Novel Mitoviruses and a Unique Tymo-Like Virus in Hypovirulent and Virulent Strains of the Fusarium Head Blight Fungus, Fusarium boothii

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culture
2.2. Biological Assessment of FGSC Strains
2.3. Genomic Polymerase Chain Reaction
2.4. dsRNA Extraction and cDNA Library Construction
2.5. RNA Seq, Reverse Transcription PCR and Rapid Amplification of cDNA Ends
2.6. Bioinformatics Analyses
2.7. Phylogenetic Analysis
3. Results
3.1. Detection of dsRNAs in F. boothii Strains
3.2. Biological Characteristics of F. boothii Strains
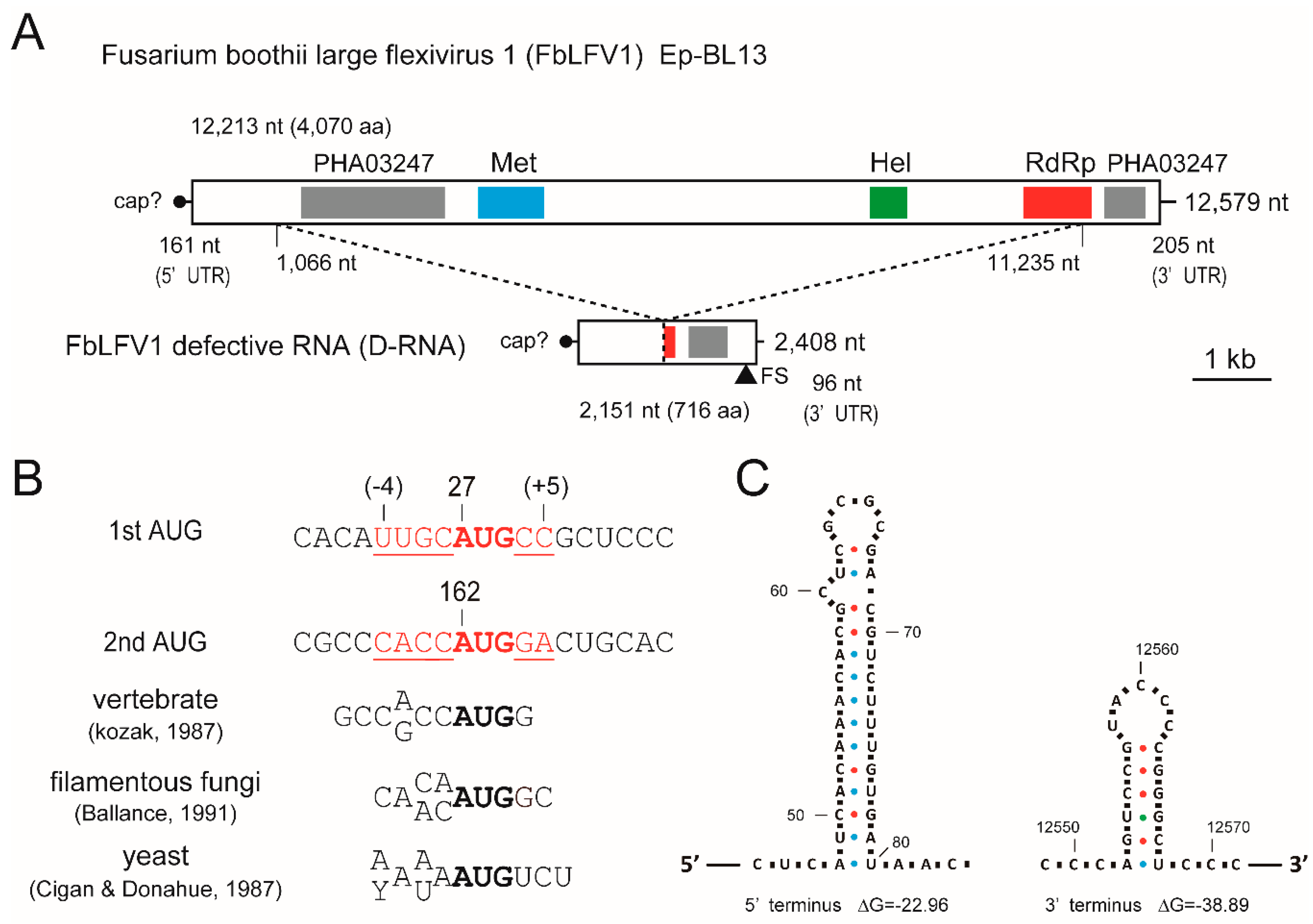
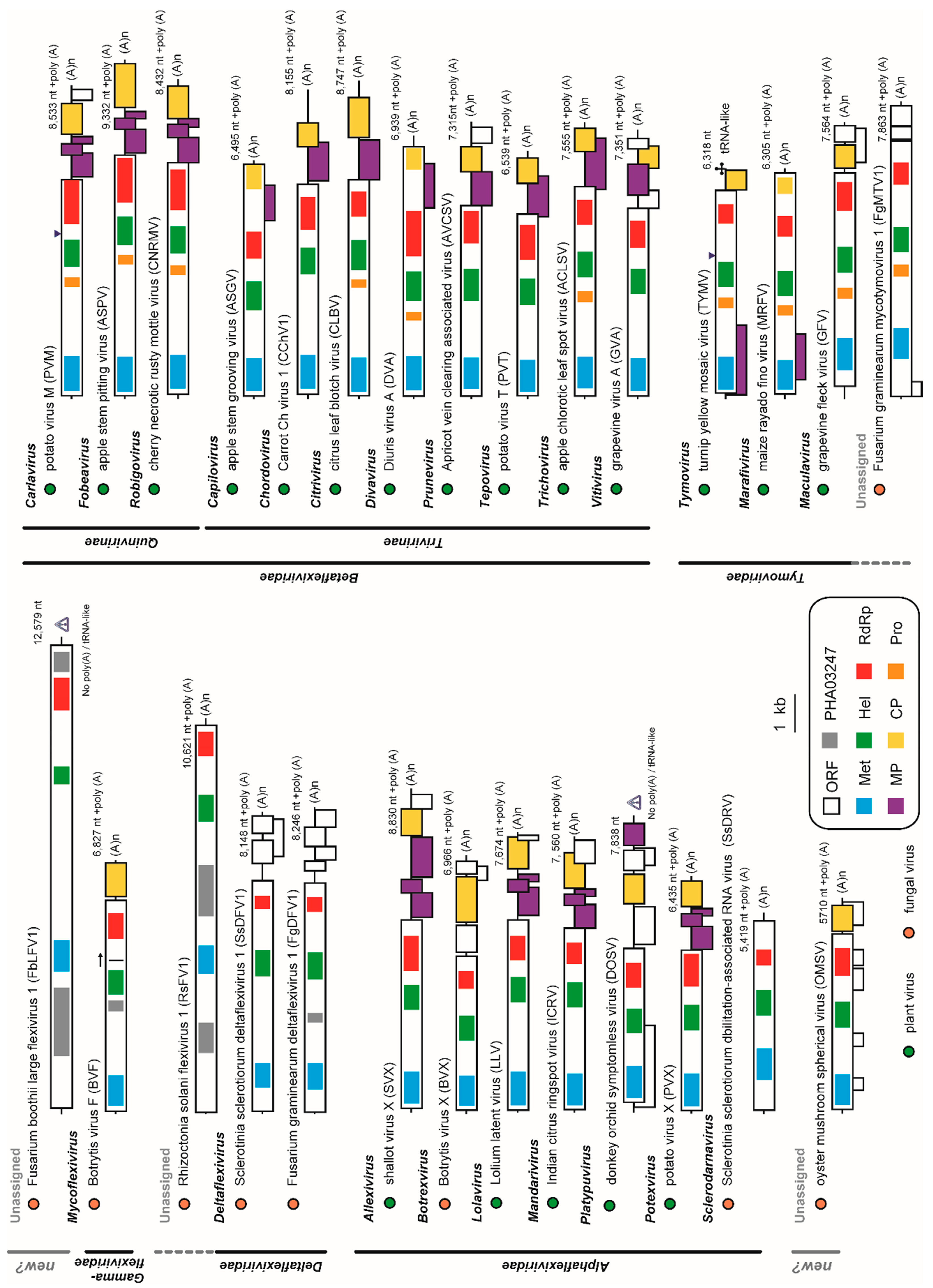
3.3. Analysis of Viral Sequences: A Tymo-Like Virus
3.4. Presence of a Defective Segment of FbFV1 in the Ep-BL13
3.5. Predicted RNA Structure of the Terminal Regions of FbLFV1 Genome
3.6. Molecular Phylogenetic Analysis of FbLFV1
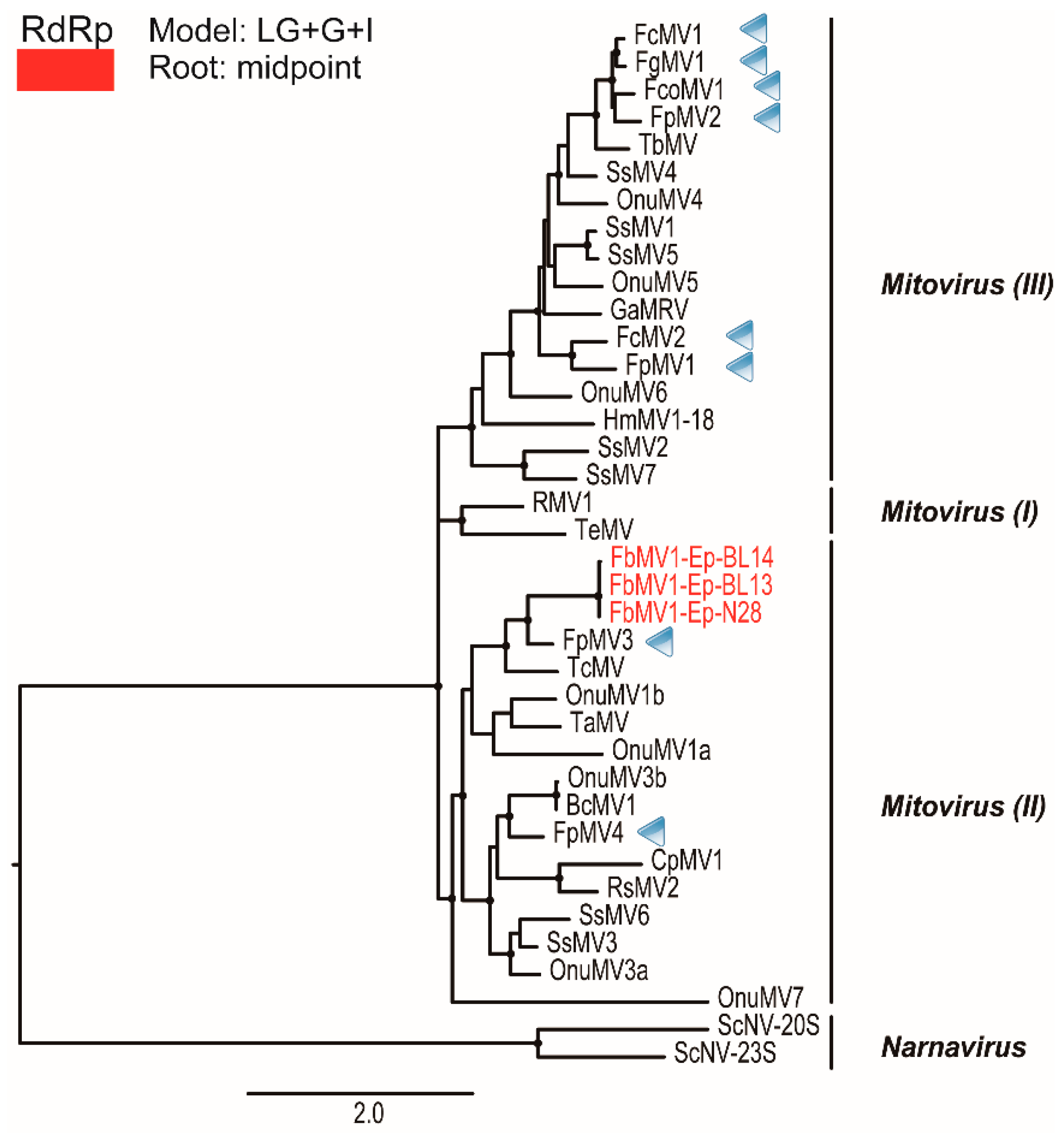
3.7. Analysis of Viral Sequences: Three Mitoviruses
3.8. Molecular Phylogenetic Analysis of Mitoviral RdRps
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479–480, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Dong, L.; Lin, L.; Ochoa, W.F.; Sinkovits, R.S.; Havens, W.M.; Nibert, M.L.; Baker, T.S.; Ghabrial, S.A.; Tao, Y.J. Atomic structure reveals the unique capsid organization of a dsRNA virus. Proc. Natl. Acad. Sci. USA 2009, 106, 4225–4230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillman, B.I.; Cai, G. The Family Narnaviridae: Simplest of RNA Viruses. Adv. Virus Res. 2013, 86, 149–176. [Google Scholar] [PubMed]
- Hillman, B.I.; Suzuki, N. Viruses of the chestnut blight fungi, Cryphonectria parasitica. Adv. Virus Res. 2004, 63, 423–472. [Google Scholar] [PubMed]
- Suzaki, K.; Ikeda, K.; Sasaki, A.; Kanematsu, S.; Matsumoto, N.; Yoshida, K. Horizontal transmission and host-virulence attenuation of totivirus in violet root rot fungus Helicobasidium mompa. J. Gen. Plant Pathol. 2005, 71, 161–168. [Google Scholar] [CrossRef]
- Moleleki, N.; van Heerden, S.W.; Wingfield, M.J.; Wingfield, B.D.; Preisig, O. Transfection of Diaporthe perjuncta with Diaporthe RNA virus. Appl. Environ. Microbiol. 2003, 69, 3952–3956. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.D.; Zhang, L.; Li, G.Q.; Jiang, D.H.; Hou, M.S.; Huang, H.C. Hypovirulence and Double-Stranded RNA in Botrytis cinerea. Phytopathology 2007, 97, 1590–1599. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.; Kramer, K.; Valdivia, L.; Ortiz, S.; Castillo, A. A double-stranded RNA mycovirus confers hypovirulence-associated traits to Botrytis cinerea. FEMS Microbiol. Lett. 2003, 228, 87–91. [Google Scholar] [CrossRef]
