Effects of Management Practices and Topography on Ectomycorrhizal Fungi of Maritime Pine during Seedling Recruitment

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
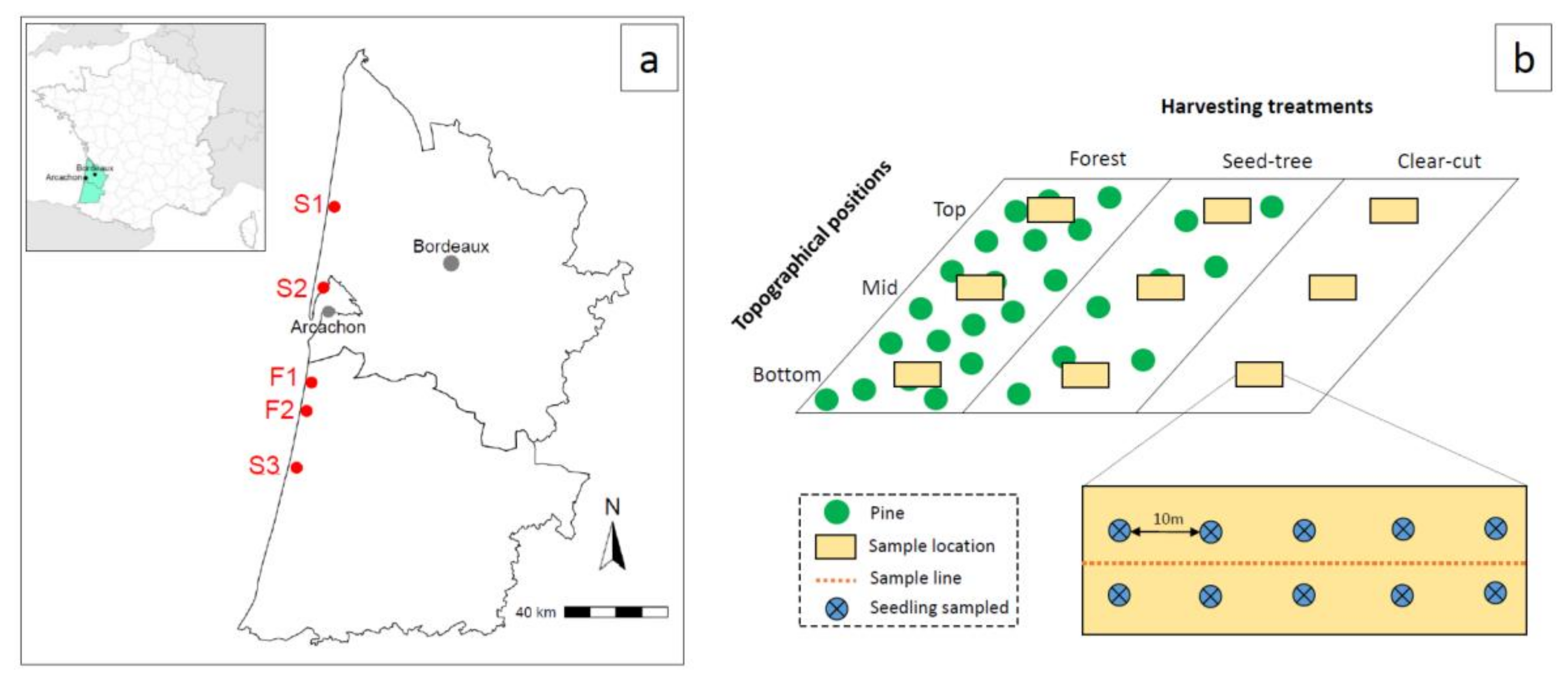
2.1. Study Area Description
2.2. Tree Harvesting and Stand Regeneration Management Methods
2.3. Sampling of Seedlings
2.4. Assessment of Seedling Dimensions and Ectomycorrhizal Status
2.5. Needle Nutrient Concentrations
2.6. Data Treatments and Statistics
2.6.1. ECM Status
