Heterologous Replicase from Cucumoviruses can Replicate Viral RNAs, but is Defective in Transcribing Subgenomic RNA4A or Facilitating Viral Movement

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants and Viruses
2.2. Plasmid Construction
2.3. Agrobacterium-Mediated Plant Inoculation
2.4. Viral Replication Assays
2.5. RNA Gel Blot Analysis
2.6. Immunoblotting Analysis
3. Results
3.1. Effect of Genomic Reassortment on Virus Infection
3.2. Heterologous Combination of CMV RNA1 and TAV RNA2 was Defective in Viral Long-Distance Movement
3.3. Heterologous Replicase Components were Compatible in Replication of GFP-Expressing RNA3
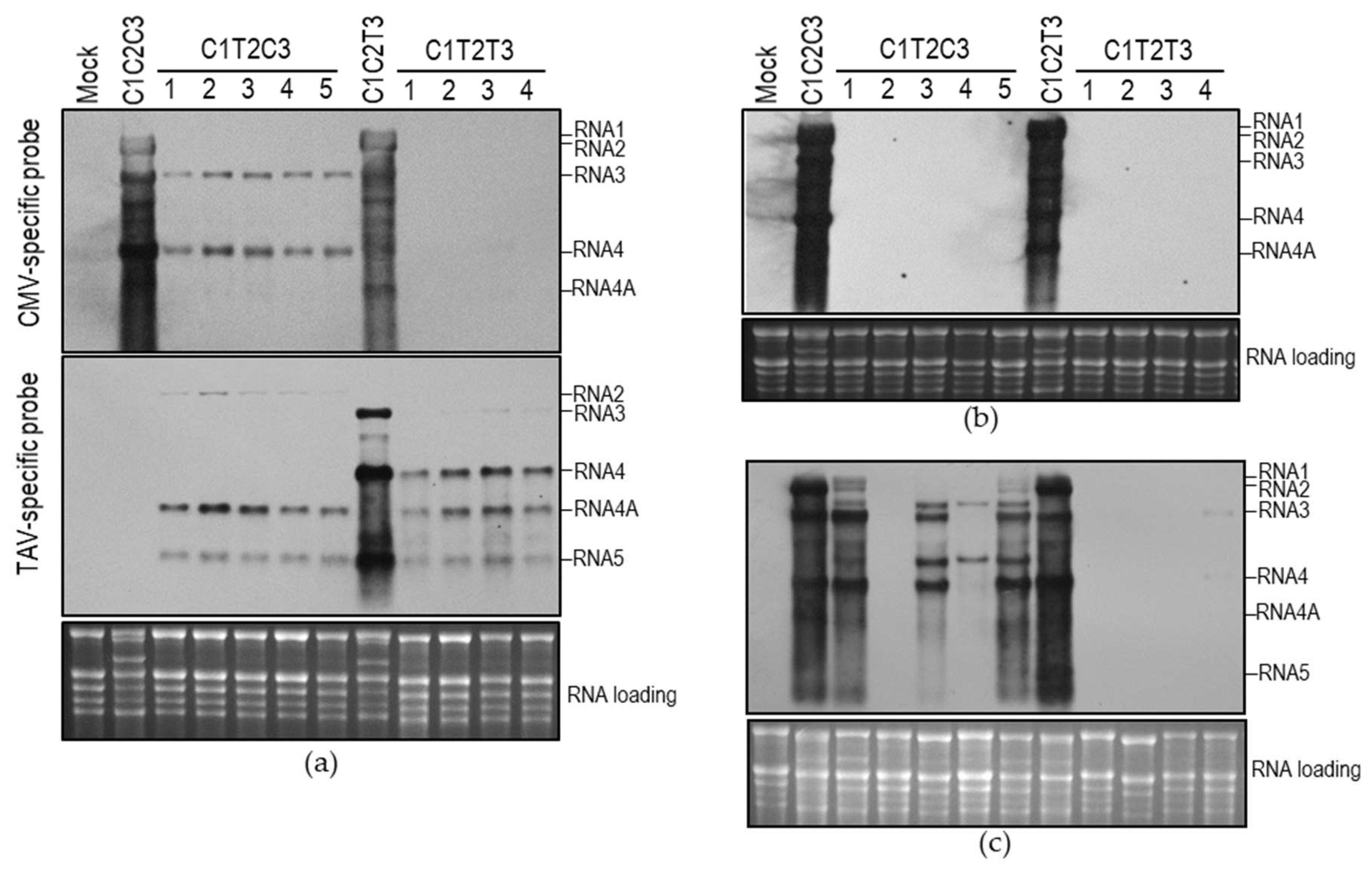
3.4. Heterologous Combination between TAV 1a and CMV 2a was Deficient in Synthesis of CMV Subgenomic RNAs
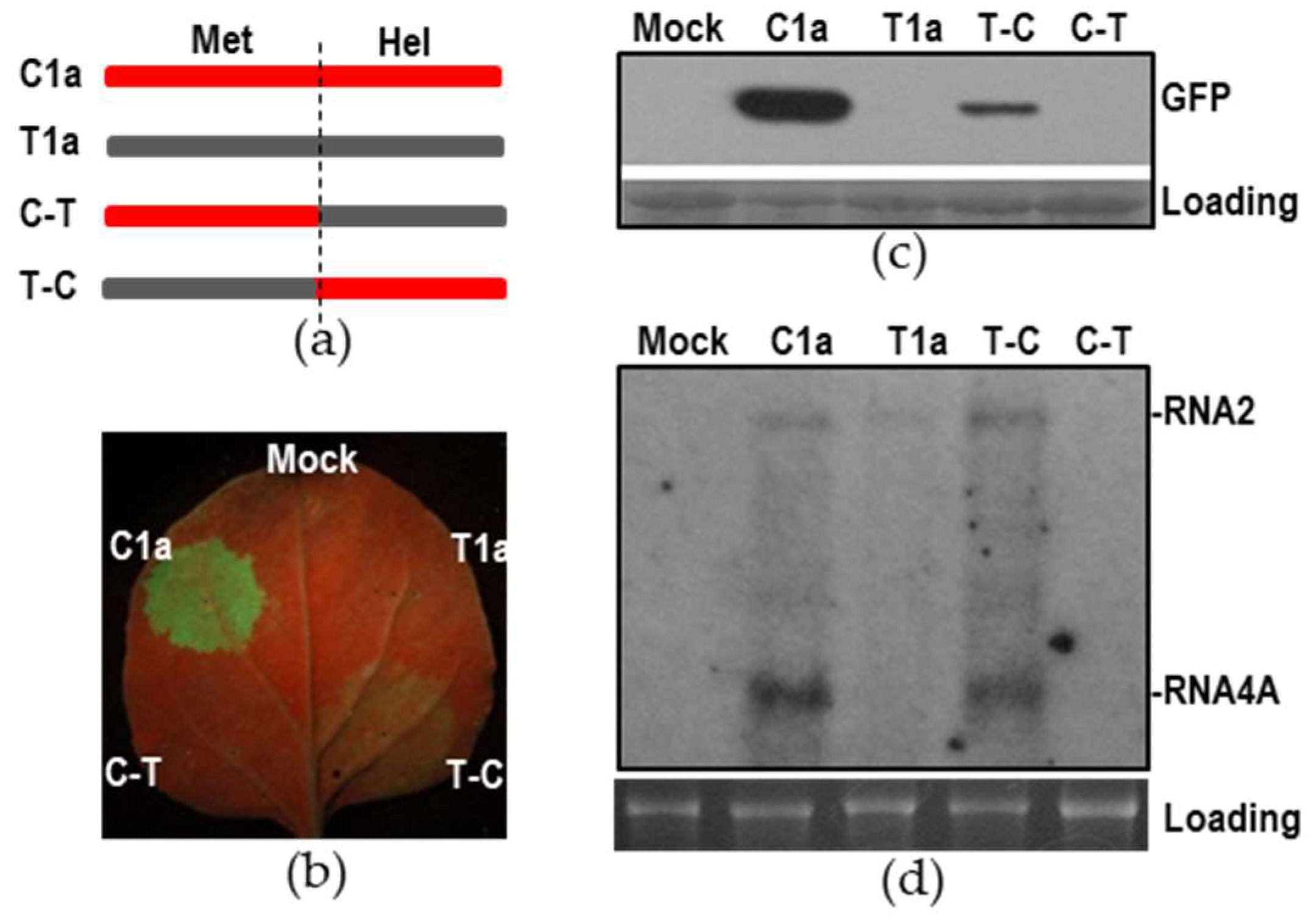
3.5. The Helicase Domain of 1a was Required for Synthesis of CMV RNA4A
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Palukaitis, P.; García-Arenal, F. Cucumoviruses. Adv. Virus Res. 2003, 62, 241–323. [Google Scholar] [CrossRef] [PubMed]
- Ahlquist, P.; Dasgupta, R.; Kaesberg, P. Near identity of 3- RNA secondary structure in bromoviruses and cucumber mosaic virus. Cell 1981, 23, 183–189. [Google Scholar] [CrossRef]
- Symons, R.H. Cucumber mosaic virus RNA contains 7-methyl guanosine at the 5′-terminus of all four RNA species. Mol. Biol. Rep. 1975, 2, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Jacquemond, M. Cucumber mosaic virus. Adv. Virus Res. 2012, 84, 439–504. [Google Scholar] [CrossRef] [PubMed]
- Ahlquist, P.; Noueiry, A.O.; Lee, W.M.; Kushner, D.B.; Dye, B.T. Host factors in positive-strand RNA virus genome replication. J. Virol. 2003, 77, 8181–8186. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.C. Lessons learned from the core RNA promoters of Brome mosaic virus and Cucumber mosaic virus. Mol. Plant Pathol. 2002, 3, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Palukaitis, P.; Park, Y.I. Phosphorylation of cucumber mosaic virus RNA polymerase 2a protein inhibits formation of replicase complex. EMBO J. 2002, 21, 2292–2300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Reilly, E.K.; Tang, N.; Ahlquist, P.; Kao, C.C. Biochemical and genetic analyses of the interaction between the helicase-like and polymerase-like proteins of the brome mosaic virus. Virology 1995, 214, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.W.; Anderson, B.J.; Haase, H.R.; Symons, R.H. New