- Chu, Y.M.; Jeon, J.J.; Yea, S.J.; Kim, Y.H.; Yun, S.H.; Lee, Y.W.; Kim, K.H. Double-stranded RNA mycovirus from Fusarium graminearum. Appl. Environ. Microbiol. 2002, 68, 2529–2534. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Dover, S.L.; Cole, T.E.; Brasier, C.M.; Buck, K.W. Multiple Mitochondrial Viruses in an Isolate of the Dutch Elm Disease Fungus Ophiostoma novo-ulmi. Virology 1999, 258, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, B.; Fu, Y.; Jiang, D.; Ghabrial, S.A.; Li, G.; Peng, Y.; Xie, J.; Cheng, J.; Huang, J.; et al. A geminivirus-related DNA mycovirus that confers hypovirulence to a plant pathogenic fungus. Proc. Natl. Acad. Sci. USA 2010, 107, 8387–8392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Wet, J.; Bihon, W.; Preisig, O.; Wingfield, B.D.; Wingfield, M.J. Characterization of a novel dsRNA element in the pine endophytic fungus Diplodia scrobiculata. Arch. Virol. 2011, 156, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Preisig, O.; Wingfield, B.D.; Wingfield, M.J. Coinfection of a Fungal Pathogen by Two Distinct Double-Stranded RNA Viruses. Virology 1998, 252, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Vainio, E.J.; Martínez-Álvarez, P.; Bezos, D.; Hantula, J.; Diez, J.J. Fusarium circinatum isolates from northern Spain are commonly infected by three distinct mitoviruses. Arch. Virol. 2015, 160, 2093–2098. [Google Scholar] [CrossRef] [PubMed]
- Smit, W.A.; Wingfield, B.D.; Wingfield, M.J. Reduction of laccase activity and other hypovirulence-associated traits in dsRNA-containig strains of Diaporthe ambigua. Phytopathology 1996, 86, 1311–1316. [Google Scholar] [CrossRef]
- Preisig, O.; Moleleki, N.; Smit, W.A.; Wingfield, B.D.; Wingfield, M.J. A novel RNA mycovirus in a hypovirulent isolate of the plant pathogen Diaporthe ambigua. J. Gen. Virol. 2000, 81, 3107–3114. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, B.; Freeborough, M.J.; Maree, H.J.; Celton, J.M.; Rees, D.J.G.; Burger, J.T. Deep sequencing analysis of viruses infecting grapevines: Virome of a vineyard. Virology 2010, 400, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Darissa, O.; Adam, G.; Schäfer, W. A dsRNA mycovirus causes hypovirulence of Fusarium graminearum to wheat and maize. Eur. J. Plant Pathol. 2012, 134, 181–189. [Google Scholar] [CrossRef]
- Wang, L.; He, H.; Wang, S.; Chen, X.; Qiu, D.; Kondo, H.; Guo, L. Evidence for a novel negative-stranded RNA mycovirus isolated from the plant pathogenic fungus Fusarium graminearum. Virology 2018, 518, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Lin, Y.; Zhang, H.; Wang, S.; Qiu, D.; Guo, L. Molecular characterization of a novel mycovirus of the family Tymoviridae isolated from the plant pathogenic fungus Fusarium graminearum. Virology 2016, 489, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhang, H.; Chen, X.; Qiu, D.; Guo, L. Molecular characterization of a novel hypovirus from the plant pathogenic fungus Fusarium graminearum. Virology 2015, 481, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.S.; Lee, K.M.; Son, M.I.; Kim, K.H. Molecular Characterization of Fusarium Graminearum Virus 2 Isolated from Fusarium graminearum Strain 98-8-60. Plant Pathol. J. 2011, 27, 285–290. [Google Scholar] [CrossRef]
