, H.J. Different clinical, virological, serological and tissue tropism outcomes of two new and one old belgian type 1 subtype 1 porcine reproductive and respiratory virus (PRRSV) isolates. Vet. Res. 2015, 46, 37. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, S.; Pineyro, P.; Tian, D.; Overend, C.; Yugo, D.M.; Matzinger, S.R.; Rogers, A.J.; Haac, M.E.; Cao, Q.; Heffron, C.L.; et al. In vivo targeting of porcine reproductive and respiratory syndrome virus antigen through porcine dc-sign to dendritic cells elicits antigen-specific cd4t cell immunity in pigs. Vaccine 2014, 32, 6768–6775. [Google Scholar] [CrossRef] [PubMed]
- Bernelin-Cottet, C.; Urien, C.; McCaffrey, J.; Collins, D.; Donadei, A.; McDaid, D.; Jakob, V.; Barnier-Quer, C.; Collin, N.; Bouguyon, E.; et al. Assessing the immunogenic properties of DNA vaccines delivered with nanoparticles, electroporation and dissolvable microneedle patches in pig skin. J. Control. Release. under review.
- Fredriksen, A.B.; Bogen, B. Chemokine-idiotype fusion DNA vaccines are potentiated by bivalency and xenogeneic sequences. Blood 2007, 110, 1797–1805. [Google Scholar] [CrossRef] [PubMed]
- Deloizy, C.; Bouguyon, E.; Fossum, E.; Sebo, P.; Osicka, R.; Bole, A.; Pierres, M.; Biacchesi, S.; Dalod, M.; Bogen, B.; et al. Expanding the tools for identifying mononuclear phagocyte subsets in swine: Reagents to porcine cd11c and xcr1. Dev. Comp. Immunol. 2016, 65, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Renier, N.; Adams, E.L.; Kirst, C.; Wu, Z.; Azevedo, R.; Kohl, J.; Autry, A.E.; Kadiri, L.; Umadevi Venkataraju, K.; Zhou, Y.; et al. Mapping of brain activity by automated volume analysis of immediate early genes. Cell 2016, 165, 1789–1802. [Google Scholar] [CrossRef] [PubMed]
- Maisonnasse, P.; Bouguyon, E.; Piton, G.; Ezquerra, A.; Urien, C.; Deloizy, C.; Bourge, M.; Leplat, J.J.; Simon, G.; Chevalier, C.; et al. The respiratory dc/macrophage network at steady-state and upon influenza infection in the swine biomedical model. Mucosal Immunol. 2016, 9, 835–849. [Google Scholar] [CrossRef]
- Charpin, C.; Mahe, S.; Keranflec’h, A.; Belloc, C.; Cariolet, R.; Le Potier, M.F.; Rose, N. Infectiousness of pigs infected by the porcine reproductive and respiratory syndrome virus (PRRSV) is time-dependent. Vet. Res. 2012, 43, 69. [Google Scholar] [CrossRef] [PubMed]
