Usutu Virus Isolated from Rodents in Senegal

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Site
2.2. Arbovirus Screening and Isolation Attempt
2.3. RT-PCR and Sequencing of USUV Isolates
2.4. Phylogenetic Tree
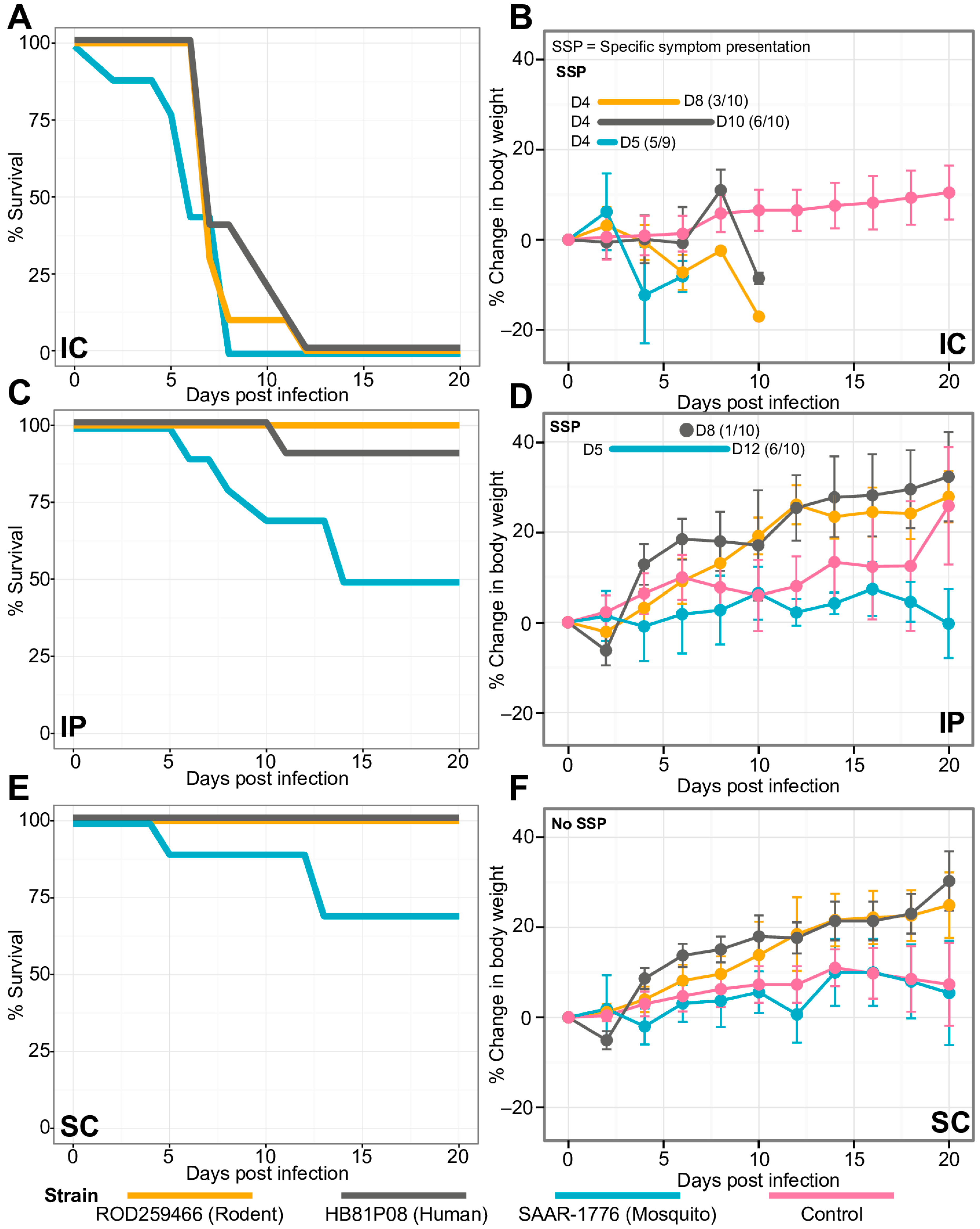
2.5. Virus Strains for In Vivo Experiments
2.6. Animal Experiments
3. Results
3.1. Usutu Virus Detection in Rodents and Shrew
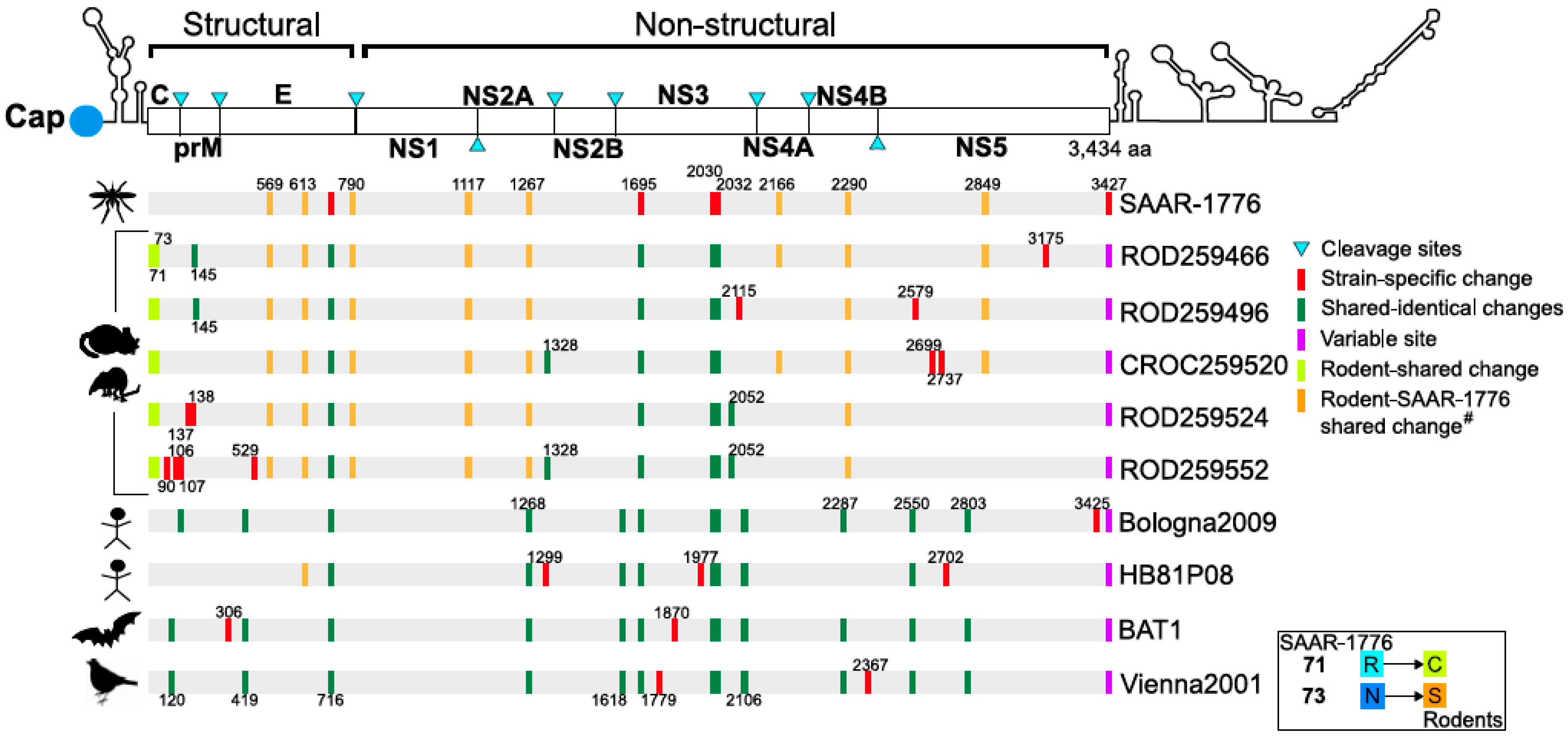
3.2. Map and Genomic Analysis
3.3. Phylogenetic Analysis
3.4. Comparative In Vivo Study of USUV Effect on Mouse Growth and Survival
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Woodall, J.P. The viruses isolated from arthropods at the East African Virus Research Institute in the 26 years ending December 1963. Proc. E Afr. Acad. 1964, 2, 141–146. [Google Scholar]
- Saiz, J.C.; Blazquez, A.B. Usutu virus: Current knowledge and future perspectives. Virus Adapt. Treat. 2017, 9, 27–40. [Google Scholar] [CrossRef]
- Odelola, H.A.; Fabiyi, A. Antigenic relationships among Nigerian strains of West Nile virus by complement fixation and agar gel precipitation techniques. Trans. R. Soc. Trop. Med. Hyg. 1976, 70, 138–144. [Google Scholar] [CrossRef]
- Cornet, M.; Robin, Y.; Chateau, R.; Heme, G.; Adam, C.; Valade, M.; Gonidec, G. Isolement d’arbovirus au Sénégal Oriental a partir de moustiques (1972–1977) et notes sur l’épidémiologie des virus transmis par les Aedes en particulier du virus amaril. Cahiers ORSTOM Série Entomologie Médicale et Parasitologie 1979, 17, 149–163. [Google Scholar]
- Weissenböck, H.; Kolodziejek, J.; Url, A.; Lussy, H.; Rebel-Bauder, B.; Nowotny, N. Emergence of Usutu virus, an African mosquito-borne flavivirus of the Japanese encephalitis virus group, central Europe. Emerg. Infect. Dis. 2002, 8, 652–656. [Google Scholar] [CrossRef] [PubMed]
