Identification of a cis-Acting Element Derived from Tomato Leaf Curl Yunnan Virus that Mediates the Replication of a Deficient Yeast Plasmid in Saccharomyces cerevisiae



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Stains
2.2. Plasmid Construction
2.3. Yeast Transformation
2.4. Plasmid Stability Assay
2.5. Southern Blot Assay
3. Results
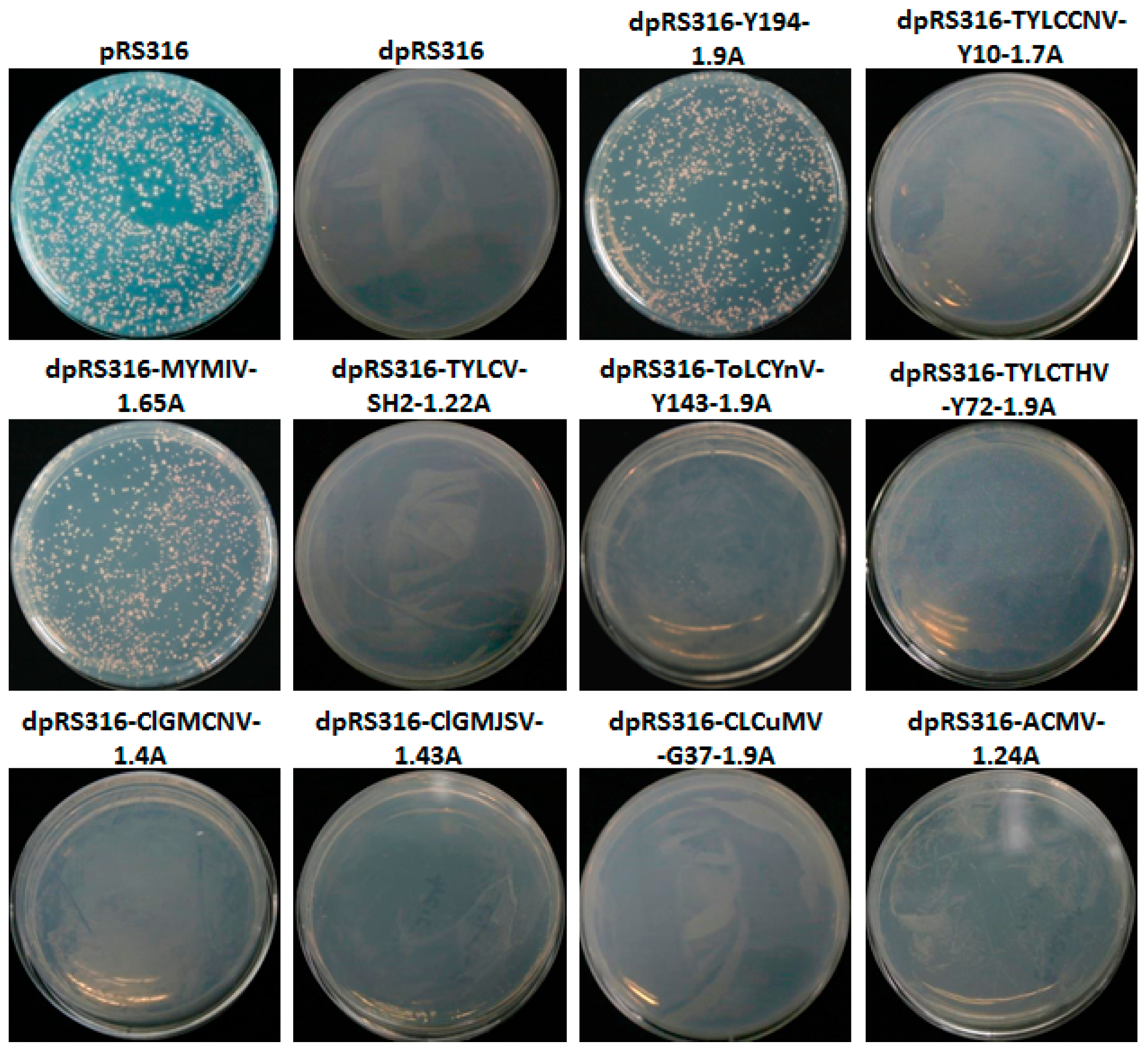
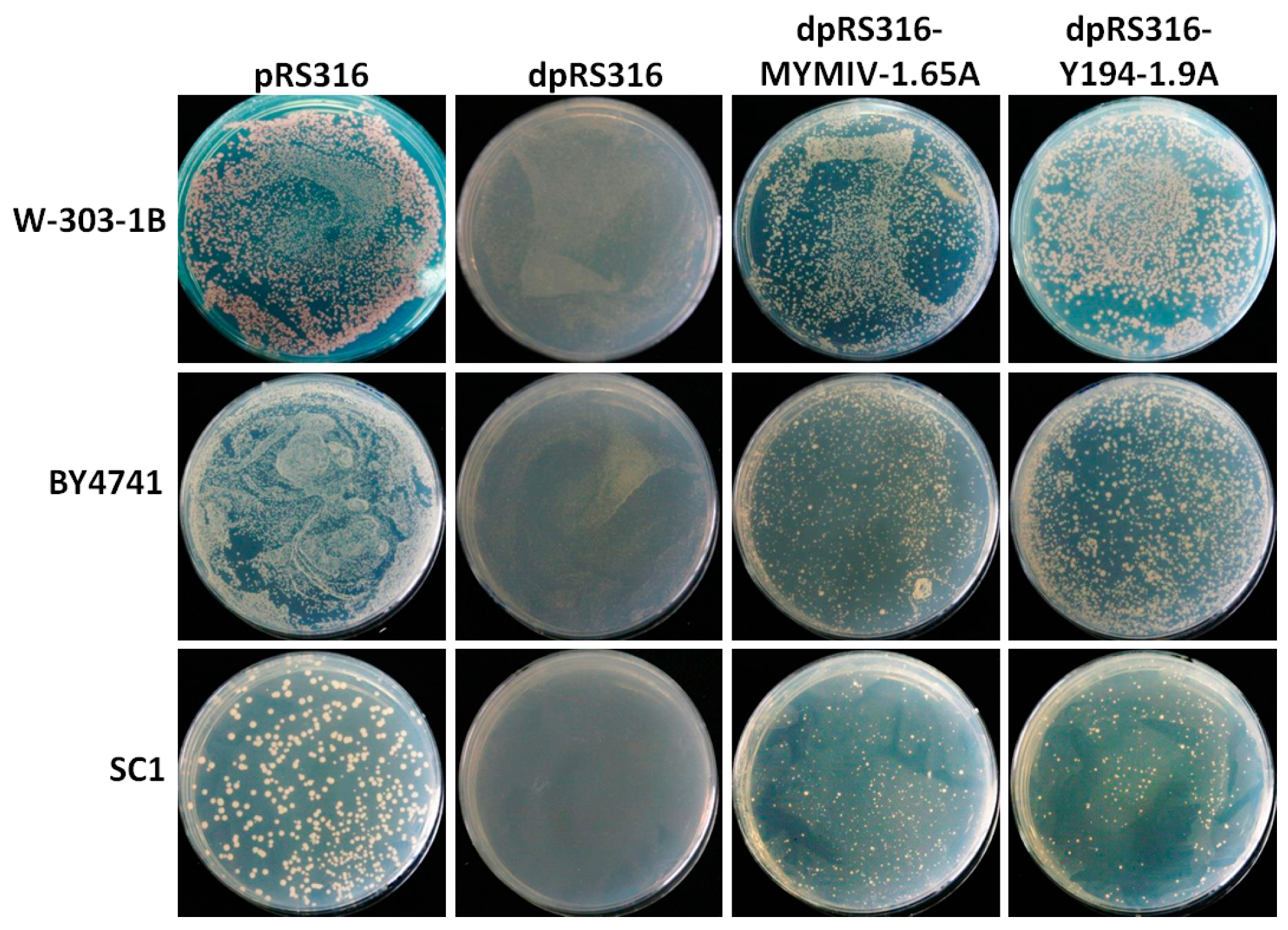
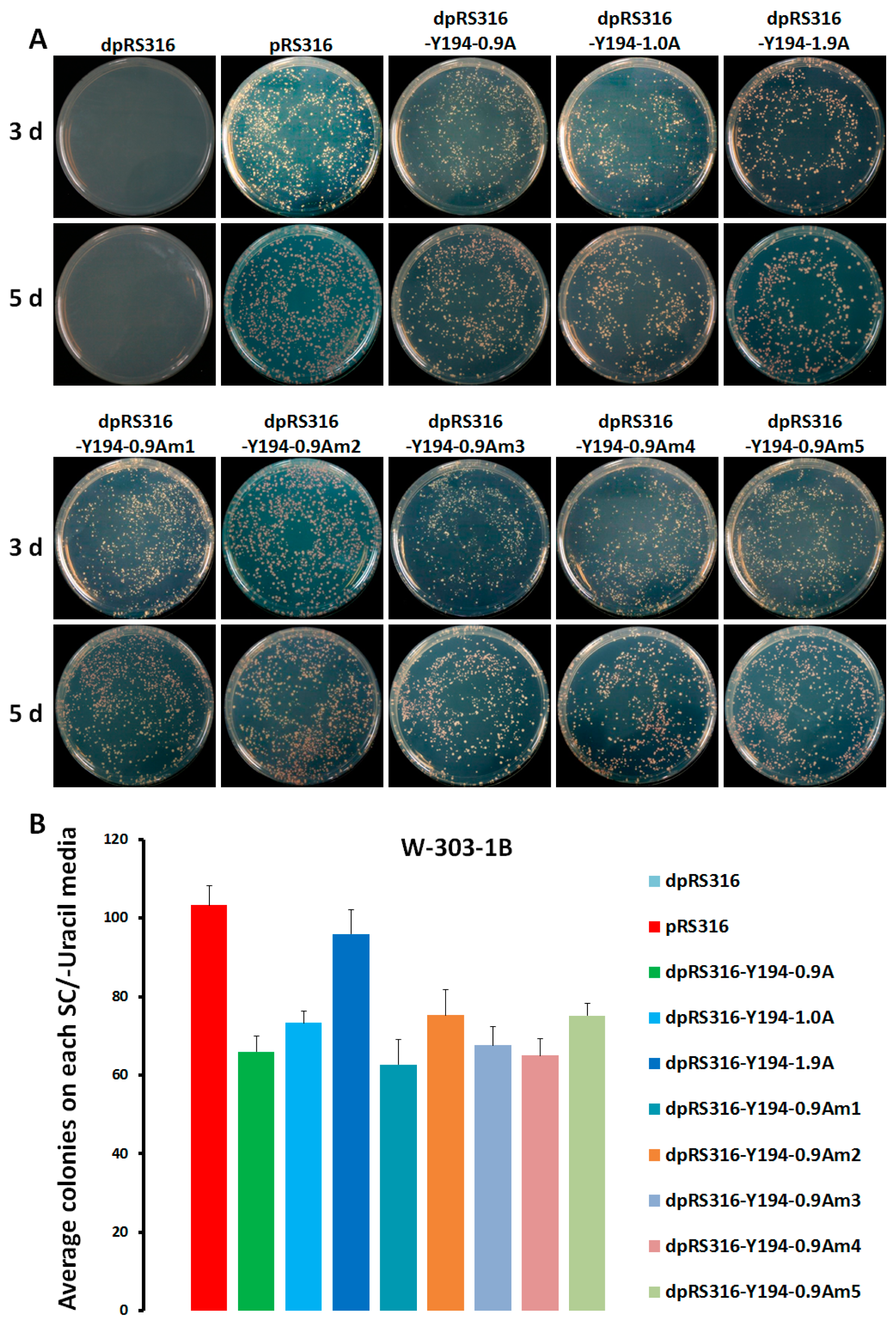
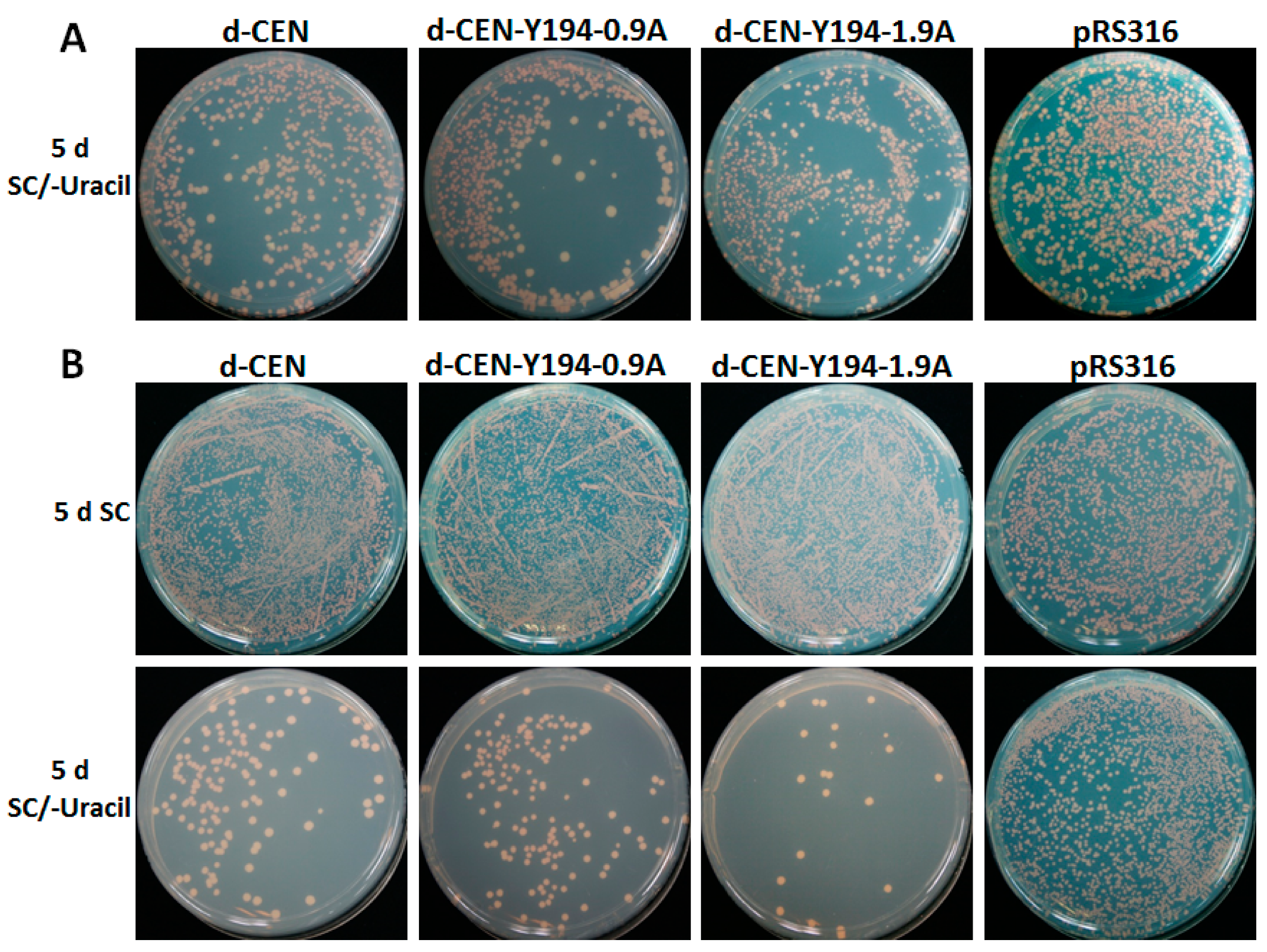
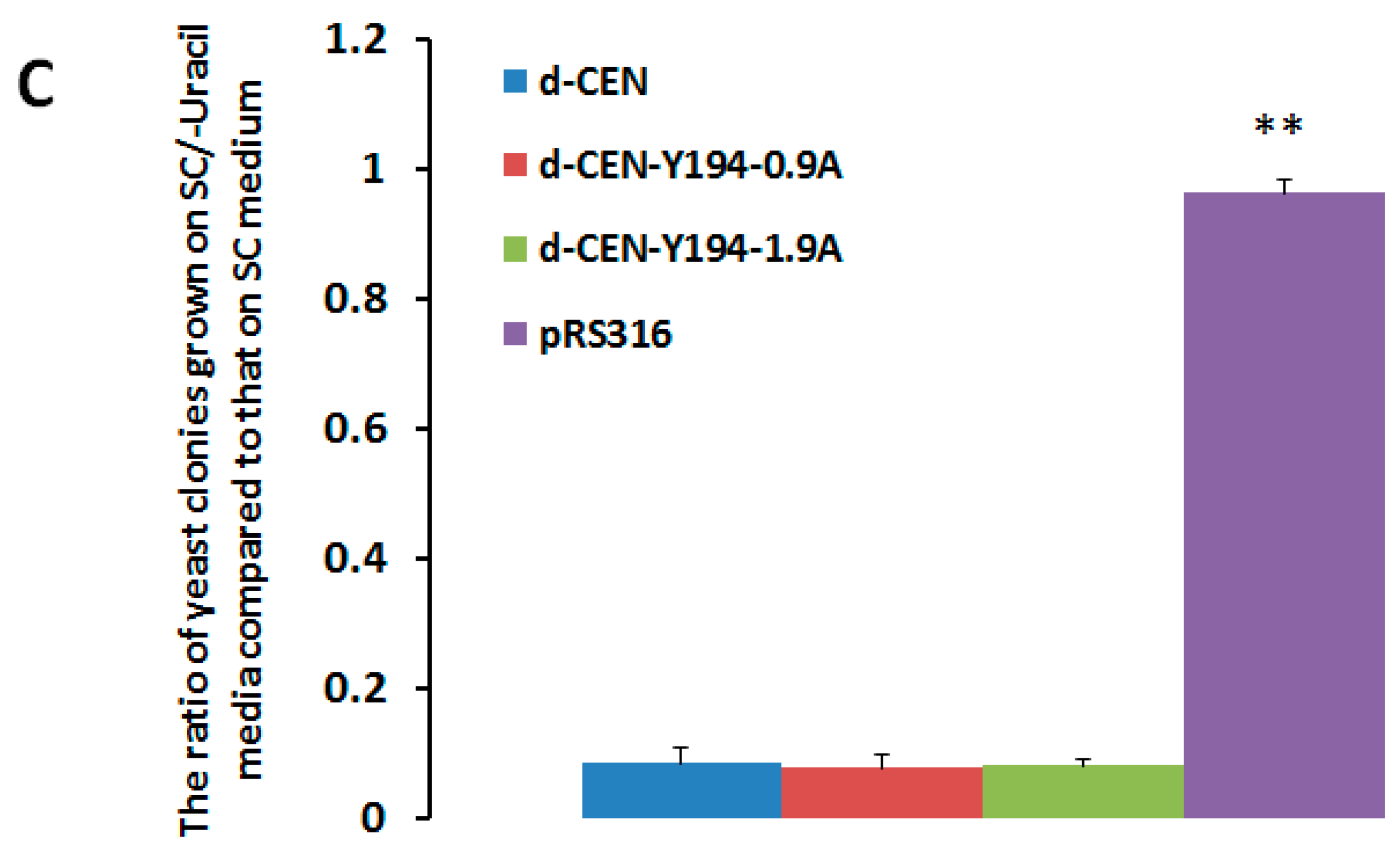
3.1. The Recombinant Plasmid Consisting of 1.9 Tandem Copies of the Y194 Genome Can Replicate in Yeast
3.2. A Non-RCR Model Supporting the Replication of the Recombinant Plasmid with Y194 DNA in Yeast Cells
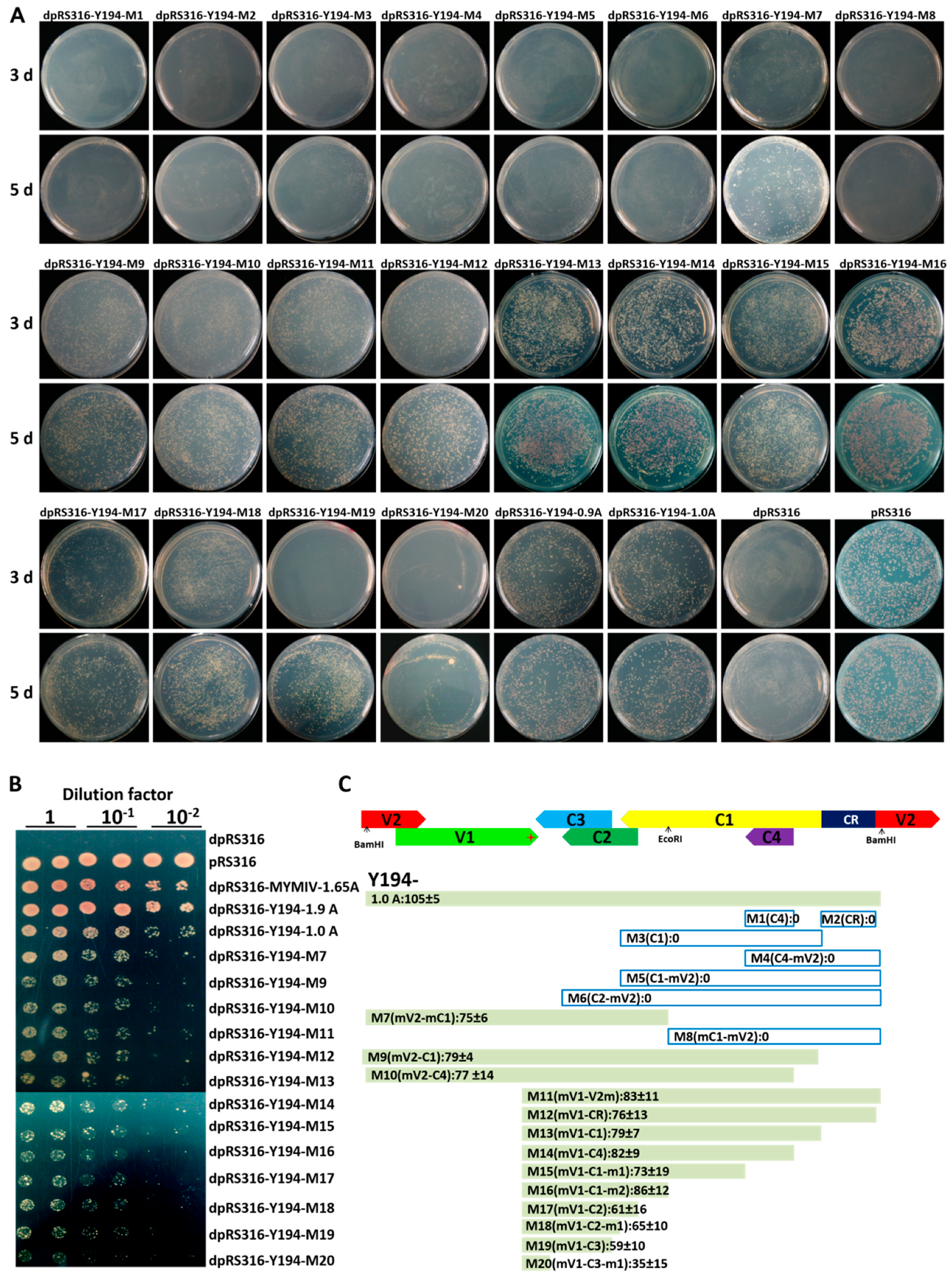
3.3. A 174-nt Sequence Located in between the V1 and C3 ORF Includes an ARS-Like Element in the Y194 Genome
3.4. Y194 Genome Lacks a Centromere (CEN)-Like Element
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Varsani, A.; Roumagnac, P.; Fuchs, M.; Navas-Castillo, J.; Moriones, E.; Idris, A.; Briddon, R.W.; Rivera-Bustamante, R.; Zerbini, F.M.; Martin, D.P. Capulavirus and Grablovirus: Two new genera in the family Geminiviridae. Arch. Virol. 2017, 162, 1819–1831. [Google Scholar] [CrossRef] [PubMed]
- Sanderfoot, A.A.; Lazarowitz, S.G. Getting it together in plant virus movement: Cooperative interactions between bipartite geminivirus movement proteins. Trends Cell Biol. 1996, 6, 353–358. [Google Scholar] [CrossRef]
