Ostreid Herpesvirus-1 Infects Specific Hemocytes in Ark Clam, Scapharca broughtonii



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Ark Clams
2.2. Experimental OsHV-1 Infection and Sample Collection
2.3. Quantification of OsHV-1 Load by RT-qPCR
2.4. Flow Cytometry Analysis
2.5. Hemocyte Morphologic Observation
2.6. Transmission Electron Microscopy (TEM) Examination
3. Results
3.1. OsHV-1 Load in Different Ark Clam Tissues
3.2. Characterization of Different Types of Ark Clam Hemocytes
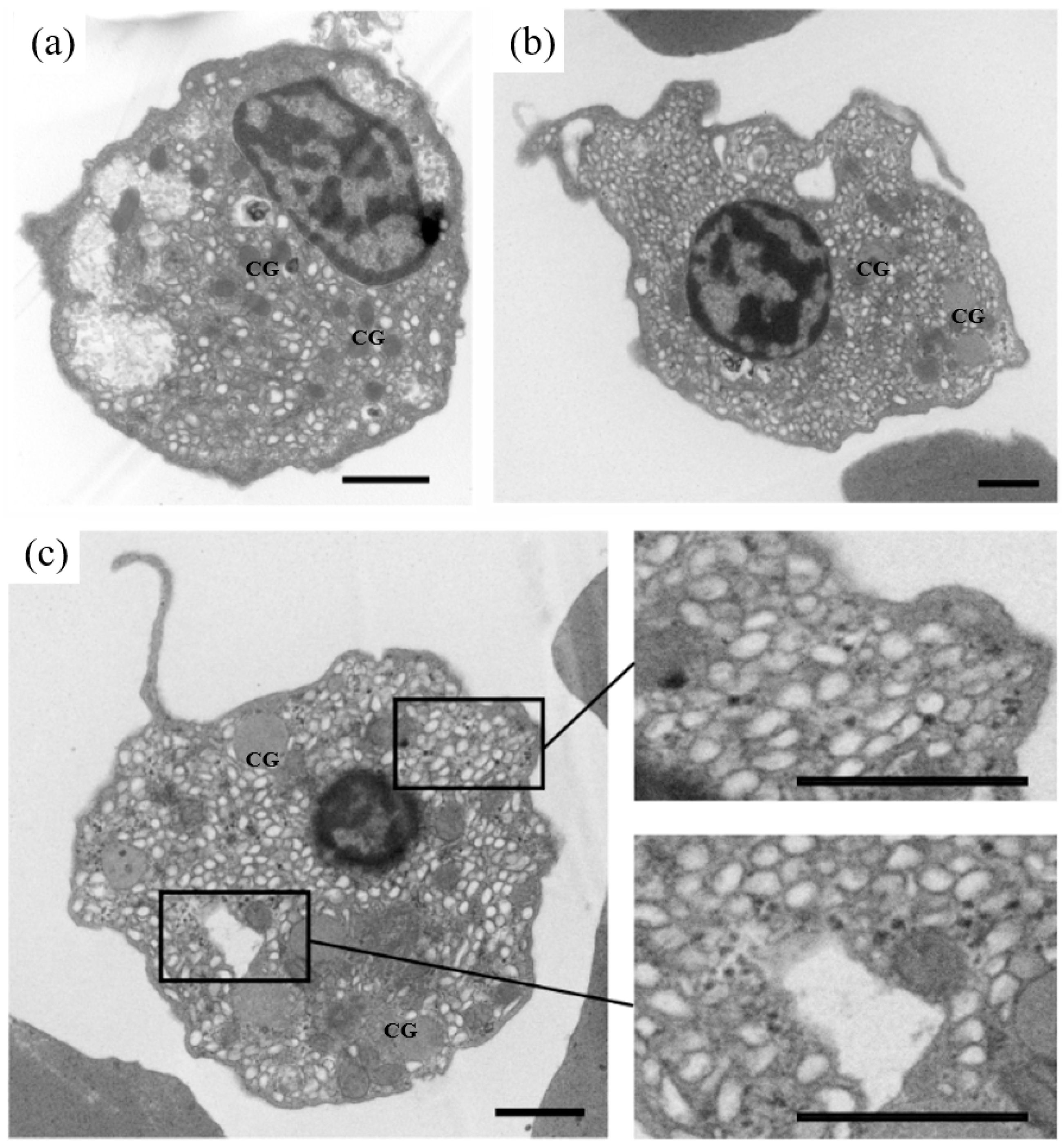
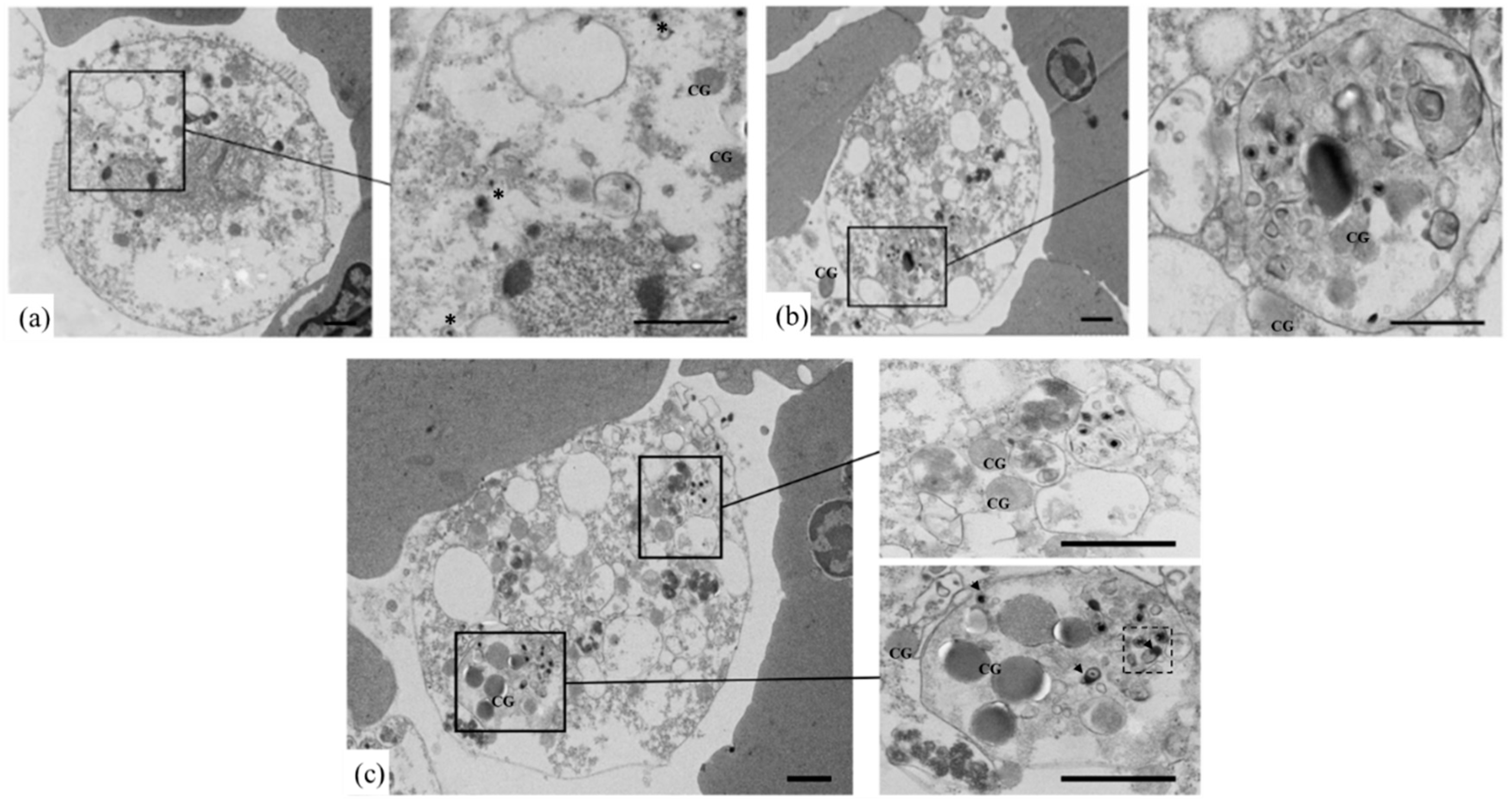
3.3. Identification of OsHV-1 Infected Hemocytes by TEM
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Farley, C.A.; Banfield, W.G., Jr.; Kasnic, G.; Foster, W.S. Oyster herpes-type virus. Science 1972, 178, 759–760. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, J.L.; Comps, M. Herpes-like virus infecting Pacific-oyster larvae, Crassostrea gigas. Bull. Eur. Assoc. Fish Pathol. 1992, 12, 11–13. [Google Scholar]
- Comps, M.; Cochennec, N. A Herpes-like Virus from the European Oyster Ostrea edulis L. J. Invertebr. Pathol. 1993, 62, 201–203. [Google Scholar] [CrossRef]
- Renault, T.; Lipart, C.; Arzul, I. A herpes-like virus infecting Crassostrea gigas and Ruditapes philippinarum larvae in France. J. Fish Dis. 2010, 24, 369–376. [Google Scholar] [CrossRef]
- Davison, A.J.; Trus, B.L.; Cheng, N.; Steven, A.C.; Watson, M.S.; Cunningham, C.; Le, D.R.; Renault, T. A novel class of herpesvirus with bivalve hosts. J. Gen. Virol. 2005, 86 Pt 1, 41–53. [Google Scholar] [CrossRef]
- Bai, C.; Gao, W.; Wang, C.; Yu, T.; Zhang, T.; Qiu, Z.; Wang, Q.; Huang, J. Identification and characterization of ostreid herpesvirus 1 associated with massive mortalities of Scapharca broughtonii broodstocks in China. Dis. Aquat. Organ. 2016, 118, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Ugalde, S.C.; Preston, J.; Ogier, E.; Crawford, C. Analysis of farm management strategies following herpesvirus (OsHV-1) disease outbreaks in Pacific oysters in Tasmania, Australia. Aquaculture 2018, 495, 179–186. [Google Scholar] [CrossRef]