- Weissenböck, H.; Bakonyi, T.; Rossi, G.; Mani, P.; Nowotny, N. Usutu virus, Italy, 1996. Emerg. Infect. Dis. 2013, 19, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Engel, D.; Jost, H.; Wink, M.; Borstler, J.; Bosch, S.; Garigliany, M.M.; Jöst, A.; Czajka, C.; Lühken, R.; Ziegler, U.; et al. Reconstruction of the evolutionary history and dispersal of Usutu virus, a neglected emerging arbovirus in Europe and Africa. MBio 2016, 7, e01938-15. [Google Scholar] [CrossRef]
- Ashraf, U.; Ye, J.; Ruan, X.; Wan, S.; Zhu, B.; Cao, S. Usutu Virus: An Emerging Flavivirus in Europe. Viruses 2015, 7, 219–238. [Google Scholar] [CrossRef] [Green Version]
- Kuno, G.; Chang, G.-J.; Tsuchiya, K.R.; Karabatsos, N.; Cropp, C.B. Phylogeny of the genus Flavivirus. J. Virol. 1998, 72, 73–83. [Google Scholar]
- Moureau, G.; Cook, S.; Lemey, P.; Nougairede, A.; Forrester, N.L.; Khasnatinov, M.; Charrel, R.N.; Firth, A.E.; Gould, E.A.; de Lamballerie, X. New insights into flavivirus evolution, taxonomy and biogeographic history, extended by analysis of canonical and alternative coding sequences. PLoS ONE 2015, 10, e0117849. [Google Scholar] [CrossRef]
- Cadar, D.; Becker, N.; de Mendonca Campos, R.; Börstler, J.; Jöst, H.; Schmidt-Chanasit, J. Usutu virus in bats, Germany, 2013. Emerg. Infect. Dis. 2014, 20, 1771–1773. [Google Scholar] [CrossRef] [PubMed]
- Pecorari, M.; Longo, G.; Gennari, W.; Grottola, A.; Sabbatini, A.; Tagliazucchi, S.; Savini, G.; Monaco, F.; Simone, M.; Lelli, R.; et al. First human case of Usutu virus neuroinvasive infection, Italy, August-September 2009. Eurosurveillance 2009, 14, 19446. [Google Scholar] [PubMed]
- Cavrini, F.; DellaPepa, M.E.; Gaibani, P.; Pierro, A.M.; Rossini, G.; Landini, M.P.; Sambri, V. A rapid and specific real-time RT-PCR assay to identify Usutu virus in human plasma, serum, and cerebrospinal fluid. J. Clin. Virol. 2011, 50, 221–223. [Google Scholar] [CrossRef] [PubMed]
- Santini, M.; Vilibic-Cavlek, T.; Barsic, B.; Barbic, L.; Savic, V.; Stevanovic, V.; Listes, E.; Di Gennaro, A.; Savini, G. First cases of human Usutu virus neuroinvasive infection in Croatia, August–September 2013, clinical and laboratory features. J. Neurovirol. 2015, 21, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Allering, L.; Jöst, H.; Emmerich, P.; Günther, S.; Lattwein, E.; Schmidt, M.; Seifried, E.; Sambri, V.; Hourfar, K.; Schmidt-Chanasit, J. Detection of Usutu virus infection in a healthy blood donor from south-west Germany, 2012. Eurosurveillance 2012, 17, 20341. [Google Scholar] [PubMed]