- Hanley-Bowdoin, L.; Settlage, S.B.; Orozco, B.M.; Nagar, S.; Robertson, D. Geminiviruses: Models for plant DNA replication, transcription, and cell cycle regulation. Crit. Rev. Plant Sci. 2010, 18, 71–106. [Google Scholar] [CrossRef]
- Zhou, X. Advances in understanding Begomovirus satellites. Annu. Rev. Phytopathol. 2013, 51, 357–381. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xie, Y.; Raja, P.; Li, S.; Wolf, J.N.; Shen, Q.; Bisaro, D.M.; Zhou, X. Suppression of methylation-mediated transcriptional gene silencing by βC1-SAHH protein interaction during geminivirus-betasatellite infection. PLoS Pathog. 2011, 7, e1002329. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Huang, C.; Li, Z.; Zhou, X. Suppression of RNA silencing by a plant DNA virus satellite requires a host calmodulin-like protein to repress RDR6 expression. PLoS Pathog. 2014, 10, e1003921. [Google Scholar] [CrossRef] [PubMed]
- Saunders, K.; Lucy, A.; Stanley, J. DNA forms of the geminivirus African cassava mosaic virus consistent with a rolling-circle mechanism of replication. Nucleic Acids Res. 1991, 19, 2325–2330. [Google Scholar] [CrossRef] [PubMed]
- Stenger, D.C.; Revington, G.N.; Stevenson, M.C.; Bisaro, D.M. Replicational release of geminivirus genomes from tandemly repeated copies: Evidence for rolling-circle replication of a plant viral DNA. Proc. Natl. Acad. Sci. USA 1991, 88, 8029–8033. [Google Scholar] [CrossRef] [PubMed]
- Fontes, E.P.; Luckow, V.A.; Hanley-Bowdoin, L. A geminivirus replication protein is a sequence-specific DNA binding protein. Plant Cell 1992, 4, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Desbiez, C.; David, C.; Mettouchi, A.; Laufs, J.; Gronenborn, B. Rep protein of tomato yellow leaf curl geminivirus has an ATPase activity required for viral DNA replication. Proc. Natl. Acad. Sci. USA 1995, 92, 5640–5644. [Google Scholar] [CrossRef] [PubMed]