overlapping gene encoded by the cucumber mosaic virus genome. Virology 1994, 198, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Shimura, H.; Masuta, C.; Yoshida, N.; Sueda, K.; Suzuki, M. The 2b protein of Asparagus virus 2 functions as an RNA silencing suppressor against systemic silencing to prove functional synteny with related cucumoviruses. Virology 2013, 442, 180–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, H.W.; Ji, L.H.; Scott, S.W.; Symons, R.H.; Ding, S.W. Ilarviruses encode a Cucumovirus-like 2b gene that is absent in other genera within the Bromoviridae. J. Virol. 1998, 72, 6956–6959. [Google Scholar] [PubMed]
- Diaz-Pendon, J.A.; Li, F.; Li, W.X.; Ding, S.W. Suppression of antiviral silencing by cucumber mosaic virus 2b protein in Arabidopsis is associated with drastically reduced accumulation of three classes of viral small interfering RNAs. Plant Cell 2007, 19, 2053–2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, C.G.; Fang, Y.Y.; Zhou, B.J.; Zhao, J.H.; Hou, W.N.; Zhu, H.; Ding, S.W.; Guo, H.S. Suppression of Arabidopsis ARGONAUTE1-mediated slicing, transgene-induced RNA silencing, and DNA methylation by distinct domains of the Cucumber mosaic virus 2b protein. Plant Cell 2012, 24, 259–274. [Google Scholar] [CrossRef] [PubMed]
- Goto, K.; Kobori, T.; Kosaka, Y.; Natsuaki, T.; Masuta, C. Characterization of silencing suppressor 2b of cucumber mosaic virus based on examination of its small RNA-binding abilities. Plant Cell Physiol. 2007, 48, 1050–1060. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.Y.; Chen, F.F.; Liao, Q.S.; Zhang, H.R.; Chen, Y.F.; Chen, J.S. 2b ORFs encoded by subgroup IB strains of cucumber mosaic virus induce differential virulence on Nicotiana species. J. Gen. Virol. 2007, 88, 2596–2604. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, I.; Martinez, L.; Rakitina, D.V.; Lewsey, M.G.; Atencio, F.A.; Llave, C.; Kalinina, N.O.; Carr, J.P.; Palukaitis, P.; Canto, T. Cucumber mosaic virus 2b protein subcellular targets and interactions: Their significance to RNA silencing suppressor activity. Mol. Plant Microbe Interact. 