- Kwon, S.J.; Lim, W.S.; Park, S.H.; Park, M.R.; Kim, K.H. Molecular Characterization of a dsRNA Mycovirus, Fusarium graminearum Virus-DK21, which Is Phylogenetically Related to Hypoviruses but Has a Genome Organization and Gene Expression Strategy Resembling Those of Plant Potex-like Viruses. Mol. Cells 2007, 23, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.S.; Kwon, S.J.; Lee, K.M.; Son, M.; Kim, K.H. Complete nucleotide sequence of double-stranded RNA viruses from Fusarium graminearum strain DK3. Arch. Virol. 2009, 154, 1855–1858. [Google Scholar] [CrossRef] [PubMed]
- Osaki, H.; Sasaki, A.; Nomiyama, K.; Tomioka, K. Multiple virus infection in a single strain of Fusarium poae shown by deep sequencing. Virus Genes 2016, 52, 835–847. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Kondo, H.; Liu, L.; Guo, L.; Qiu, D. A novel virus in the family Hypoviridae from the plant pathogenic fungus Fusarium graminearum. Virus Res. 2013, 174, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; He, H.; Yang, X.; Zeng, H.; Qiu, D.; Guo, L. The complete genome sequence of a novel Fusarium graminearum RNA virus in a new proposed family within the order Tymovirales. Arch. Virol. 2016, 161, 2899–2903. [Google Scholar] [CrossRef] [PubMed]
- Marvelli, R.A.; Hobbs, H.A.; Li, S.; McCoppin, N.K.; Domier, L.L.; Hartman, G.L.; Eastburn, D.M. Identification of novel double-stranded RNA mycoviruses of Fusarium virguliforme and evidence of their effects on virulence. Arch. Virol. 2014, 159, 349–352. [Google Scholar] [CrossRef] [PubMed]
- King, A.M.Q.; Adams, M.J.; Carstens, E.B.; Lefkowitz, E.J. Narnaviridae. In Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 1055–1060. [Google Scholar]
- Polashock, J.J.; Hillman, B.I. A small mitochondrial double-stranded (ds) RNA element associated with a hypovirulent strain of the chestnut blight fungus and ancestrally related to yeast cytoplasmic T and W dsRNAs. Proc. Natl. Acad. Sci. USA 1994, 91, 8680–8684. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, M.E.; Pearson, M.N. Molecular characterization of three mitoviruses co-infecting a hypovirulent isolate of Sclerotinia sclerotiorum fungus. Virology 2013, 441, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Zhang, L.; Li, G.; Jiang, D.; Ghabrial, S.A. Genome characterization of a debilitation-associated mitovirus infecting the phytopathogenic fungus Botrytis