- Laufs, J.; Traut, W.; Heyraud, F.; Matzeit, V.; Rogers, S.G.; Schell, J.; Gronenborn, B. In vitro cleavage and joining at the viral origin of replication by the replication initiator protein of tomato yellow leaf curl virus. Proc. Natl. Acad. Sci. USA 1995, 3879–3883. [Google Scholar] [CrossRef]
- Stanley, J. Analysis of African cassava mosaic virus recombinants suggests strand nicking occurs within the conserved nonanucleotide motif during the initiation of rolling circle DNA replication. Virology 1995, 206, 707–712. [Google Scholar] [CrossRef]
- Choudhury, N.R.; Malik, P.S.; Singh, D.K.; Islam, M.N.; Kaliappan, K.; Mukherjee, S.K. The oligomeric Rep protein of mungbean yellow mosaic India virus (MYMIV) is a likely replicative helicase. Nucleic Acids Res. 2006, 34, 6362–6377. [Google Scholar] [CrossRef] [PubMed]
- Cleŕot, D.; Bernardi, F. DNA helicase activity is associated with the replication initiator protein Rep of tomato yellow leaf curl geminivirus. J. Virol. 2006, 80, 1322–11330. [Google Scholar] [CrossRef] [PubMed]
- Haley, A.; Zhan, X.; Richardson, K.; Head, K.; Morris, B. Regulation of the activities of African cassava mosaic virus promoters by the AC1, AC2, and AC3 gene products. Virology 1992, 188, 905–909. [Google Scholar] [CrossRef]