2.6.2. Seedling Growth and Nutrient Concentrations
2.6.3. Relationship between ECM and Seedling Response after Filtering out Effects of Harvesting Treatments and Topographical Positions
3. Results
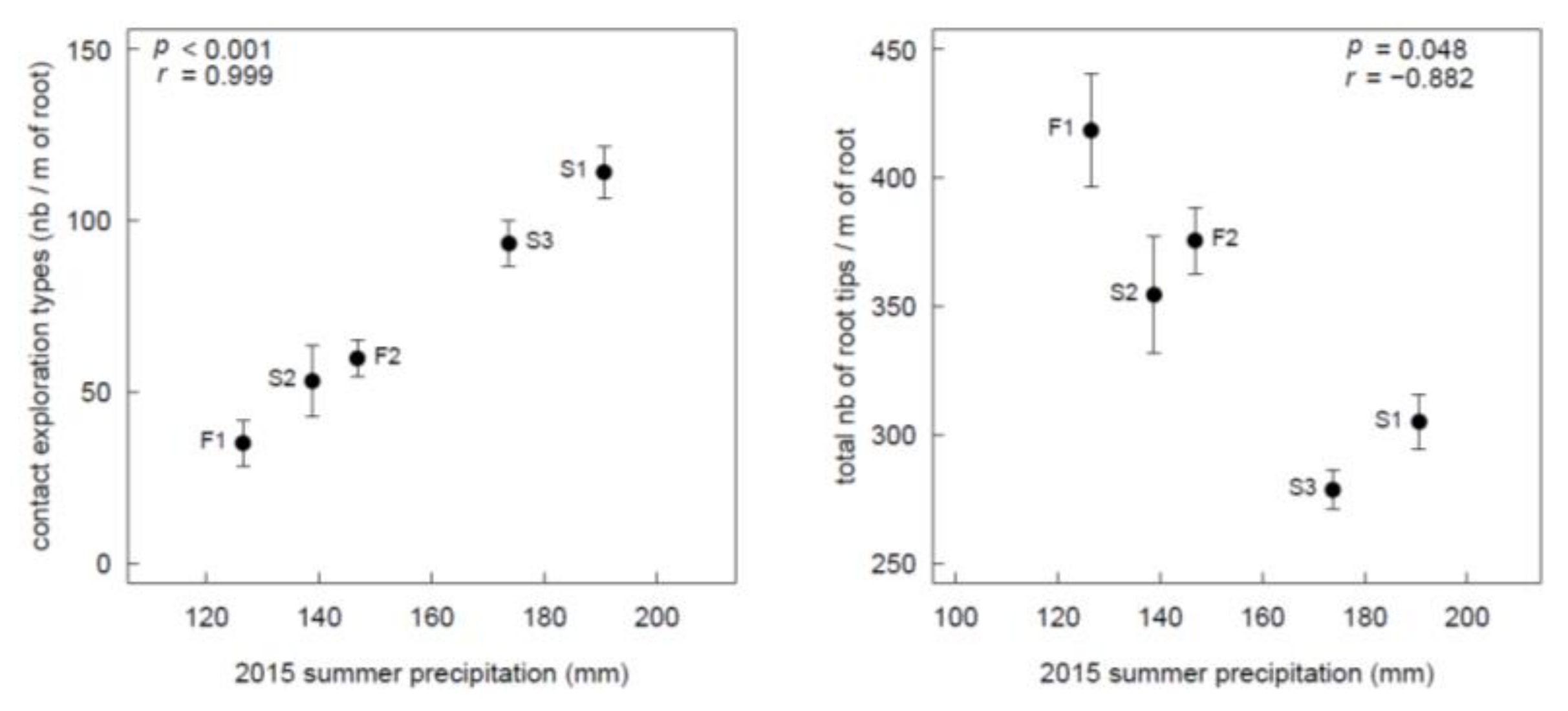
3.1. Sites
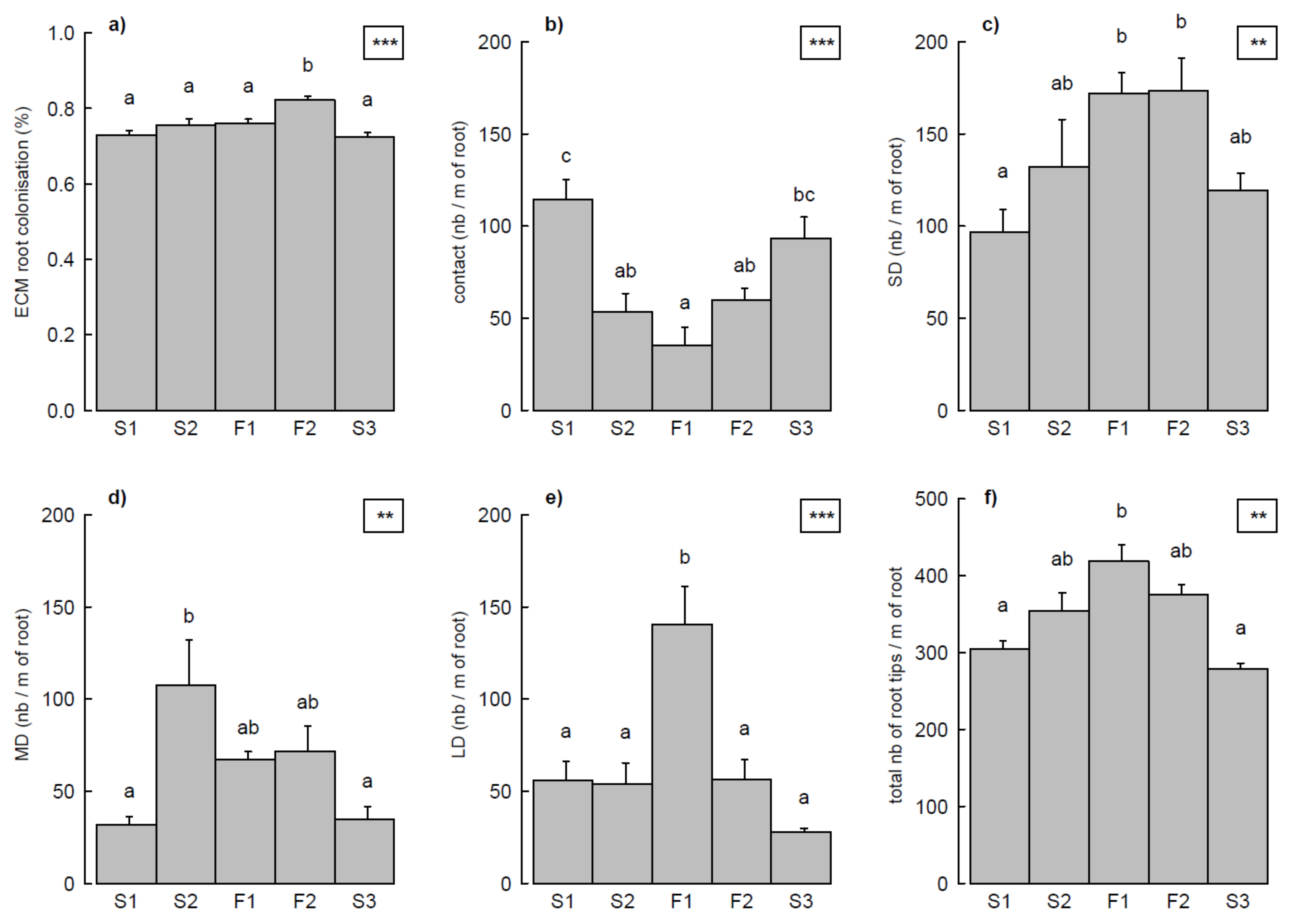
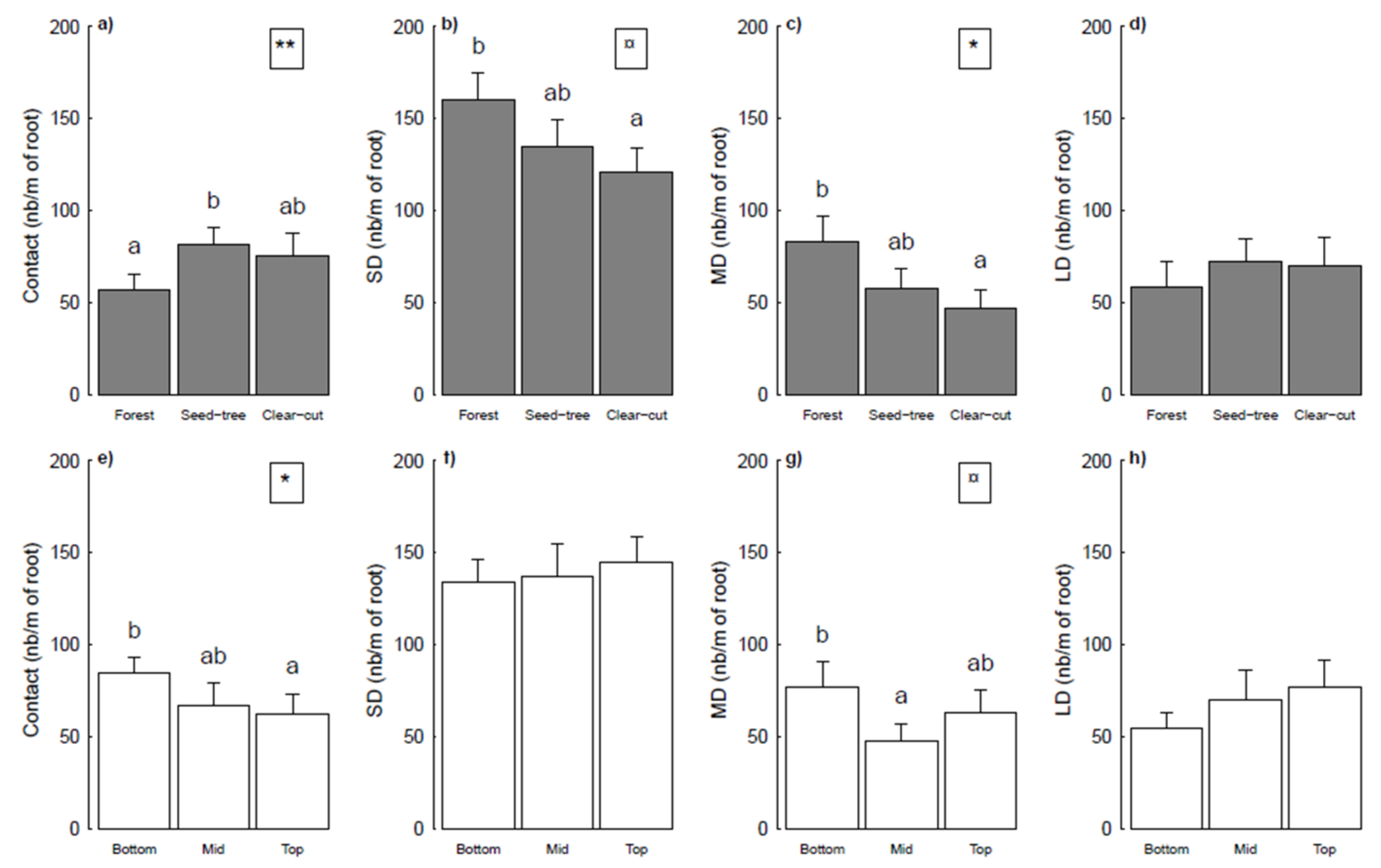
3.2. ECM Status Related to Harvesting Treatments and Topographical Positions
3.3. Seedling Growth and Needle Nutrient Concentrations
3.4. Effects of ECM on Seedling Root Properties and Nutrient Concentrations
4. Discussion
4.1. Role of Abiotic Factors in ECM Composition
4.2. Impact of Forest Management on ECM Composition
4.3. Impact of Forest Management on Seedling Characteristics
4.4. Effects of ECM on Seedling Root Development and Nutritive Status
4.5. Can ECM Explain Failures of Forest Regeneration?
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- IFN Inventaire Forestier National. Available online: https://inventaire-forestier.ign.fr/ (accessed on 18 December 2017).