2010, 23, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, I.; Rakitina, D.; Semashko, M.; Taliansky, M.; Praveen, S.; Palukaitis, P.; Carr, J.P.; Kalinina, N.; Canto, T. RNA binding is more critical to the suppression of silencing function of Cucumber mosaic virus 2b protein than nuclear localization. RNA 2012, 18, 771–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cillo, F.; Mascia, T.; Pasciuto, M.M.; Gallitelli, D. Differential effects of mild and severe Cucumber mosaic virus strains in the perturbation of MicroRNA-regulated gene expression in tomato map to the 3′ sequence of RNA 2. Mol. Plant Microbe Interact. 2009, 22, 1239–1249. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Chen, A.; Chen, W.; Westwood, J.H.; Baulcombe, D.C.; Carr, J.P. Using a Viral Vector to Reveal the Role of MicroRNA159 in Disease Symptom Induction by a Severe Strain of Cucumber mosaic virus. Plant Physiol. 2014, 164, 1378–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewsey, M.; Robertson, F.C.; Canto, T.; Palukaitis, P.; Carr, J.P. Selective targeting of miRNA-regulated plant development by a viral counter-silencing protein. Plant J. 2007, 50, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yuan, Y.R.; Pei, Y.; Lin, S.S.; Tuschl, T.; Patel, D.J.; Chua, N.H. Cucumber mosaic virus-encoded 2b suppressor inhibits Arabidopsis Argonaute1 cleavage activity to counter plant defense. Genes Dev. 2006, 20, 3255–3268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allison, R.F.; Janda, M.; Ahlquist, P. Infectious in vitro transcripts from cowpea chlorotic mottle virus cDNA clones and exchange of individual RNA components with brome mosaic virus. J. Virol. 1988, 62, 3581–3588. [Google Scholar] [PubMed]
- Rao, A.L.; Francki, R.I. Comparative studies on tomato aspermy and cucumber mosaic viruses. VI. Partial compatibility of genome segments from the two viruses. Virology 1981, 114, 573–575. [Google Scholar] [CrossRef]
- Suzuki, M.; Yoshida, M.; Yoshinuma, T.; Hibi, T. Interaction of replicase components between Cucumber mosaic virus and Peanut stunt virus. J. Gen. Virol. 2003, 84, 1931–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinant, S.; Janda, M.; Kroner, P.A.; Ahlquist, P. Bromovirus RNA replication and transcription require compatibility between the polymerase- and helicase-like viral RNA synthesis proteins. J. Virol. 1993, 67, 7181–7189. [Google Scholar] [PubMed]
- Salanki, K.; Carrere, I.; Jacquemond, M.; Balazs, E.; Tepfer, M. Biological properties of pseudorecombinant and recombinant strains created with cucumber mosaic virus and tomato aspermy virus. J. Virol. 1997, 71, 3597–3602. [Google Scholar] [PubMed]