- Biernacka, K.; Podgorska, K.; Tyszka, A.; Stadejek, T. Comparison of six commercial elisas for the detection of antibodies against porcine reproductive and respiratory syndrome virus (PRRSV) in field serum samples. Res. Vet. Sci. 2018, 121, 40–45. [Google Scholar] [CrossRef]
- Savard, C.; Alvarez, F.; Provost, C.; Chorfi, Y.; D’Allaire, S.; Benoit-Biancamano, M.O.; Gagnon, C.A. Efficacy of fostera prrs modified live virus vaccine against a canadian heterologous virulent field strain of porcine reproductive and respiratory syndrome virus. Can. J. Vet. Res. 2016, 80, 1–11. [Google Scholar] [PubMed]
- Vanhee, M.; Costers, S.; Van Breedam, W.; Geldhof, M.F.; Van Doorsselaere, J.; Nauwynck, H.J. A variable region in gp4 of european-type porcine reproductive and respiratory syndrome virus induces neutralizing antibodies against homologous but not heterologous virus strains. Viral. Immunol. 2010, 23, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Caminschi, I.; Maraskovsky, E.; Heath, W.R. Targeting dendritic cells in vivo for cancer therapy. Front. Immunol. 2012, 3, 13. [Google Scholar] [CrossRef] [PubMed]
- Neubert, K.; Lehmann, C.H.; Heger, L.; Baranska, A.; Staedtler, A.M.; Buchholz, V.R.; Yamazaki, S.; Heidkamp, G.F.; Eissing, N.; Zebroski, H.; et al. Antigen delivery to CD11c+ CD8− dendritic cells induces protective immune responses against experimental melanoma in mice in vivo. J. Immunol. 2014, 192, 5830–5838. [Google Scholar] [CrossRef] [PubMed]
- Tregoning, J.S.; Kinnear, E. Using plasmids as DNA vaccines for infectious diseases. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef]
- Luke, J.M.; Simon, G.G.; Soderholm, J.; Errett, J.S.; August, J.T.; Gale, M., Jr.; Hodgson, C.P.; Williams, J.A. Coexpressed rig-i agonist enhances humoral immune response to influenza virus DNA vaccine. J. Virol. 2011, 85, 1370–1383. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.R.; Rahe, M.C.; Gray, D.K.; Martins, K.V.; Murtaugh, M.P. Porcine reproductive and respiratory syndrome virus neutralizing antibodies provide in vivo cross-protection to prrsv1 and prrsv2 viral challenge. Virus Res. 2018, 248, 13–23. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, H.; Ma, Z.; Wang, Y.; Feng, W.H. Ctla4 mediated targeting enhances immunogenicity against prrsv in a DNA prime/killed virus boost strategy. Vet. Immunol. Immunopathol. 2013, 154, 121–128. [Google Scholar] [CrossRef]
- Yu, M.; Qiu, Y.; Chen, J.; Jiang, W. Enhanced humoral and cellular immune responses to prrs virus gp5 glycoprotein by DNA prime-adenovirus boost vaccination in mice. Virus Genes 2016, 52, 228–234. [Google Scholar] [CrossRef]