- Gaibani, P.; Pierro, A.; Alicino, R.; Rossini, G.; Cavrini, F.; Landini, M.P.; Sambri, V. Detection of Usutu-virus-specific IgG in blood donors from northern Italy. Vector Borne Zoonotic Dis. 2012, 12, 431–433. [Google Scholar] [CrossRef] [PubMed]
- Vilibic-Cavlek, T.; Kaic, B.; Barbic, L.; Pem-Novosel, I.; Slavic-Vrzic, V.; Lesnikar, V.; Kurecic-Filipovic, S.; Babic-Erceg, A.; Listes, E.; Stevanovic, V.; et al. First evidence of simultaneous occurrence of West Nile virus and Usutu virus neuroinvasive disease in humans in Croatia during the 2013 outbreak. Infection 2014, 42, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Achazi, K.; Růžek, D.; Donoso-Mantke, O.; Schlegel, M.; Ali, H.S.; Wenk, M.; Schmidt-Chanasit, J.; Ohlmeyer, L.; Rühe, F.; Vor, T.; et al. Rodents as Sentinels for the Prevalence of Tick-Borne Encephalitis Virus. Vector Borne Zoonotic Dis. 2011, 11, 641–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cigarroa-Toledo, N.; Talavera-Aguilar, L.G.; Baak-Baak, C.M.; García-Rejón, J.E.; Hernandez-Betancourt, S.; Blitvich, B.J.; Machain-Williams, C. Serologic Evidence of Flavivirus Infections in Peridomestic Rodents in Merida, Mexico. J. Wildl. Dis. 2016, 52, 168–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatmi, S.S.; Zehra, R.; Carpenter, D.O. Powassan Virus—A New Reemerging Tick-Borne Disease. Front. Public Health 2017, 5, 342. [Google Scholar] [CrossRef] [PubMed]
- Mlera, L.; Bloom, M.E. The Role of Mammalian Reservoir Hosts in Tick-Borne Flavivirus Biology. Front. Cell. Infect. Microbiol. 2018, 8, 298. [Google Scholar] [CrossRef]
- Meerburg, B.G.; Singleton, G.R.; Kijlstra, A. Rodent-borne diseases and their risks for public health. Crit. Rev. Microbiol. 2009, 35, 22170. [Google Scholar] [CrossRef]