- Masuta, C.; Ueda, S.; Suzuki, M.; Uyeda, I. Evolution of a quadripartite hybrid virus by interspecific exchange and recombination between replicase components of two related tripartite RNA viruses. Proc. Natl. Acad. Sci. USA 1998, 95, 10487–10492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, J.; Wang, K.; Liu, X.; Chen, S.; Chen, J. The quantification of tomato microRNAs response to viral infection by stem-loop real-time RT-PCR. Gene 2009, 437, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Jin, S.J.; Zhang, H.; Wang, T.W.; Chen, J.; Liao, Q. Sequences and Infectious Clones of Tomato aspermy virus Strain Isolated from Beijing. J. Agric. Biotechnol. 2011, 19, 1120–1126. (In Chinese) [Google Scholar]
- Liao, Q.; Tu, Y.; Carr, J.P.; Du, Z. An improved cucumber mosaic virus-based vector for efficient decoying of plant microRNAs. Sci. Rep. 2015, 5, 13178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, T.M.; Palukaitis, P. Construction of full-length cDNA clones of cucumber mosaic virus RNAs 1, 2 and 3: Generation of infectious RNA transcripts. Mol. Gen. Genet. 1990, 222, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Weigel, D.; Glazebrook, J. Transformation of agrobacterium using the freeze-thaw method. CSH Protoc. 2006, 7. [Google Scholar] [CrossRef] [PubMed]
- Silhavy, D.; Molnar, A.; Lucioli, A.; Szittya, G.; Hornyik, C.; Tavazza, M.; Burgyan, J. A viral protein suppresses RNA silencing and binds silencing-generated, 21- to 25-nucleotide double-stranded RNAs. EMBO J. 2002, 21, 3070–3080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGarvey, P.; Tousignant, M.; Geletka, L.; Cellini, F.; Kaper, J.M. The complete sequence of a cucumber mosaic virus from Ixora that is deficient in the replication of satellite RNAs. J. Gen. Virol. 1995, 76, 2257–2270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Z.; Chen, A.; Chen, W.; Liao, Q.; Zhang, H.; Bao, Y.; Roossinck, M.J.; Carr, J.P. Nuclear-cytoplasmic partitioning of the Cucumber mosaic virus 2b protein determines the balance between its roles as a virulence determinant and RNA silencing suppressor. J. Virol. 2014, 88, 5228–5241. [Google Scholar] [CrossRef] [PubMed]
- Naylor, M.; Murphy, A.; Berry, J.; Carr, J. Salicylic acid can induce resistance to plant virus movement. Mol. Plant Microbe Interact. 1998, 11, 860–868. [Google Scholar] [CrossRef]