- Carrasco, N.; Gairin, I.; Pérez, J.; Andree, K.B.; Roque, A.; Fernándeztejedor, M.; Rodgers, C.J.; Aguilera, C.; Furones, M.D. A Production Calendar Based on Water Temperature, Spat Size, and Husbandry Practices Reduce OsHV-1 μvar Impact on Cultured Pacific Oyster Crassostrea gigas in the Ebro Delta (Catalonia), Mediterranean Coast of Spain. Front. Physiol. 2017, 8, 125. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.Y.; Park, J.J.; Yu, H.J.; Hur, Y.B.; Arzul, I.; Couraleau, Y.; Park, M.A. Ostreid herpesvirus 1 infection in farmed Pacific oyster larvae Crassostrea gigas (Thunberg) in Korea. J. Fish Dis. 2013, 36, 969–972. [Google Scholar]
- Arzul, I.; Nicolas, J.L.; Davison, A.J.; Renault, T. French scallops: A new host for ostreid herpesvirus-1. Virology 2001, 290, 342–349. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Jouaux, A.; Ford, S.E.; Lelong, C.; Sourdaine, P.; Mathieu, M.; Guo, X. Transcriptome analysis reveals strong and complex antiviral response in a mollusc. Fish Shellfish Immunol. 2015, 46, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Green, T.J.; Speck, P. Antiviral Defense and Innate Immune Memory in the Oyster. Viruses 2018, 10, 133. [Google Scholar] [CrossRef] [PubMed]
- Pierrick, M.; Kevin, M.; Amélie, S.; Delphine, T.; Marie-Agnès, T.; Rubinsztein, D.C.; Tristan, R. Autophagy plays an important role in protecting Pacific oysters from OsHV-1 and Vibrio aestuarianus infections. Autophagy 2015, 11, 516–526. [Google Scholar]
- Green, T.J.; Rolland, J.-L.; Vergnes, A.; Raftos, D.; Montagnani, C. OsHV-1 countermeasures to the Pacific oyster’s anti-viral response. Fish Shellfish Immunol. 2015, 47, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Segarra, A.; Faury, N.; Pépin, J.-F.; Renault, T. Transcriptomic study of 39 ostreid herpesvirus 1 genes during an experimental infection. J. Invertebr. Pathol. 2014, 119, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhou, Z.; Jiang, Q.; Wang, L.; Yi, Q.; Qiu, L.; Song, L. The neuroendocrine immunomodulatory axis-like pathway mediated by circulating haemocytes in pacific oyster Crassostrea gigas. Open Biol. 2017, 7, 160289. [Google Scholar] [CrossRef] [PubMed]