- Coz, J.; Valade, M.; Cornet, M.; Robin, Y. Transmission transovarienne d’un Flavivirus le virus Koutango chez Aedes aegypti L. C. R. Acad. Sci. Paris 1976, 283, 109–110. [Google Scholar]
- Diagne, M.M.; Faye, M.; Faye, O.; Sow, A.; Balique, F.; Sembene, M.; Granjon, L.; Handschumacher, P.; Faye, O.; Diallo, M.; et al. Emergence of Wesselsbron virus among black rat and humans in Eastern Senegal in 2013. One Health 2017, 3, 23–28. [Google Scholar] [CrossRef]
- Dalecky, A.; Ba, K.; Piry, S.; Lippens, C.; Diagne, C.A.; Kane, M.; Sow, A.; Diallo, M.; Niang, Y.; Konecny, A.; et al. Range expansion of the invasive house mouse Mus musculus domesticus in Senegal, West Africa: A synthesis of trapping data over three decades, 1983–2014. Mammal Rev. 2015, 45, 176–190. [Google Scholar] [CrossRef]
- Granjon, L.; Duplantier, J.M. Les Rongeurs de l’Afrique Sahélo-Soudanienne; (Faune et Flore tropicales; 43); IRD/MNHN: Paris, France, 2010; p. 215. ISBN 978-2-7099-1675-2. [Google Scholar]
- Happold, M.; Happold, D.C.D. Mammals of Africa, Vol. IV, Hedgehogs, Shrews and Bats; Bloomsbury Publishing: London, UK, 2013; p. 789. ISBN 978-1-4081-2253-2. [Google Scholar]
- Granjon, L.; Dobigny, G. The importance of cytotaxonomy in understanding the biogeography of African rodents: Lake Chad murids as an example. Mammal Rev. 2003, 33, 77–91. [Google Scholar] [CrossRef]
- Lecompte, E.; Brouat, C.; Duplantier, J.M.; Galan, M.; Granjon, L.; Loiseau, A.; Mouline, K.; Cosson, J.F. Molecular identification of four cryptic species of Mastomys (Rodentia, Murinae). Biochem. Syst. Ecol. 2005, 33, 681–689. [Google Scholar] [CrossRef]
- Dobigny, G.; Tatard, C.; Kane, M.; Gauthier, P.; Brouat, C.; Bâ, K.; Duplantier, J.-M. A cytotaxonomic and DNA-based survey of rodents from Northern Cameroon and WesternChad. Mamm. Biol. 2011, 76, 417–427. [Google Scholar] [CrossRef]
- Mills, J.N.; Childs, J.E.; Ksiazek, T.G.; Peters, C.J.; Velleca, W.M. Methods for Trapping and Sampling Small Mammals for Virologic Testing; U.S. Department of Health & Human Services, CDC: Atlanta, GA, USA, 1995.