- Suradhat, S.; Wongyanin, P.; Kesdangsakonwut, S.; Teankum, K.; Lumyai, M.; Triyarach, S.; Thanawongnuwech, R. A novel DNA vaccine for reduction of prrsv-induced negative immunomodulatory effects: A proof of concept. Vaccine 2015, 33, 3997–4003. [Google Scholar] [CrossRef]
- Rahe, M.C.; Murtaugh, M.P. Mechanisms of adaptive immunity to porcine reproductive and respiratory syndrome virus. Viruses 2017, 9, 148. [Google Scholar] [CrossRef]
- Renson, P.; Rose, N.; Le Dimna, M.; Mahe, S.; Keranflec’h, A.; Paboeuf, F.; Belloc, C.; Le Potier, M.F.; Bourry, O. Dynamic changes in bronchoalveolar macrophages and cytokines during infection of pigs with a highly or low pathogenic genotype 1 prrsv strain. Vet. Res. 2017, 48, 15. [Google Scholar] [CrossRef]
- Rose, N.; Renson, P.; Andraud, M.; Paboeuf, F.; Le Potier, M.F.; Bourry, O. Porcine reproductive and respiratory syndrome virus (PRRSV) modified-live vaccine reduces virus transmission in experimental conditions. Vaccine 2015, 33, 2493–2499. [Google Scholar] [CrossRef]
- Mokhtar, H.; Biffar, L.; Somavarapu, S.; Frossard, J.P.; McGowan, S.; Pedrera, M.; Strong, R.; Edwards, J.C.; Garcia-Duran, M.; Rodriguez, M.J.; et al. Evaluation of hydrophobic chitosan-based particulate formulations of porcine reproductive and respiratory syndrome virus vaccine candidate t cell antigens. Vet. Microbiol. 2017, 209, 66–74. [Google Scholar] [CrossRef]
- Mokhtar, H.; Pedrera, M.; Frossard, J.P.; Biffar, L.; Hammer, S.E.; Kvisgaard, L.K.; Larsen, L.E.; Stewart, G.R.; Somavarapu, S.; Steinbach, F.; et al. The non-structural protein 5 and matrix protein are antigenic targets of t cell immunity to genotype 1 porcine reproductive and respiratory syndrome viruses. Front. Immunol. 2016, 7, 40. [Google Scholar] [CrossRef]
- Suradhat, S.; Wongyanin, P.; Sirisereewan, C.; Nedumpun, T.; Lumyai, M.; Triyarach, S.; Chaturavittawong, D.; Paphavasit, T.; Panyatong, R.; Thanawongnuwech, R. Transdermal delivery of plasmid encoding truncated nucleocapsid protein enhanced prrsv-specific immune responses. Vaccine 2015, 34, 609–615. [Google Scholar] [CrossRef]
- Diaz, I.; Darwich, L.; Pappaterra, G.; Pujols, J.; Mateu, E. Immune responses of pigs after experimental infection with a european strain of porcine reproductive and respiratory syndrome virus. J. Gen. Virol. 2005, 86, 1943–1951. [Google Scholar] [CrossRef]