- Adamo, S.A. Norepinephrine and octopamine: Linking stress and immune function across phyla. Invertebr. Surviv. J. 2008, 5, 12–19. [Google Scholar]
- Martenot, C.; Gervais, O.; Chollet, B.; Houssin, M.; Renault, T. Haemocytes collected from experimentally infected Pacific oysters, Crassostrea gigas: Detection of ostreid herpesvirus 1 DNA, RNA, and proteins in relation with inhibition of apoptosis. PLoS ONE 2017, 12, e0177448. [Google Scholar] [CrossRef] [PubMed]
- Friedman, C.S.; Estes, R.M.; Stokes, N.A.; Burge, C.A.; Hargove, J.S.; Barber, B.J.; Elston, R.A.; Burreson, E.M.; Reece, K.S. Herpes virus in juvenile Pacific oysters Crassostrea gigas from Tomales Bay, California, coincides with summer mortality episodes. Dis. Aquat. Organ. 2005, 63, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.T.; Liu, W.; Seah, J.N.; Lam, C.S.; Xiang, J.H.; Korzh, V.; Kwang, J. White spot syndrome virus (WSSV) infects specific hemocytes of the shrimp Penaeus merguiensis. Dis. Aquat. Organ. 2002, 52, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Jia, Z.; Zhang, T.; Wang, L.; Qiu, L.; Sun, J.; Song, L. Functional characterisation of phagocytes in the Pacific oyster Crassostrea gigas. PeerJ 2016, 4, e2590. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Bai, C.; Wang, C.; Song, X.; Huang, J. Complete genome sequence of Ostreid herpesvirus-1 associated with mortalities of Scapharca broughtonii broodstocks. Virol. J. 2015, 12, 110. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Yang, A.; Liu, Z.; Wu, B.; Sun, X.; Lv, Z.; Tian, J.T.; Du, M. Changes in hemolymph characteristics of ark shell Scapharaca broughtonii dealt with Vibrio anguillarum challenge in vivo and various of anticoagulants in vitro. Fish Shellfish Immunol. 