- Sardin, T. Guide des Sylvicultures Forêts Littorales Atlantiques Dunaires, ONF ed.; Office National des Forêts: Paris, France, 2009; p. 175. ISBN 9782842073374. [Google Scholar]
- Ouallet, P. Quels Peuvent-être les Facteurs Écologiques Responsables des Échecs de Régénération Naturelle du pin Maritime sur les Dunes Littorales des Forêts Domaniales de Biscarrosse et de Sainte-Eulalie? Master’s Thesis, Bordeaux Sciences Agro et ONF, Gradignan, France, 2012. [Google Scholar]
- Clark, J.S.; Macklin, E.; Wood, L. Stages and Spatial Scales of Recruitment Limitation in Southern Apalachians forests. Ecol. Monogr. 1998, 68, 213–235. [Google Scholar] [CrossRef]
- Kitajima, K.; Fenner, M. Ecology of seedling regeneration. In Seeds: The Ecology of Regeneration in Plant Communities; CABI: Wallingford, UK, 2000; pp. 331–359. ISBN 0851994326. [Google Scholar]
- Harvey, A.E.; Jurgensen, M.F.; Larsen, M.J.; Graham, R.T. Relationships among soil microsite, ectomycorrhizae, and natural conifer regeneration of old-growth forests in western Montana. Can. J. For. Res. 1987, 17, 58–62. [Google Scholar] [CrossRef]
- Harmer, R. The effect of plant competition and simulated summer browsing by deer on tree regeneration. J. Appl. Ecol. 2001, 38, 1094–1103. [Google Scholar] [CrossRef]
- Rodríguez-García, E.; Gratzer, G.; Bravo, F. Climatic variability and other site factor influences on natural regeneration of Pinus pinaster Ait. in Mediterranean forests. Ann. For. Sci. 2011, 68, 811–823. [Google Scholar] [CrossRef]
- Perry, D.A.; Molina, R.; Amaranthus, M.P. Mycorrhizae, mycorrhizospheres, and reforestation: Current knowledge and research needs. Can. J. For. Res. 1987, 17, 929–940. [Google Scholar] [CrossRef]
- Miller, S.L.; McClean, T.M.; Stanton, N.L.; Williams, S.E. Mycorrhization, physiognomy, and first-year survivability of conifer seedlings following natural fire in Grand Teton National Park. Can. J. For. Res. 1998, 28, 115–122. [Google Scholar] [CrossRef]
- Barker, J.S.; Simard, S.W.; Jones, M.D.; Durall, D.M. Ectomycorrhizal fungal community assembly on regenerating Douglas-fir after wildfire and clearcut harvesting. Oecologia 2013, 172, 1179–1189. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, J.V.D.; Sylvia, D.M.; Fox, A.J. Contribution of ectomycorrhiza to the potential nutrient-absorbing surface of pine. New Phytol. 1994, 128, 639–644. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Cambridge, MA, USA, 2008; ISBN 9780123705266. [Google Scholar]
- Teste, F.P.; Simard, S.W.; Durall, D.M.; Guy, R.D.; Jones, M.D.; Schoonmaker, A.L. Access to mycorrhizal networks and roots of trees: Importance for seedling survival and resource transfer. Ecology 2009, 90, 2808–2822. [Google Scholar] [CrossRef] [PubMed]
- Perrin, R.; Garbaye, J. Influence of ectomycorrhizae on infectivity of Pythium-infested soils and substrates. Plant Soil 1983, 71, 345–351. [Google Scholar] [CrossRef]
- Van Tichelen, K.K.; Colpaert, J.V.; Vangronsveld, J. Ectomycorrhizal protection of Pinus sylvestris against copper toxicity. New Phytol. 2001, 150, 203–213. [Google Scholar] [CrossRef]
- Lehto, T.; Zwiazek, J.J. Ectomycorrhizas and water relations of trees: A review. Mycorrhiza 2011, 21, 71–90. [Google Scholar] [CrossRef] [PubMed]
- Mousain, D. Effets de deux facteurs edaphiques (teneur en phosphore et qualite de la matiere organique des sols) sur l’etablissement de la symbiose ectomycorhizienne du pin maritime (Pinus pinaster Soland. in Ait.). Rev. Ecol. Biol. Sol 1975, 12, 239–259. [Google Scholar]
- Read, D.J.; Leake, J.R.; Perez-Moreno, J. Mycorrhizal fungi as drivers of ecosystem processes in heathland and boreal forest biomes. Can. J. Bot. 2004, 82, 1243–1263. [Google Scholar] [CrossRef]
- Agerer, R. Exploration types of ectomycorrhizae: A proposal to classify ectomycorrhizal mycelial systems according to their patterns of differentiation and putative ecological importance. Mycorrhiza 2001. [Google Scholar] [CrossRef]
- Lilleskov, E.A.; Hobbie, E.A.; Horton, T.R. Conservation of ectomycorrhizal fungi: Exploring the linkages between functional and taxonomic responses to anthropogenic N deposition. Fungal Ecol. 2011, 4, 174–183. [Google Scholar] [CrossRef]
- Bakker, M.R.; Augusto, L.; Achat, D.L. Fine root distribution of trees and understory in mature stands of maritime pine (Pinus pinaster) on dry and humid sites. Plant Soil 2006, 286, 37–51. [Google Scholar] [CrossRef]