- Eagle, P.A.; Orozco, B.M.; Hanley-Bowdoin, L. A DNA sequence required for geminivirus replication also mediates transcriptional regulation. Plant Cell 1994, 6, 1157–1170. [Google Scholar] [CrossRef] [PubMed]
- Fontes, E.P.; Eagle, P.A.; Sipe, P.S.; Luckow, V.A. Hanley-Bowdoin, L. Interaction between a geminivirus replication protein and origin DNA is essential for viral replication. J. Biol. Chem. 1994, 269, 8459–8465. [Google Scholar] [PubMed]
- Gröning, B.R.; Hayes, R.J.; Buck, K.W. Simultaneous regulation of tomato golden mosaic virus coat protein and AL1 gene expression: Expression of the AL4 gene may contribute to suppression of the AL1 gene. J. Gen. Virol. 1994, 75, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Stanley, J. Regulation of African cassava mosaic virus complementary-sense gene expression by N-terminal sequences of the replication- associated protein AC1. J. Gen. Virol. 1995, 76, 2415–2422. [Google Scholar] [CrossRef] [PubMed]
- Rigden, J.E.; Krake, L.R.; Rezaian, M.A.; Dry, I.B. ORF C4 of tomato leaf curl geminivirus is a determinant of symptom severity. Virology 1994, 204, 847–850. [Google Scholar] [CrossRef] [PubMed]
- Vanitharani, R.; Chellappan, P.; Pita, J.S.; Fauquet, C.M. Differential roles of AC2 and AC4 of cassava geminiviruses in mediating synergism and suppression of posttranscriptional gene silencing. J. Virol. 2004, 78, 9487–9498. [Google Scholar] [CrossRef] [PubMed]
- Raghavan, V.; Malik, P.S.; Choudhury, N.R.; Mukherjee, S.K. The DNA-A component of a plant geminivirus (indian mung bean yellow mosaic virus) replicates in budding yeast cells. J. Virol. 2004, 78, 2405–2413. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Xu, X.; Huang, C.; Gu, Z.; Cao, L.; Hu, T.; Ding, M.; Li, Z.; Zhou, X. The AC5 protein encoded by Mungbean yellow mosaic India virus is a pathogenicity determinant that suppresses RNA silencing-based antiviral defenses. New Phytol. 2015, 208, 555–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanley-Bowdoin, L.; Settlage, S.B.; Robertson, D. Reprogramming plant gene expression: A prerequisite to geminivirus DNA replication. Mol. Plant Pathol. 2004, 5, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Sanz-Burgos, A.P.; Hannon, G.J.; Gutierrez, C. Plant cells contain a novel member of the retinoblastoma family of growth regulatory proteins. EMBO J. 1996, 15, 4900–4908. [Google Scholar] [CrossRef] [PubMed]
- Ach, R.A.; Durfee, T.; Miller, A.B.; Taranto, P.; Hanley-Bowdoin, L.; Zambriski, P.C.; Gruissem, W. RRB1 and RRB2 encode maize retinoblastoma-related proteins that interact with a plant D-type cyclin and geminivirus replication protein. Mol. Cell. Biol. 1997, 17, 5077–5086. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.J.; Orozco, B.M.; Roe, J.R.; Nagar, S.; Ou, S.; Feiler, H.S.; Durfee, T.; Miller, A.B.; Gruissem, W.; Robertson, D.; et al. A geminivirus replication protein interacts with the retinoblastoma protein through a novel domain to determine symptoms and tissue specificity of infection in plants. EMBO J. 2000, 19, 3485–3495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chellappan, S.P.; Hiebert, S.; Mudryj, M.; Horowitz, J.M.; Nevins, J.R. The E2F transcription factor is a cellular target for the RB protein. Cell 1991, 65, 1053–1061. [Google Scholar] [CrossRef]