2017, 61, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Schikorski, D.; Renault, T.; Saulnier, D.; Faury, N.; Moreau, P.; Pépin, J.F. Experimental infection of Pacific oyster Crassostrea gigas spat by ostreid herpesvirus 1: Demonstration of oyster spat susceptibility. Vet. Res. 2011, 42, 27. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.M.; Wang, Q.C.; Morga, B.; Shi, J.; Wang, C.M. Experimental infection of adult Scapharca broughtonii with Ostreid herpesvirus SB strain. J. Invertebr. Pathol. 2016, 143, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Martenot, C.; Oden, E.; Travaillé, E.; Malas, J.P.; Houssin, M. Comparison of two real-time PCR methods for detection of ostreid herpesvirus 1 in the Pacific oyster Crassostrea gigas. J. Virol. Methods 2010, 170, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, M.; Wang, L.; Chen, H.; Liu, Z.; Jia, Z.; Qiu, L.; Song, L. The granulocytes are the main immunocompetent hemocytes in Crassostrea gigas. Dev. Comp. Immunol. 2016, 67, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Allam, B.; Ashtonalcox, K.A.; Ford, S.E. Flow cytometric comparison of haemocytes from three species of bivalve molluscs. Fish Shellfish Immunol. 2002, 13, 141–158. [Google Scholar] [CrossRef] [PubMed]
- Gougeon, M.L. Apoptosis as an HIV strategy to escape immune attack. Nat. Rev. Immunol. 2003, 3, 392–404. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xin, L.; Li, C.; Bai, C.; Wang, C. Ostreid Herpesvirus-1 Infects Specific Hemocytes in Ark Clam, Scapharca broughtonii. Viruses 2018, 10, 529. https://doi.org/10.3390/v10100529
Xin L, Li C, Bai C, Wang C. Ostreid Herpesvirus-1 Infects Specific Hemocytes in Ark Clam, Scapharca broughtonii. Viruses. 2018; 10(10):529. https://doi.org/10.3390/v10100529
Chicago/Turabian StyleXin, Lusheng, Chen Li, Changming Bai, and Chongming Wang. 2018. "Ostreid Herpesvirus-1 Infects Specific Hemocytes in Ark Clam, Scapharca broughtonii" Viruses 10, no. 10: 529. https://doi.org/10.3390/v10100529
APA StyleXin, L., Li, C., Bai, C., & Wang, C. (2018). Ostreid Herpesvirus-1 Infects Specific Hemocytes in Ark Clam, Scapharca broughtonii. Viruses, 10(10), 529. https://doi.org/10.3390/v10100529