- Sikes, R.S.; Gannon, W.L.; The Animal Care and Use Committee of the American Society of Mammalogists. Guidelines of the American Society of Mammalogists for the use of wild mammals in research. J. Mammal. 2011, 92, 235–253. [Google Scholar] [CrossRef] [Green Version]
- Nikolay, B.; Dupressoir, A.; Firth, C.; Faye, O.; Boye, C.S.; Diallo, M.; Sall, A.A. Comparative full length genome sequence analysis of usutu virus isolates from Africa. Virol. J. 2013, 10, 217. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Rambaut, A. Se-Al: Sequence Alignment Editor. 1996. Available online: http://evolve.zoo.ox.ac.uk/ (accessed on 2 March 2018).
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4, Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, 1–5. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Fall, G.; Di Paola, N.; Faye, M.; Dia, M.; de Melo Freire, C.C.; Loucoubar, C.; de Andrade Zanotto, P.M.; Faye, O.; Sall, A.A. Biological and phylogenetic characteristics of West African lineages of West Nile virus. PLoS Negl. Trop. Dis. 2017, 11, e0006078. [Google Scholar] [CrossRef]
- Nikolay, B.; Weidmann, M.; Dupressoir, A.; Faye, O.; Boye, C.S.; Diallo, M.; Sall, A.A. Development of a Usutu virus specific real-time reverse transcription PCR assay based on sequenced strains from Africa and Europe. J. Virol. Methods 2014, 197, 51–54. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, U.; Fast, C.; Eiden, M.; Bock, S.; Schulze, C.; Hoeper, D.; Ochs, A.; Schlieben, P.; Keller, M.; Zielke, D.E.; et al. Evidence for an independent third Usutu virus introduction into Germany. Vet. Microbiol. 2016, 192, 60–66. [Google Scholar] [CrossRef]
- Cadar, D.; Lühken, R.; van der Jeugd, H.; Garigliany, M.; Ziegler, U.; Keller, M.; Lahoreau, J.; Lachmann, L.; Becker, N.; Kik, M.; et al. Widespread activity of multiple lineages of Usutu virus, western Europe, 2016. Eurosurveillance 2017, 22, 30452. [Google Scholar] [CrossRef]
- Figuerola, J.; Baouab, R.E.; Soriguer, R.; Fassi-Fihri, O.; Llorente, F.; Jimenez-Clavero, M.A. West Nile virus antibodies in wild birds, Morocco, 2008. Emerg. Infect. Dis. 2009, 15, 1651–1653. [Google Scholar] [CrossRef]
- Ayadi, T.; Hammouda, A.; Poux, A.; Boulinier, T.; Lecollinet, S.; Selmi, S. Evidence of exposure of laughing doves (Spilopelia senegalensis) to West Nile and Usutu viruses in southern Tunisian oases. Epidemiol. Infect. 2017, 13, 2808–2816. [Google Scholar] [CrossRef]
- Bakonyi, T.; Erdélyi, K.; Brunthaler, R.; Dán, Á.; Weissenböck, H.; Nowotny, N. Usutu virus, Austria and Hungary, 2010–2016. Emerg. Microbes Infect. 2017, 6, e85. [Google Scholar] [CrossRef]
- Michel, F.; Fischer, D.; Eiden, M.; Fast, C.; Reuschel, M.; Müller, K.; Rinder, M.; Urbaniak, S.; Brandes, F.; Schwehn, R.; et al. West Nile Virus and Usutu Virus Monitoring of Wild Birds in Germany. Int. J. Environ. Res. Public Health 2018, 15, 171. [Google Scholar] [CrossRef]