- Egelkrout, E.M.; Robertson, D.; Hanley-Bowdoin, L. Proliferating cell nuclear antigen transcription is repressed through an E2F consensus element and activated by geminivirus infection in mature leaves. Plant Cell 2001, 13, 1437–1452. [Google Scholar] [CrossRef] [PubMed]
- Arguello-Astorga, G.; Lopez-Ochoa, L.; Kong, L.J.; Orozco, B.M.; Settlage, S.B.; Hanley-Bowdoin, L. A novel motif in geminiviral replication proteins interacts with the plant retinoblastoma related protein. J. Virol. 2004, 78, 4817–4826. [Google Scholar] [CrossRef] [PubMed]
- Bagewadi, B.; Chen, S.; Lal, S.K.; Choudhury, N.R.; Mukherjee, S.K. PCNA Interacts with Indian Mung Bean Yellow Mosaic Virus Rep and Down-regulates Rep Activity. J. Virol. 2004, 78, 11890–11903. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, Y.I.; Maki, S.; Maki, H.; Kunkel, T.A. Evidence for interplay among yeast replicative DNA polymerases alpha, delta and epsilon from studies of exonuclease and polymerase active site mutations. BMC Biol. 2004, 2, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loor, G.; Zhang, S.; Zhang, P.; Toomey, N.L.; Lee, M.Y. Identification of DNA replication and cell cycle proteins that interact with PCNA. Nucleic Acids Res. 1997, 25, 5041–5046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaliappan, K.; Choudhury, N.R.; Suyal, G.; Mukherjee, S.K. A novel role for RAD54: This host protein modulates geminiviral DNA replication. FASEB J. 2012, 26, 1142–1160. [Google Scholar] [CrossRef] [PubMed]