- Hobbie, E.A.; Agerer, R. Nitrogen isotopes in ectomycorrhizal sporocarps correspond to belowground exploration types. Plant Soil 2010, 327, 71–83. [Google Scholar] [CrossRef]
- Slankis, V. Soil Factors Influencing Formation of Mycorrhizae. Annu. Rev. Phytopathol. 1974, 12, 437–457. [Google Scholar] [CrossRef]
- Miyamoto, Y.; Sakai, A.; Hattori, M.; Nara, K. Strong effect of climate on ectomycorrhizal fungal composition: Evidence from range overlap between two mountains. ISME J. 2015, 9, 1870–1879. [Google Scholar] [CrossRef] [PubMed]
- Cox, F.; Barsoum, N.; Lilleskov, E.A.; Bidartondo, M.I. Nitrogen availability is a primary determinant of conifer mycorrhizas across complex environmental gradients. Ecol. Lett. 2010, 13, 1103–1113. [Google Scholar] [CrossRef] [PubMed]
- Kilpeläinen, J.; Barbero-López, A.; Vestberg, M.; Heiskanen, J.; Lehto, T. Does severe soil drought have after-effects on arbuscular and ectomycorrhizal root colonisation and plant nutrition? Plant Soil 2017. [Google Scholar] [CrossRef]
- Fernandez, C.W.; Nguyen, N.H.; Stefanski, A.; Han, Y.; Hobbie, S.E.; Montgomery, R.A.; Reich, P.B.; Kennedy, P.G. Ectomycorrhizal fungal response to warming is linked to poor host performance at the boreal-temperate ecotone. Glob. Chang. Biol. 2017, 23, 1598–1609. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, S.; Woodward, S.; Alexander, I.J.; Taylor, A.F.S. Regional scale gradients of climate and nitrogen deposition drive variation in ectomycorrhizal fungal communities associated with native Scots pine. Glob. Chang. Biol. 2013, 19, 1688–1696. [Google Scholar] [CrossRef] [PubMed]
- De Witte, L.C.; Rosenstock, N.P.; van der Linde, S.; Braun, S. Nitrogen deposition changes ectomycorrhizal communities in Swiss beech forests. Sci. Total Environ. 2017, 605–606, 1083–1096. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.E.; Jurgensen, M.F.; Larsen, M.J.; Schlieter, J.A. Distribution of Active Ectomycorrhizal Short Roots of the Inland Northwest: Effects of Site and Disturbance; INT-374; Intermountain Research Station: Ogden, UT, USA, 1986; p. 8. [Google Scholar]
- Jones, M.D.; Durall, D.M.; Cairney, J.W.G. Ectomycorrhizal fungal communities in young forest stands regenerating after clearcut logging. New Phytol. 2003, 157, 399–422. [Google Scholar] [CrossRef]
- Lazaruk, L.W.; Macdonald, S.E.; Kernaghan, G. The effect of mechanical site preparation on ectomycorrhizae of planted white spruce seedlings in conifer-dominated boreal mixedwood forest. Can. J. For. Res. 2008, 38, 2072–2079. [Google Scholar] [CrossRef]
- Page-Dumroese, D.S.; Harvey, A.E.; Jurgensen, M.F.; Amaranthus, M.P. Impacts of soil compaction and tree stump removal on soil properties and outplanted seedlings in northern Idaho, USA. Can. J. Soil Sci. 1998, 78, 29–34. [Google Scholar] [CrossRef]
- Kranabetter, J.M.; Haeussler, S.; Wood, C. Vulnerability of boreal indicators (ground-dwelling beetles, understory plants and ectomycorrhizal fungi) to severe forest soil disturbance. For. Ecol. Manag. 2017, 402, 213–222. [Google Scholar] [CrossRef]
- Lazaruk, L.W.; Kernaghan, G.; Macdonald, S.E.; Khasa, D. Effects of partial cutting on the ectomycorrhizae of Picea glauca forests in northwestern Alberta. Can. J. For. Res. 2005, 35, 1442–1454. [Google Scholar] [CrossRef]
- Jones, M.D.; Twieg, B.D.; Durall, D.M.; Berch, S.M. Location relative to a retention patch affects the ECM fungal community more than patch size in the first season after timber harvesting on Vancouver Island, British Columbia. For. Ecol. Manag. 2008, 255, 1342–1352. [Google Scholar] [CrossRef]
- Rodríguez-García, E.; Bravo, F.; Spies, T.A. Effects of overstorey canopy, plant–plant interactions and soil properties on Mediterranean maritime pine seedling dynamics. For. Ecol. Manag. 2011, 262, 244–251. [Google Scholar] [CrossRef]
- Bingham, M.A.; Simard, S. Ectomycorrhizal Networks of Pseudotsuga menziesii var. glauca Trees Facilitate Establishment of Conspecific Seedlings under Drought. Ecosystems 2012, 15, 188–199. [Google Scholar] [CrossRef]