- Meier, W.A.; Galeota, J.; Osorio, F.A.; Husmann, R.J.; Schnitzlein, W.M.; Zuckermann, F.A. Gradual development of the interferon-gamma response of swine to porcine reproductive and respiratory syndrome virus infection or vaccination. Virology 2003, 309, 18–31. [Google Scholar] [CrossRef]
- Salguero, F.J.; Frossard, J.P.; Rebel, J.M.; Stadejek, T.; Morgan, S.B.; Graham, S.P.; Steinbach, F. Host-pathogen interactions during porcine reproductive and respiratory syndrome virus 1 infection of piglets. Virus Res. 2015, 202, 135–143. [Google Scholar] [CrossRef]
- Weesendorp, E.; Morgan, S.; Stockhofe-Zurwieden, N.; Popma-De Graaf, D.J.; Graham, S.P.; Rebel, J.M. Comparative analysis of immune responses following experimental infection of pigs with european porcine reproductive and respiratory syndrome virus strains of differing virulence. Vet. Microbiol. 2013, 163, 1–12. [Google Scholar] [CrossRef]
- Morgan, S.B.; Graham, S.P.; Salguero, F.J.; Sanchez Cordon, P.J.; Mokhtar, H.; Rebel, J.M.; Weesendorp, E.; Bodman-Smith, K.B.; Steinbach, F.; Frossard, J.P. Increased pathogenicity of european porcine reproductive and respiratory syndrome virus is associated with enhanced adaptive responses and viral clearance. Vet. Microbiol. 2013, 163, 13–22. [Google Scholar] [CrossRef]
- Park, C.; Seo, H.W.; Han, K.; Kang, I.; Chae, C. Evaluation of the efficacy of a new modified live porcine reproductive and respiratory syndrome virus (PRRSV) vaccine (Fostera PRRS) against heterologous prrsv challenge. Vet. Microbiol. 2014, 172, 432–442. [Google Scholar] [CrossRef]
- Diaz, I.; Darwich, L.; Pappaterra, G.; Pujols, J.; Mateu, E. Different european-type vaccines against porcine reproductive and respiratory syndrome virus have different immunological properties and confer different protection to pigs. Virology 2006, 351, 249–259. [Google Scholar] [CrossRef]
- Balasch, M.; Fort, M.; Taylor, L.P.; Calvert, J.G. Vaccination of 1-day-old pigs with a porcine reproductive and respiratory syndrome virus (PRRSV) modified live attenuated virus vaccine is able to overcome maternal immunity. Porc. Health Manag. 2018, 4, 25. [Google Scholar] [CrossRef]
- Mukherjee, S.; Dowd, K.A.; Manhart, C.J.; Ledgerwood, J.E.; Durbin, A.P.; Whitehead, S.S.; Pierson, T.C. Mechanism and significance of cell type-dependent neutralization of flaviviruses. J. Virol. 2014, 88, 7210–7220. [Google Scholar] [CrossRef]