cinerea. Virology 2010, 406, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Ghabrial, S.A. Molecular characterizations of two mitoviruses co-infecting a hyovirulent isolate of the plant pathogenic fungus Sclerotinia sclerotiorum. Virology 2012, 428, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Wu, S.; Liu, L.; Cheng, J.; Fu, Y.; Jiang, D.; Xie, J. A mitovirus related to plant mitochondrial gene confers hypovirulence on the phytopathogenic fungus Sclerotinia sclerotiorum. Virus Res. 2015, 197, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Jian, J.; Lakshman, D.K.; Tavantzis, S.M. Association of Distinct Double-Stranded RNAs with Enhanced or Diminished Virulence in Rhizoctonia solani Infecting Potato. Mol. Plant-Microbe Interact. 1997, 10, 1002–1009. [Google Scholar] [CrossRef]
- Martınetz, P.; Vainio, E.J.; Botella, L.; Hantula, J.; Diez, J.J. Three mitovirus strains infecting a single isolate of Fusarium circinatum are the first putative members of the family Narnaviridae detected in a fungus of the genus Fusarium. Arch. Virol. 2014, 159, 2153–2155. [Google Scholar] [CrossRef] [PubMed]
- Osaki, H.; Sasaki, A.; Nomiyama, K.; Sekiguchi, H.; Tomioka, K.; Takehara, T. Isolation and characterization of two mitoviruses and a putative alphapartitivirus from Fusarium spp. Virus Genes 2015, 50, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Bartholomäus, A.; Wibberg, D.; Winkler, A.; Pühler, A.; Schlüter, A.; Varrelmann, M. Identification of a novel mycovirus isolated from Rhizoctonia solani (AG 2-2 IV) provides further information about genome plasticity within the order Tymovirales. Arch. Virol. 2017, 162, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Howitt, R.L.J.; Beever, R.E.; Forster, R.L.S.; Pearson, M.N. Genome characterization of Botrytis virus F, a flexuous rod-shaped mycovirus resembling plant ‘potex-like’ viruses. J. Gen. Virol. 2001, 82, 67–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howitt, R.L.J.; Beever, R.E.; Pearson, M.N.; Forster, R.L.S. Genome characterization of a flexuous rod-shaped mycovirus, Botrytis virus X, reveals high amino acid identity to genes from plant ‘potex-like’ viruses. Arch. Virol. 2006, 151, 563–579. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Wei, D.; Jiang, D.; Fu, Y.; Li, G.; Ghabrial, S.; Peng, Y. Characterization of debilitation-associated mycovirus infecting the plant-pathogenic fungus Sclerotinia sclerotiorum. J. Gen. Virol. 2006, 87, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Zheng, D.; Cheng, J.; Chen, T.; Fu, Y.; Jiang, D.; Xie, J. Characterization of a novel Sclerotinia sclerotiorum RNA virus as the prototype of a new proposed family within the order Tymovirales. Virus Res. 2015, 219, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Bartholomäus, A.; Wibberg, D.; Winkler, A.; Pühler, A.; Schlüter, A.; Varrelmann, M. Deep sequencing analysis reveals the mycoviral diversity of the virome of an avirulent isolate of Rhizoctonia solani AG-2-2 IV. PLoS ONE 2016, 11, e0165965. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.J.; Lim, D.; Lee, H.S. Characterization of a novel single-stranded RNA mycovirus in pleurotus ostreatus. Virology 2003, 314, 9–15. [Google Scholar] [CrossRef]
- Hamid, M.; Xie, J.; Wu, S.; Maria, S.; Zheng, D.; Assane Hamidou, A.; Wang, Q.; Cheng, J.; Fu, Y.; Jiang, D.; et al. A Novel Deltaflexivirus that Infects the Plant Fungal Pathogen, Sclerotinia sclerotiorum, Can Be Transmitted Among Host Vegetative Incompatible Strains. Viruses 2018, 10, 295. [Google Scholar] [CrossRef] [PubMed]
- Mackintosh, C.A.; Garvin, D.F.; Radmer, L.E.; Heinen, S.J.; Muehlbauer, G.J. A model wheat cultivar for transformation to improve resistance to Fusarium Head Blight. Plant Cell Rep. 2006, 25, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Suga, H.; Kageyama, K.; Shimizu, M.; Hyakumachi, M. A Natural Mutation Involving both Pathogenicity and Perithecium Formation in the Fusarium graminearum Species Complex. G3 (Bethesda). 2016, 6, 3883–3892. [Google Scholar] [CrossRef] [PubMed]
- Ban, T.; Suenaga, K. Genetic analysis of resistance to Fusarium head blight caused by Fusarium graminearum in Chinese wheat cultivar Sumai 3 and the Japanese cultivar Saikai 165. Euphytica 2000, 113, 87–99. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, K.; Cigelnik, E.; Nirenberg, H.I. Molecular Systematics and Phylogeography of the Gibberella fujikuroi Species Complex. Mycologia 1998, 90, 465–493. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 25 October 2018).
- Joshi, N.; Fass, J. Sickle: A Sliding-Window, Adaptive, Quality-Based Trimming Tool for FastQ Files (Version 1.33). Available online: https://github.com/najoshi/sickle (accessed on 25 October 2018).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, R.D.; Attwood, T.K.; Babbitt, P.C.; Bateman, A.; Bork, P.; Bridge, A.J.; Chang, H.Y.; Dosztányi, Z.; El-Gebali, S.; Fraser, M.; et al. InterPro in 2017—Beyond protein family and domain annotations. Nucleic Acids Res. 2017, 45, D190–D199. [Google Scholar] [CrossRef] [PubMed]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K.; Battistuzzi, F.U. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, A.M.Q.; Adams, M.J.; Carstens, E.B.; Lefkowitz, E.J. Tymovirales. In Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 920–941. [Google Scholar]
- Ballance, D.J. Sequences important for gene expression in filamentous fungi. Yeast 1986, 2, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Kozak, M. At least six nucleotides preceding the AUG initiator codon enhance translation in mammalian cells. J. Mol. Biol. 1987, 196, 947–950. [Google Scholar] [CrossRef]