- Warren, J.M.; Brooks, J.R.; Meinzer, F.C.; Eberhart, J.L. Hydraulic redistribution of water from Pinus ponderosa trees to seedlings: Evidence for an ectomycorrhizal pathway. New Phytol. 2008, 178, 382–394. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, L.; Dahlberg, A.; Nilsson, M.C.; Kårén, O.; Zackrisson, O. Continuity of ectomycorrhizal fungi in self-regenerating boreal Pinus sylvestris forests studied by comparing mycobiont diversity on seedlings and mature trees. New Phytol. 1999, 142, 151–162. [Google Scholar] [CrossRef]
- Dahlberg, A.; Schimmel, J.; Taylor, A.F.S.; Johannesson, H. Post-fire legacy of ectomycorrhizal fungal communities in the Swedish boreal forest in relation to fire severity and logging intensity. Biol. Conserv. 2001, 100, 151–161. [Google Scholar] [CrossRef]
- Pitaud, G. L’hydrogéologie et la Végétation dans les Dunes du Littoral Aquitain. Ph.D. Thesis, University Bordeaux, Nouvelle-Aquitaine, France, 1967. [Google Scholar]
- Augusto, L.; Bakker, M.R.; Morel, C.; Meredieu, C.; Trichet, P.; Badeau, V.; Arrouays, D.; Plassard, C.; Achat, D.L.; Gallet-Budynek, A.; et al. Is “grey literature” a reliable source of data to characterize soils at the scale of a region? A case study in a maritime pine forest in southwestern France. Eur. J. Soil Sci. 2010, 61, 807–822. [Google Scholar] [CrossRef]
- Porté, A.; Bosc, A.; Champion, I.; Loustau, D. Estimating the foliage area of Maritime pine (Pinus pinaster Aït.) branches and crowns with application to modelling the foliage area distribution in the crown. Ann. For. Sci. 2000, 57, 73–86. [Google Scholar] [CrossRef]
- Berbigier, P.; Bonnefond, J. Measurement and modelling of radiation transmission within a stand of maritime pine (Pinus pinaster Ait). Ann. Sci. For. 1995, 52, 23–42. [Google Scholar] [CrossRef]
- Tennant, D. A test of a modified line intersect method of estimating root length. J. Ecol. 1975, 63, 995–1001. [Google Scholar] [CrossRef]
- Van den Burg, J. Foliar Analysis for Determination of Tree Nutrient Status: A Compilation of Literature Data; Rijksinstituut voor Onderzoek in de Bos-en Landschapsbouw de Dorschkamp: Wageningen, The Netherlands, 1985. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 3 July 2017).
- Warton, D.I.; Hui, F.K.C. The arcsine is asinine: The analysis of proportions in ecology. Ecology 2011, 92, 3–10. [Google Scholar] [CrossRef] [PubMed]
- McConnaughay, K.D.M.; Coleman, J.S. Biomass Allocation in Plants: Ontogeny or Optimality? A Test along Three Resource Gradients. Ecology 1999, 80, 2581–2593. [Google Scholar] [CrossRef]
- Achat, D.L.; Deleuze, C.; Landmann, G.; Pousse, N.; Ranger, J.; Augusto, L. Quantifying consequences of removing harvesting residues on forest soils and tree growth—A meta-analysis. For. Ecol. Manag. 2015, 348, 124–141. [Google Scholar] [CrossRef]
- Augusto, L.; Ranger, J.; Ponette, Q.; Rapp, M. Relationships between forest tree species, stand production and stand nutrient amount. Ann. For. Sci. 2000, 57, 313–324. [Google Scholar] [CrossRef]
- Ritson, P.; Sochacki, S. Measurement and prediction of biomass and carbon content of Pinus pinaster trees in farm forestry plantations, south-western Australia. For. Ecol. Manag. 2003, 175, 103–117. [Google Scholar] [CrossRef]
- Pena, R.; Simon, J.; Rennenberg, H.; Polle, A. Ectomycorrhiza affect architecture and nitrogen partitioning of beech (Fagus sylvatica L.) seedlings under shade and drought. Environ. Exp. Bot. 2013, 87, 207–217. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Toots, M.; Diédhiou, A.G.; Henkel, T.W.; Kjøller, R.; Morris, M.H.; Nara, K.; Nouhra, E.; Peay, K.G.; et al. Towards global patterns in the diversity and community structure of ectomycorrhizal fungi. Mol. Ecol. 2012, 21, 4160–4170. [Google Scholar] [CrossRef] [PubMed]
- Bahram, M.; Põlme, S.; Kõljalg, U.; Zarre, S.; Tedersoo, L. Regional and local patterns of ectomycorrhizal fungal diversity and community structure along an altitudinal gradient in the Hyrcanian forests of northern Iran. New Phytol. 2012, 193, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Guttenberger, M.; Kottke, I.; Hampp, R. The effect of drought on mycorrhizas of beech (Fagus sylvatica L.): Changes in community structure, and the content of carbohydrates and nitrogen storage bodies of the fungi. Mycorrhiza 2002, 12, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Sewerniak, P.; Jankowski, M. Topographically-controlled site conditions drive vegetation pattern on inland dunes in Poland. Acta Oecol. 2017, 82, 52–60. [Google Scholar] [CrossRef]