- Shi, B.J.; Ding, S.W.; Symons, R.H. In vivo expression of an overlapping gene encoded by the cucumoviruses. J. Gen. Virol. 1997, 78, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Soards, A.J.; Murphy, A.M.; Palukaitis, P.; Carr, J.P. Virulence and differential local and systemic spread of cucumber mosaic virus in tobacco are affected by the CMV 2b protein. Mol. Plant Microbe Interact. 2002, 15, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Kang, M.-Y.; Lee, J.-H.; Kang, W.-H.; Hwang, J.; Kwon, J.-K.; Kang, B.-C. Isolation and Characterization of Pepper Genes Interacting with the CMV-P1 Helicase Domain. PLoS ONE 2016, 11, E0146320. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.-H.; Seo, J.-K.; Chung, B.N.; Kim, K.-H.; Kang, B.-C. Helicase Domain Encoded by Cucumber mosaic virus RNA1 Determines Systemic Infection of Cmr1 in Pepper. PLoS ONE 2012, 7, E43136. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.; Lucas, W.J.; Ding, B.; Zaitlin, M. Viral RNA trafficking is inhibited in replicase-mediated resistant transgenic tobacco plants. Proc. Natl. Acad. Sci. USA 1996, 93, 12643–12647. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.K.; Kwon, S.J.; Choi, H.S.; Kim, K.H. Evidence for alternate states of Cucumber mosaic virus replicase assembly in positive- and negative-strand RNA synthesis. Virology 2009, 383, 248–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivakumaran, K.; Chen, M.H.; Roossinck, M.J.; Kao, C.C. Core promoter for initiation of Cucumber mosaic virus subgenomic RNA4A. Mol. Plant Pathol. 2002, 3, 43–52. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, S.; Lu, J.; Cheng, X.; Gu, Z.; Liao, Q.; Du, Z. Heterologous Replicase from Cucumoviruses can Replicate Viral RNAs, but is Defective in Transcribing Subgenomic RNA4A or Facilitating Viral Movement. Viruses 2018, 10, 590. https://doi.org/10.3390/v10110590
Gao S, Lu J, Cheng X, Gu Z, Liao Q, Du Z. Heterologous Replicase from Cucumoviruses can Replicate Viral RNAs, but is Defective in Transcribing Subgenomic RNA4A or Facilitating Viral Movement. Viruses. 2018; 10(11):590. https://doi.org/10.3390/v10110590
Chicago/Turabian StyleGao, Shuangyu, Jinda Lu, Xiaodong Cheng, Zhouhang Gu, Qiansheng Liao, and Zhiyou Du. 2018. "Heterologous Replicase from Cucumoviruses can Replicate Viral RNAs, but is Defective in Transcribing Subgenomic RNA4A or Facilitating Viral Movement" Viruses 10, no. 11: 590. https://doi.org/10.3390/v10110590