- Mark Cigan, A.; Donahue, T.F. Sequence and structural features associated with translational initiator regions in yeast—A review. Gene 1987, 59, 1–18. [Google Scholar] [CrossRef]
- Dreher, T.W. Role of tRNA-like structures in controlling plant virus replication. Virus Res. 2009, 139, 217–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, D.; Dreher, T.W. The tRNA-like structure of Turnip yellow mosaic virus RNA is a 3′-translational enhancer. Virology 2004, 321, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef] [PubMed]
- Osaki, H.; Nakamura, H.; Nomura, K.; Matsumoto, N.; Yoshida, K. Nucleotide sequence of a mitochondrial RNA virus from the plant pathogenic fungus, Helicobasidium mompa Tanaka. Virus Res. 2005, 107, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Marzano, S.Y.L.; Domier, L.L. Novel mycoviruses discovered from metatranscriptomics survey of soybean phyllosphere phytobiomes. Virus Res. 2016, 213, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, M.E.; Varsani, A.; Ganley, A.R.D.; Pearson, M.N. Comparison of Illumina de novo assembled and Sanger sequenced viral genomes: A case study for RNA viruses recovered from the plant pathogenic fungus Sclerotinia sclerotiorum. Virus Res. 2016, 219, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Mu, F.; Xie, J.; Cheng, S.; You, M.P.; Barbetti, M.J.; Jia, J.; Wang, Q.; Cheng, J.; Fu, Y.P.; Chen, T.; et al. Virome Characterization of a Collection of S. sclerotiorum from Australia. Front. Microbiol. 2018, 8, 2540. [Google Scholar] [CrossRef] [PubMed]
- Arjona-Lopez, J.M.; Telengech, P.; Jamal, A.; Hisano, S.; Kondo, H.; Yelin, M.D.; Arjona-Girona, I.; Kanematsu, S.; Lopez-Herrera, C.J.; Suzuki, N. Novel, diverse RNA viruses from Mediterranean isolates of the phytopathogenic fungus, Rosellinia necatrix: Insights into evolutionary biology of fungal viruses. Environ. Microbiol. 2018, 20, 1464–1483. [Google Scholar] [CrossRef] [PubMed]
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of filamentous fungi and their relevance to plant pathology. Mol. Plant Pathol. 2009, 10, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Wylie, S.J.; Li, H.; Jones, M.G.K. Donkey Orchid Symptomless Virus: A Viral ‘Platypus’ from Australian Terrestrial Orchids. PLoS ONE 2013, 8, e79587. [Google Scholar] [CrossRef] [PubMed]
- Tzanetakis, I.E.; Tsai, C.H.; Martin, R.R.; Dreher, T.W. A tymovirus with an atypical 3′-UTR illuminates the possibilities for 3′-UTR evolution. Virology 2009, 392, 238–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, W.H.; Roberts, A.P.E.; McElwee, M.; Bhella, D.; Rixon, F.J.; Lauder, R. The large tegument protein pUL36 is essential for formation of the capsid vertex-specific component at the capsid-tegument interface of herpes simplex virus 1. J. Virol. 2015, 89, 1502–1511. [Google Scholar] [CrossRef] [PubMed]