- Bahr, A.; Ellström, M.; Akselsson, C.; Ekblad, A.; Mikusinska, A.; Wallander, H. Growth of ectomycorrhizal fungal mycelium along a Norway spruce forest nitrogen deposition gradient and its effect on nitrogen leakage. Soil Biol. Biochem. 2013, 59, 38–48. [Google Scholar] [CrossRef]
- Bakker, M.; Garbaye, J.; Nys, C. Effect of liming on the ectomycorrhizal status of oak. For. Ecol. Manag. 2000, 126, 121–131. [Google Scholar] [CrossRef]
- Tedersoo, L.; Smith, M.E. Lineages of ectomycorrhizal fungi revisited: Foraging strategies and novel lineages revealed by sequences from belowground. Fungal Biol. Rev. 2013, 27, 83–99. [Google Scholar] [CrossRef]
- Cline, E.T.; Ammirati, J.F.; Edmonds, R.L. Does proximity to mature trees influence ectomycorrhizal fungus communities of Douglas-fir seedlings? New Phytol. 2005, 166, 993–1009. [Google Scholar] [CrossRef] [PubMed]
- Peay, K.G.; Schubert, M.G.; Nguyen, N.H.; Bruns, T.D. Measuring ectomycorrhizal fungal dispersal: Macroecological patterns driven by microscopic propagules. Mol. Ecol. 2012, 21, 4122–4136. [Google Scholar] [CrossRef] [PubMed]
- Robakowski, P.; Wyka, T.; Samardakiewicz, S.; Kierzkowski, D. Growth, photosynthesis, and needle structure of silver fir (Abies alba Mill.) seedlings under different canopies. For. Ecol. Manag. 2004, 201, 211–227. [Google Scholar] [CrossRef]
- Chen, H.Y.; Klinka, K. Survival, growth, and allometry of planted Larix occidentalis seedlings in relation to light availability. For. Ecol. Manag. 1998, 106, 169–179. [Google Scholar] [CrossRef]
- Elliott, K.J.; White, A.S. Effects of light, nitrogen, and phosphorus on red pine seedling growth and nutrient use efficiency. For. Sci. 1994, 40, 47–58. [Google Scholar]
- Rodríguez-García, E.; Bravo, F. Plasticity in Pinus pinaster populations of diverse origins: Comparative seedling responses to light and Nitrogen availability. For. Ecol. Manag. 2013, 307, 196–205. [Google Scholar] [CrossRef]
- Ruano, I.; Pando, V.; Bravo, F. How do light and water influence Pinus pinaster Ait. germination and early seedling development? For. Ecol. Manag. 2009, 258, 2647–2653. [Google Scholar] [CrossRef]
- Ingleby, K.; Munro, R.C.; Noor, M.; Mason, P.A.; Clearwater, M.J. Ectomycorrhizal populations and growth of Shorea parvifolia (Dipterocarpaceae) seedlings regenerating under three different forest canopies following logging. For. Ecol. Manag. 1998, 111, 171–179. [Google Scholar] [CrossRef]
- Zavala, M.A.; Espelta, J.M.; Caspersen, J.; Retana, J. Interspecific differences in sapling performance with respect to light and aridity gradients in Mediterranean pine–oak forests: Implications for species coexistence. Can. J. For. Res. 2011, 41, 1432–1444. [Google Scholar] [CrossRef]
- Wang, G.G.; Qian, H.; Klinka, K. Growth of Thuja plicata seedlings along a light gradient. Can. J. Bot. 1994, 72, 1749–1757. [Google Scholar] [CrossRef]
- Markesteijn, L.; Poorter, L. Seedling root morphology and biomass allocation of 62 tropical tree species in relation to drought- and shade-tolerance. J. Ecol. 2009, 97, 311–325. [Google Scholar] [CrossRef]
- Brunner, I.; Herzog, C.; Dawes, M.A.; Arend, M.; Sperisen, C. How tree roots respond to drought. Front. Plant Sci. 2015, 6, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Alexander, I.J. The Picea sitchensis + Lactarius rufus mycorrhizal association and its effects on seedling growth and development. Trans. Br. Mycol. Soc. 1981, 76, 417–423. [Google Scholar] [CrossRef]
- Ostonen, I.; Tedersoo, L.; Suvi, T.; Lõhmus, K. Does a fungal species drive ectomycorrhizal root traits in Alnus spp.? Can. J. For. Res. 2009, 39, 1787–1796. [Google Scholar] [CrossRef]
- Nara, K. Ectomycorrhizal networks and seedling establishment during early primary succession. New Phytol. 2006, 169, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Pigott, C.D. Survival of mycorrhiza formed by Cenococcum geophilum Fr. in dry soils. New Phytol. 1982, 92, 513–517. [Google Scholar] [CrossRef]
- Kreuzwieser, J.; Gessler, A. Global climate change and tree nutrition: Influence of water availability. Tree Physiol. 2010, 30, 1221–1234. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S1 | S2 | F1 | F2 | S3 | |
|---|---|---|---|---|---|
| Regeneration status ⌂ | |||||
| Survival (%) | 59.8 | 31.0 | 15.8 | 25.7 | 66.2 |
| Seedlings/ha | 3062 | 3875 | 1000 | 1188 | 8438 |