- Tian, D.; Cao, D.; Lynn Heffron, C.; Yugo, D.M.; Rogers, A.J.; Overend, C.; Matzinger, S.R.; Subramaniam, S.; Opriessnig, T.; LeRoith, T.; et al. Enhancing heterologous protection in pigs vaccinated with chimeric porcine reproductive and respiratory syndrome virus containing the full-length sequences of shuffled structural genes of multiple heterologous strains. Vaccine 2017, 35, 2427–2434. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
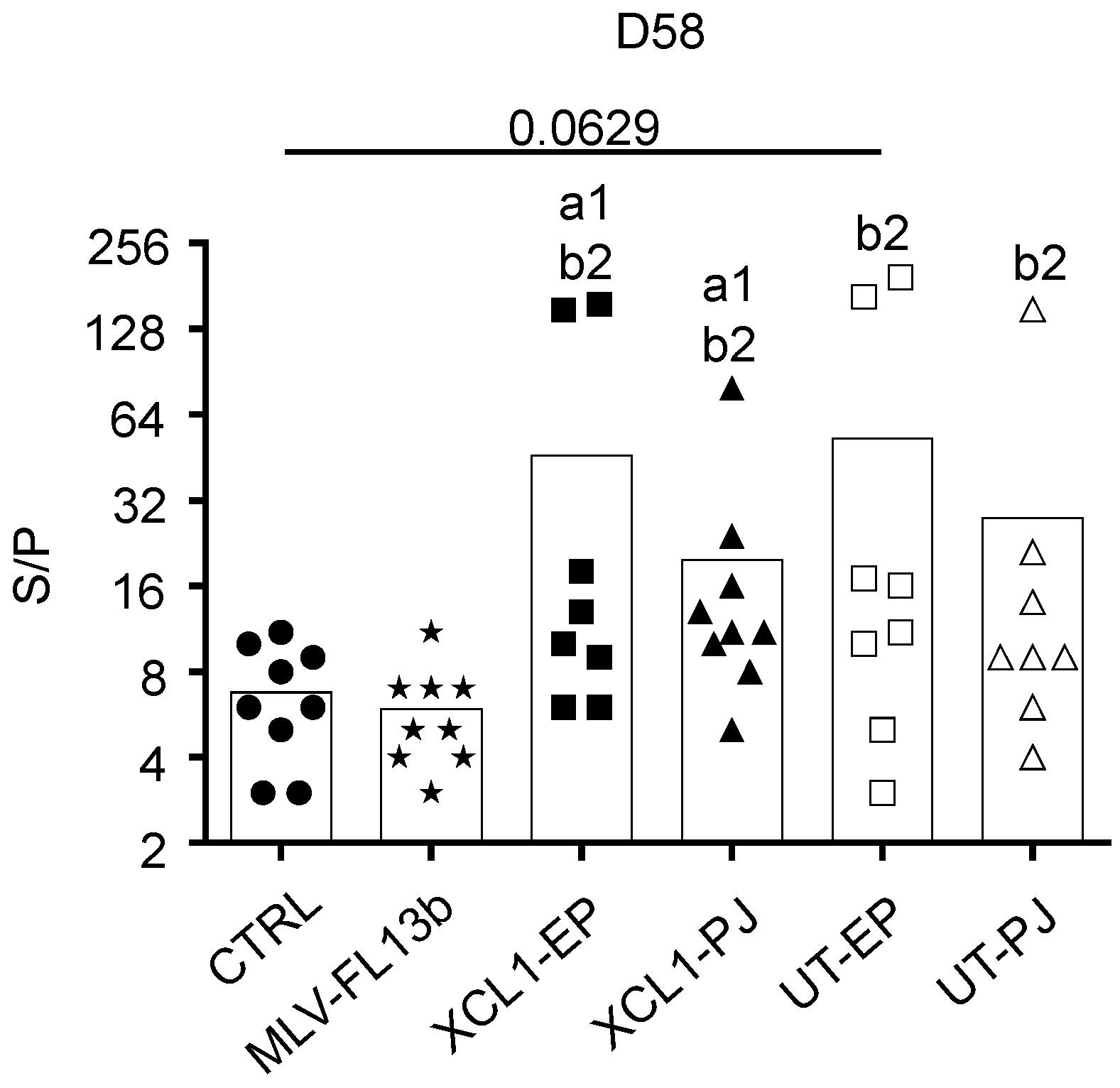{kind=link}
{kind=link}
{kind=link}
| Vaccines and Dates | Groups | |||||
|---|---|---|---|---|---|---|
| Ctrl 1 | MLV-FL13b | DNA+MLV-FL13b | ||||
| XCL1-EP | XCL1-PJ | UT-EP | UT-PJ | |||
| DNA D0 | pXCL1-N 2 pXCL1-GP4GP5M pXCL1-NSP1β1 pXCL1-NSP1β2 pRdRp | pXCL1-N pXCL1-GP4GP5M pXCL1-NSP1β1 pXCL1-NSP1β2 pRdRp | pN pGP4GP5M pNSP1β pRdRp | pN pGP4GP5M pNSP1β pRdRp | ||
| MLV-FL13b D34 | - 3 | + 4 | + | + | + | + |
| FL07 challenge D63 | + | + | + | + | + | + |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernelin-Cottet, C.; Urien, C.; Fretaud, M.; Langevin, C.; Trus, I.; Jouneau, L.; Blanc, F.; Leplat, J.-J.; Barc, C.; Boulesteix, O.; et al. A DNA Prime Immuno-Potentiates a Modified Live Vaccine against the Porcine Reproductive and Respiratory Syndrome Virus but Does Not Improve Heterologous Protection. Viruses 2019, 11, 576. https://doi.org/10.3390/v11060576
Bernelin-Cottet C, Urien C, Fretaud M, Langevin C, Trus I, Jouneau L, Blanc F, Leplat J-J, Barc C, Boulesteix O, et al. A DNA Prime Immuno-Potentiates a Modified Live Vaccine against the Porcine Reproductive and Respiratory Syndrome Virus but Does Not Improve Heterologous Protection. Viruses. 2019; 11(6):576. https://doi.org/10.3390/v11060576
Chicago/Turabian StyleBernelin-Cottet, Cindy, Céline Urien, Maxence Fretaud, Christelle Langevin, Ivan Trus, Luc Jouneau, Fany Blanc, Jean-Jacques Leplat, Céline Barc, Olivier Boulesteix, and et al. 2019. "A DNA Prime Immuno-Potentiates a Modified Live Vaccine against the Porcine Reproductive and Respiratory Syndrome Virus but Does Not Improve Heterologous Protection" Viruses 11, no. 6: 576. https://doi.org/10.3390/v11060576
APA StyleBernelin-Cottet, C., Urien, C., Fretaud, M., Langevin, C., Trus, I., Jouneau, L., Blanc, F., Leplat, J.-J., Barc, C., Boulesteix, O., Riou, M., Dysart, M., Mahé, S., Studsrub, E., Nauwynck, H., Bertho, N., Bourry, O., & Schwartz-Cornil, I. (2019). A DNA Prime Immuno-Potentiates a Modified Live Vaccine against the Porcine Reproductive and Respiratory Syndrome Virus but Does Not Improve Heterologous Protection. Viruses, 11(6), 576. https://doi.org/10.3390/v11060576