- Lakshman, D.K.; Jian, J.; Tavantzis, S.M. A double-stranded RNA element from a hypovirulent strain of Rhizoctonia solani occurs in DNA form and is genetically related to the pentafunctional AROM protein of the shikimate pathway (RNA-dependent RNA polymerase). Microbiology 1998, 95, 6425–6429. [Google Scholar] [CrossRef]
- Deng, F.; Xu, R.; Boland, G.J. Hypovirulence-Associated Double-Stranded RNA from Sclerotinia homoeocarpa Is Conspecific with Ophiostoma novo-ulmi Mitovirus 3a-Ld. Phytopathology 2003, 93, 1407–1414. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Yu, J.; Son, M.; Lee, Y.W.; Kim, K.H.; Arkowitz, R.A. Transmission of Fusarium boothii Mycovirus via Protoplast Fusion Causes Hypovirulence in Other Phytopathogenic Fungi. PLoS ONE 2011, 6, e21629. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Virus Name (Abbreviation) | Accession | Met * | Hel * | RdRp * |
|---|---|---|---|---|---|
| Alphaflexiviridae | |||||
| Allexivirus | Shallot virus X (SVX) | NC_003795 | 58/198 (29%) | 37/154 (24%) | 74/221 (33%) |
| Botrexvirus | Botrytis virus X (BVX) | NC_005132 | 60/196 (30%) | 45/178 (25%) | 76/221 (34%) |
| Lolavirus | Lolium latent virus (LLV) | NC_010434 | 64/193 (33%) | 50/241 (20%) | 80/238 (33%) |
| Mandarivirus | Indian citrus ringspot virus (ICRV) | NC_003093 | 59/162 (36%) | 63/244 (25%) | 84/229 (36%) |
| Platypuvirus | Donkey orchid symptomless virus (DOSV) | NC_022894 | 60/201 (29%) | 49/189 (25%) | 69/225 (30%) |
| Potexvirus | Potato virus X (PVX) | NC_011620 | 60/198 (30%) | 58/245 (23%) | 79/227 (34%) |
| Sclerodarnavirus | Sclerotinia sclerotiorum debilitation-associatedRNA virus (SsDARV) | NC_007415 | 63/198 (31%) | 49/191 (25%) | 71/227 (31%) |
| Betaflexiviridae | |||||
| Quinvirinae | |||||
| Carlavirus | Potato virus M (PVM) | NC_001361 | 55/217 (25%) | 35/127 (27%) | 62/224 (27%) |
| Foveavirus | Apple stem pitting virus (ASPV) | NC_001749 | 60/222 (27%) | 17/45 (37%) | 63/222 (28%) |
| Robigovirus | Cherry necrotic rusty mottle virus (CNRMV) | NC_002468 | 52/224 (23%) | 44/159 (27%) | 61/219 (27%) |
| Trivirinae | |||||
| Capillovirus | Apple stem grooving virus (ASGV) | NC_003462 | 64/214 (29%) | 33/97 (34%) | 72/231 (31%) |
| Chordovirus | Carrot Ch virus 1 (CChV) | NC_025469 | 54/214 (25%) | 33/118 (27%) | 66/225 (29%) |
| Citrivirus | Citrus leaf blotch virus (CLBV) | NC_003877 | 46/205 (22%) | 46/160 (28%) | 68/223 (30%) |
| Divavirus | Diuris virus A (DVA) | NC_019029 | 53/217 (24%) | 26/90 (28%) | 66/224 (29%) |
| Prunevirus | Apricot vein clearing associated virus (AVCSV) | NC_023295 | 49/183 (26%) | 24/62 (38%) | 66/224 (29%) |