- Ben Hassine, T.; De Massis, F.; Calistri, P.; Savini, G.; BelHaj Mohamed, B.; Ranen, A.; Di Gennaro, A.; Sghaier, S.; Hammami, S. First detection of cocirculation of West Nile and Usutu viruses in equids in the south-west of Tunisia. Transbound. Emerg. Dis. 2014, 61, 385–389. [Google Scholar] [CrossRef]
- Vanhomwegen, J.; Beck, C.; Despres, P.; Figuerola, A.; García, R.; Lecollinet, S.; López-Roig, M.; Manuguerra, J.C.; Serra-Cobo, J. Circulation of Zoonotic Arboviruses in Equine Populations of Mallorca Island (Spain). Vector Borne Zoonotic Dis. 2017, 17, 340–346. [Google Scholar] [CrossRef]
- Durand, B.; Haskouri, H.; Lowenski, S.; Vachiery, N.; Beck, C.; Lecollinet, S. Seroprevalence of West Nile and Usutu viruses in military working horses and dogs, Morocco, 2012, dog as an alternative WNV sentinel species? Epidemiol. Infect. 2016, 144, 1857–1864. [Google Scholar] [CrossRef]
- García-Bocanegra, I.; Paniagua, J.; Gutiérrez-Guzmán, A.V.; Lecollinet, S.; Boadella, M.; Arenas-Montes, A.; Cano-Terriza, D.; Lowenski, S.; Gortázar, C.; Höfle, U. Spatio-temporal trends and risk factors affecting West Nile virus and related flavivirus exposure in Spanish wild ruminants. BMC Vet. Res. 2016, 12, 249. [Google Scholar] [CrossRef]
- Nikolay, B.; Diallo, M.; Boye, C.S.; Sall, A.A. Usutu virus in Africa. Vector Borne Zoonotic Dis. 2011, 11, 1417–1423. [Google Scholar] [CrossRef]
- Saiz, J.C.; Vazquez-Calvo, A.; Blazquez, A.B.; Merino-Ramos, T.; Escribano-Romero, E.; Martin-Acebes, M.A. Zika Virus: The Latest Newcomer. Front Microbiol. 2016, 7, 496. [Google Scholar] [CrossRef]
- Urbanowski, M.D.; Hobman, T.C. The West Nile virus capsid protein blocks apoptosis through a phosphatidylinositol 3-kinase-dependent mechanism. J. Virol. 2012, 87, 872–881. [Google Scholar] [CrossRef]
- Okamoto, T.; Suzuki, T.; Kusakabe, S.; Tokunaga, M.; Hirano, J.; Miyata, Y.; Matsuura, Y. Regulation of Apoptosis during Flavivirus Infection. Viruses 2017, 9, 243. [Google Scholar] [CrossRef]
- Woolhouse, M.E.; Taylor, L.H.; Haydon, D.T. Population biology of multihost pathogens. Science 2001, 292, 1109–1112. [Google Scholar] [CrossRef] [PubMed]
- Freire, J.M.; Santos, N.C.; Veiga, A.S.; Da Poian, A.T.; Castanho, M.A. Rethinking the capsid proteins of enveloped viruses:multifunctionality from genome packagingto genome transfection. FEBS J. 2015, 282, 2267–2278. [Google Scholar] [CrossRef] [PubMed]
- Blázquez, A.B.; Escribano-Romero, E.; Martín-Acebes, M.A.; Petrovic, T.; Saiz, J.C. Limited susceptibility of mice to Usutu virus (USUV) infection and induction of flavivirus cross-protective immunity. Virology 2015, 482, 67–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dow, C.; Jarrett, W.F. Age, strain and sex differences in susceptibility to Cysticercus fasciolaris in the mouse. Exp. Parasitol. 1960, 10, 72–74. [Google Scholar] [CrossRef]
- Coffey, L.L.; Forrester, N.; Tsetsarkin, K.; Vasilakis, N.; Weaver, S.C. Factors shaping the adaptive landscape for arboviruses: Implications for the emergence of disease. Future Microbiol. 2013, 8, 155–176. [Google Scholar] [CrossRef] [PubMed]
- Gaye, A.; IPD Medical Entomology Team. Personal communication, 2017.