- Suyal, G.; Mukherjee, S.K.; Choudhury, N.R. The host factor RAD51 is involved in mungbean yellow mosaic India virus (MYMIV) DNA replication. Arch. Virol. 2013, 158, 1931–1941. [Google Scholar] [CrossRef] [PubMed]
- Suyal, G.; Mukherjee, S.K.; Srivastava, P.S.; Choudhury, N.R. Arabidopsis thaliana MCM2 plays role(s) in mungbean yellow mosaic India virus (MYMIV) DNA replication. Arch. Virol. 2013, 158, 981–992. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Negrete, E.; Lozano-Durán, R.; Piedra-Aguilera, A.; Cruzado, L.; Bejarano, E.R.; Castillo, A.G. Geminivirus Rep protein interferes with the plant DNA methylation machinery and suppresses transcriptional gene silencing. New Phytol. 2013, 199, 464–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panavas, T.; Nagy, P.D. Yeast as a model host to study replication and recombination of defective interfering RNA of tomato bushy stunt virus. Virology 2013, 314, 315–325. [Google Scholar] [CrossRef]
- Pantaleo, V.; Rubino, L.; Russo, M. Replication of carnation italian ringspot virus defective interfering RNA in Saccharomyces cerevisiae. J. Virol. 2003, 77, 2116–2123. [Google Scholar] [CrossRef] [PubMed]
- Nawaz-ul-Rehman, M.S.; Reddisiva, P.K.; Baker, J.; Nagy, P.D. Yeast screens for host factors in positive-strand RNA virus replication based on a library of temperature-sensitive mutants. Methods 2013, 59, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Zhao, L.; Jiao, X.; Jiang, T.; Gong, H.; Wang, B.; Briddon, R.W.; Zhou, X. A recombinant begomovirus resulting from exchange of the C4 gene. J. Gen. Virol. 2013, 94, 896–1907. [Google Scholar] [CrossRef] [PubMed]