| Tepovirus | Potato virus T (PVT) | NC_011062 | 56/186 (30%) | 34/115 (29%) | 67/219 (30%) |
| Trichovirus | Apple chlorotic leaf spot virus (ACLSV) | NC_001409 | 62/195 (31%) | 43/172 (25%) | 64/218 (29%) |
| Vitivirus | Grapevine virus A (GVA) | NC_003604 | 50/201 (24%) | 22/57 (38%) | 65/218 (29%) |
| Gammaflexiviridae | |||||
| Mycoflexivirus | Botrytis virus F (BVF) | NC_002604 | 63/193 (32%) | 71/252 (28%) | 82/224 (36%) |
| Deltaflexiviridae | |||||
| Deltaflexivirus | Sclerotinia sclerotiorum deltaflexivirus 1 (SsDFV1) | NC_038977 | 54/202 (26%) | 61/253 (24%) | 45/174 (25%) |
| Sclerotinia sclerotiorum deltaflexivirus 2 (SsDFV2) | MH299810 | 63/222 (28%) | 44/184 (23%) | 59/207 (28%) | |
| Fusarium graminearum deltaflexivirus 1 (FgDFV1) | NC_030654 | 67/204 (32%) | 37/157 (23%) | 57/211 (27%) | |
| Soybean leaf-associated deltaflexivirus 1 (SlaDFV1) | KT598226 | 54/203 (26%) | 63/260 (24%) | 46/174 (26%) | |
| Tymoviridae | |||||
| Maculavirus | Grapevine fleck virus (GFV) | NC_003347 | 57/203 (28%) | 43/144 (29%) | 54/143 (37%) |
| Marafivirus | Maize rayado fino virus (MRFV) | NC_002786 | 58/197 (29%) | 57/201 (28%) | 45/140 (32%) |
| Tymovirus | Turnip yellow mosaic virus (TUMV) | NC_004063 | 56/196 (28%) | 58/214 (27%) | 61/204 (29%) |
| Unassigned | |||||
| Mycotymovirus | Fusarium graminearum mycotymovirus 1 (FgMTV1) | KT360947 | 60/196 (30%) | 57/197 (26%) | 73/219 (33%) |
| Unassigned | Rhizoctonia solani flexivirus 1 (RnFV1) | NC_030655 | 57/198 (28%) | 53/228 (23%) | 65/224 (29%) |
| Unassigned | Rhizoctonia solani flexivirus 2 (RnFV1) | KX349069 | N/A ** | N/A ** | 50/177 (28%) |
| Unassigned | Oyster muschroom spherical virus (OMSV) | AY182001 | 48/189 (25%) | 61/237 (25%) | 73/219 (33%) |
| Unassigned | Fusarium boothii large flexivirus 1 (FbLFV1) | LC425115 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mizutani, Y.; Abraham, A.; Uesaka, K.; Kondo, H.; Suga, H.; Suzuki, N.; Chiba, S. Novel Mitoviruses and a Unique Tymo-Like Virus in Hypovirulent and Virulent Strains of the Fusarium Head Blight Fungus, Fusarium boothii. Viruses 2018, 10, 584. https://doi.org/10.3390/v10110584
Mizutani Y, Abraham A, Uesaka K, Kondo H, Suga H, Suzuki N, Chiba S. Novel Mitoviruses and a Unique Tymo-Like Virus in Hypovirulent and Virulent Strains of the Fusarium Head Blight Fungus, Fusarium boothii. Viruses. 2018; 10(11):584. https://doi.org/10.3390/v10110584
Chicago/Turabian StyleMizutani, Yukiyoshi, Adane Abraham, Kazuma Uesaka, Hideki Kondo, Haruhisa Suga, Nobuhiro Suzuki, and Sotaro Chiba. 2018. "Novel Mitoviruses and a Unique Tymo-Like Virus in Hypovirulent and Virulent Strains of the Fusarium Head Blight Fungus, Fusarium boothii" Viruses 10, no. 11: 584. https://doi.org/10.3390/v10110584