| Location | |||||
| Forest | Lacanau | Lège-Cap-Ferret | Biscarrosse | Sainte-Eulalie-en-Born | Saint-Julien-en-Born |
| Latitude | 45°1’43.2” N | 44°44’9.4” N | 44°24’37.1” N | 44°18’14” N | 44°6’8” N |
| Longitude | 1°10’44.1” W | 1°12’35.6” W | 1°14’3.4” W | 1°15’8.2” W | 1°17’27” W |
| Meteo | |||||
| Annual P (mm) | 709 (879) | 608 (840) | 659 (894) | 718 (936) | 880 (1007) |
| Summer P (mm) | 191 (155) | 139 (141) | 127 (151) | 147 (157) | 174 (165) |
| Annual T (°C) | 13.8 (13.7) | 14.4 (14.1) | 14.4 (14.1) | 14.4 (14.1) | 14.2 (14.0) |
| Summer T (°C) | 19.5 (19.7) | 20.2 (20.3) | 20.2 (20.2) | 20.2 (20.2) | 19.9 (20.1) |
| Annual PET (mm) | 759 (756) | 780 (773) | 778 (770) | 778 (771) | 774 (769) |
| Soil | |||||
| pH (H2O) ± SE | 4.46 ± 0.14 a | 4.82 ± 0.21 c | 4.57 ± 0.13 ab | 4.68 ± 0.19 bc | 4.44 ± 0.14 a |
| OM (%) ± SE | 42.5 ± 13.7 d | 24.8 ± 13.9 ab | 28.2 ± 10.9 bc | 36.8 ± 15.7 cd | 14.3 ± 8.5 a |
| Harvesting Treatments | Topographical Positions | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Forest | Seed-Tree | Clear-Cut | Bottom | Mid | Top | |||||||
| Stem diameter (mm) | 1.55 (0.05) | a | 1.69 (0.06) | b | 1.92 (0.06) | c | 1.77 (0.07) | 1.71 (0.08) | 1.68 (0.06) | |||
| Height (mm) | 125.2 (4.7) | b | 109.4 (3.2) | a | 110.9 (2.7) | a | 115.6 (3.4) | ab | 120.2 (4.2) | b | 109.8 (4.2) | a |
| Shoot biomass (g) | 0.29 (0.02) | a | 0.43 (0.03) | b | 0.56 (0.05) | c | 0.44 (0.05) | 0.44 (0.05) | 0.41 (0.05) | |||
| Root biomass (g) | 0.12 (0.01) | a | 0.20 (0.02) | b | 0.20 (0.02) | b | 0.17 (0.02) | 0.17 (0.02) | 0.18 (0.02) | |||
| Seedling biomass (g) | 0.41 (0.03) | a | 0.63 (0.05) | b | 0.76 (0.06) | c | 0.60 (0.06) | 0.61 (0.07) | 0.59 (0.06) | |||
| Fine root length (cm) | 99.4 (13.5) | a | 187.7 (30.0) | b | 163.5 (23.7) | b | 148.1 (25.2) | 168.6 (26.8) | 133.9 (23.4) | |||
| Tap root length (cm) | 18.5 (0.71) | a | 21.7 (0.82) | b | 23.1 (0.56) | b | 21.1 (0.84) | 21.4 (0.80) | 20.8 (0.95) | |||
| Height:diameter (mm mm−1) ▪ | 82.2 (2.2) | c | 67.0 (2.3) | b | 59.8 (1.8) | a | 67.6 (2.6) | a | 74.0 (3.8) | b | 67.3 (3.0) | a |
| Root:shoot (g g−1) ▪ | 0.42 (0.03) | a | 0.50 (0.03) | b | 0.39 (0.02) | a | 0.42 (0.03) | 0.42 (0.03) | 0.47 (0.04) | |||
| Specific root length (m g−1) | 9.04 (0.86) | 9.76 (1.04) | 8.94 (0.92) | 9.68 (1.03) | b | 9.98 (0.73) | b | 8.08 (0.97) | a | |||
| N-needle (mg g−1) ▪ | 11.5 (0.3) ‡ | a | 18.8 (0.6) | b | 24.6 (0.6) | c | 18.3 (1.5) | 18.7 (1.4) | 17.9 (1.6) | |||
| P-needle (mg g−1) ▪ | 1.26 (0.05) ‡ | a | 2.36 (0.09) | b | 3.12 (0.18) | c | 2.20 (0.23) | 2.38 (0.24) | 2.16 (0.23) | |||
| K-needle (mg g−1) ▪ | 5.82 (0.31) † | a | 9.47 (0.34) | b | 10.12 (0.31) | b | 8.37 (0.60) | 8.90 (0.60) | 8.14 (0.57) | |||
| Ca-needle (mg g−1) ▪ | 1.93 (0.05) † | a | 2.65 (0.11) † | b | 2.43 (0.08) † | a | 2.32 (0.10) † | 2.41 (0.15) † | 2.27 (0.09) † | |||
| Mg-needle (mg g−1) | 1.58 (0.07) | 1.59 (0.09) | 1.65 (0.04) | 1.60 (0.07) | 1.57 (0.07) | 1.65 (0.07) | ||||||
| ECM Root Colonization (%) | Contact (nb/m of Root) | SD (nb/m of Root) | MD (nb/m of Root) | LD (nb/m of Root) | Total nb of Root Tips/m of Root | |
|---|---|---|---|---|---|---|
| Root:shoot (g g−1) | 0.05 ¤ | |||||
| Specific root length (m g−1) | 0.32 *** | −0.06 ¤ | −0.31 *** | −0.21 *** | −0.24 *** | |
| N-needle (mg g−1) | 0.17 ** | 0.07 * | ||||
| P-needle (mg g−1) | ||||||
| K-needle (mg g−1) | 0.17 ** | 0.21 ** | ||||
| Ca-needle (mg g−1) | 0.12 * | −0.11 * | ||||
| Mg-needle (mg g−1) | 0.07 * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guignabert, A.; Delerue, F.; Gonzalez, M.; Augusto, L.; Bakker, M.R. Effects of Management Practices and Topography on Ectomycorrhizal Fungi of Maritime Pine during Seedling Recruitment. Forests 2018, 9, 245. https://doi.org/10.3390/f9050245
Guignabert A, Delerue F, Gonzalez M, Augusto L, Bakker MR. Effects of Management Practices and Topography on Ectomycorrhizal Fungi of Maritime Pine during Seedling Recruitment. Forests. 2018; 9(5):245. https://doi.org/10.3390/f9050245
Chicago/Turabian StyleGuignabert, Arthur, Florian Delerue, Maya Gonzalez, Laurent Augusto, and Mark R. Bakker. 2018. "Effects of Management Practices and Topography on Ectomycorrhizal Fungi of Maritime Pine during Seedling Recruitment" Forests 9, no. 5: 245. https://doi.org/10.3390/f9050245