- Nikolay, B.; Diallo, M.; Faye, O.; Boye, C.S.; Sall, A.A. Vector Competence of Culex neavei (Diptera: Culicidae) for Usutu Virus. Am. J. Trop. Med. Hyg. 2012, 86, 993–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fros, J.J.; Miesen, P.; Vogels, C.B.; Gaibani, P.; Sambri, V.; Martina, B.E.; Koenraadt, C.J.; van Rij, R.P.; Vlak, J.M.; Takken, W.; et al. Comparative Usutu and West Nile virus transmission potential by local Culex pipiens mosquitoes in north-western Europe. One Health 2015, 1, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Molaei, G.; Andreadis, T.G.; Armstrong, P.M.; Bueno, R., Jr.; Dennett, J.A.; Real, S.V.; Sargent, C.; Bala, A.; Randle, Y.; Guzman, H.; et al. Host feeding pattern of Culex quinquefasciatus (Diptera: Culicidae) and its role in transmission of West Nile virus in Harris County, Texas. Am. J. Trop. Med. Hyg. 2007, 77, 73–81. [Google Scholar] [CrossRef]
- Savage, H.M.; Kothera, L. The Culex pipiens complex in the Mississippi River basin: Identification, distribution, and bloodmeal hosts. J. Am. Mosq. Control Assoc. 2012, 28, 93–99. [Google Scholar] [CrossRef]
- Janssen, N.; Fernandez-Salas, I.; Díaz González, E.E.; Gaytan-Burns, A.; Medina-de la Garza, C.E.; Sanchez-Casas, R.M.; Börstler, J.; Cadar, D.; Schmidt-Chanasit, J.; Jöst, H. Mammalophilic feeding behavior of Culex quinquefasciatus mosquitoes collected in the cities of Chetumal and Cancun, Yucatán Peninsula, Mexico. Trop. Med. Int. Health 2015, 20, 1488–1491. [Google Scholar] [CrossRef] [Green Version]
- Bonwitt, J.; Sáez, A.M.; Lamin, J.; Ansumana, R.; Dawson, M.; Buanie, J.; Lamin, J.; Sondufu, D.; Borchert, M.; Sahr, F.; et al. At Home with Mastomys and Rattus: Human-Rodent Interactions and Potential for Primary Transmission of Lassa Virus in Domestic Spaces. Am. J. Trop. Med. Hyg. 2017, 96, 935–943. [Google Scholar] [CrossRef]
- Mitchell-Jones, A.J.; Amori, G.; Bogdanowicz, W.; Kryštufek, B.; Reijnders, P.J.H.; Spitzenberger, E.; Stubbe, M.; Thissen, J.B.M.; Vohralík, V.; Zima, J. Atlas of European Mammals; Academic Press: London, UK, 1999. [Google Scholar]
- Kryštufek, B.; Griffiths, H.I. Species richness and rarity in European rodents. Ecography 2002, 25, 120–128. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GenBank Accession Number | Locality | Latitude Coordinates | Longitude Coordinates | ID Number | Host Species |
|---|---|---|---|---|---|
| MH727238 | Kedougou | 12.553997 | −12.179781 | ROD259466 | Mastomys natalensis |
| MH727239 | Kedougou | 12.553614 | −12.179837 | ROD259496 | Mastomys natalensis |
| MH727240 | Kedougou | 12.555794 | −12.177485 | CROC259520 | Shrew Crocidura sp. |
| MH727241 | Kedougou | 12.555835 | −12.178598 | ROD259524 | Mastomys natalensis |
| MH727242 | Kedougou | 12.55654 | −12.177977 | ROD259552 | Rattus rattus |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diagne, M.M.; Ndione, M.H.D.; Di Paola, N.; Fall, G.; Bedekelabou, A.P.; Sembène, P.M.; Faye, O.; Zanotto, P.M.d.A.; Sall, A.A. Usutu Virus Isolated from Rodents in Senegal. Viruses 2019, 11, 181. https://doi.org/10.3390/v11020181
Diagne MM, Ndione MHD, Di Paola N, Fall G, Bedekelabou AP, Sembène PM, Faye O, Zanotto PMdA, Sall AA. Usutu Virus Isolated from Rodents in Senegal. Viruses. 2019; 11(2):181. https://doi.org/10.3390/v11020181
Chicago/Turabian StyleDiagne, Moussa Moïse, Marie Henriette Dior Ndione, Nicholas Di Paola, Gamou Fall, André Pouwedeou Bedekelabou, Pape Mbacké Sembène, Ousmane Faye, Paolo Marinho de Andrade Zanotto, and Amadou Alpha Sall. 2019. "Usutu Virus Isolated from Rodents in Senegal" Viruses 11, no. 2: 181. https://doi.org/10.3390/v11020181