- Angeletti, P.C.; Kim, K.; Fernandes, F.J.; Lambert, P.F. Stable replication of papilloma virus genomes in Saccharomyces cerevisiae. J. Virol. 2002, 76, 3350–3358. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Angeletti, P.C.; Hassebroek, E.C.; Lambert, P.F. Identification of cis-acting elements that mediate the replication and maintenance of human papillomavirus type 16 genomes in Saccharomyces cerevisiae. J. Virol. 2005, 79, 5933–5942. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Xu, X.; Yang, X.; Li, Z.; Zhou, X. Identification of a cis-Acting Element Derived from Tomato Leaf Curl Yunnan Virus that Mediates the Replication of a Deficient Yeast Plasmid in Saccharomyces cerevisiae. Viruses 2018, 10, 536. https://doi.org/10.3390/v10100536
Li F, Xu X, Yang X, Li Z, Zhou X. Identification of a cis-Acting Element Derived from Tomato Leaf Curl Yunnan Virus that Mediates the Replication of a Deficient Yeast Plasmid in Saccharomyces cerevisiae. Viruses. 2018; 10(10):536. https://doi.org/10.3390/v10100536
Chicago/Turabian StyleLi, Fangfang, Xiongbiao Xu, Xiuling Yang, Zhenghe Li, and Xueping Zhou. 2018. "Identification of a cis-Acting Element Derived from Tomato Leaf Curl Yunnan Virus that Mediates the Replication of a Deficient Yeast Plasmid in Saccharomyces cerevisiae" Viruses 10, no. 10: 536. https://doi.org/10.3390/v10100536
APA StyleLi, F., Xu, X., Yang, X., Li, Z., & Zhou, X. (2018). Identification of a cis-Acting Element Derived from Tomato Leaf Curl Yunnan Virus that Mediates the Replication of a Deficient Yeast Plasmid in Saccharomyces cerevisiae. Viruses, 10(10), 536. https://doi.org/10.3390/v10100536