Willow Short Rotation Coppice Trial in a Former Mining Area in Northern Spain: Effects of Clone, Fertilization and Planting Density on Yield after Five Years



Abstract
:1. Introduction
2. Materials and Methods
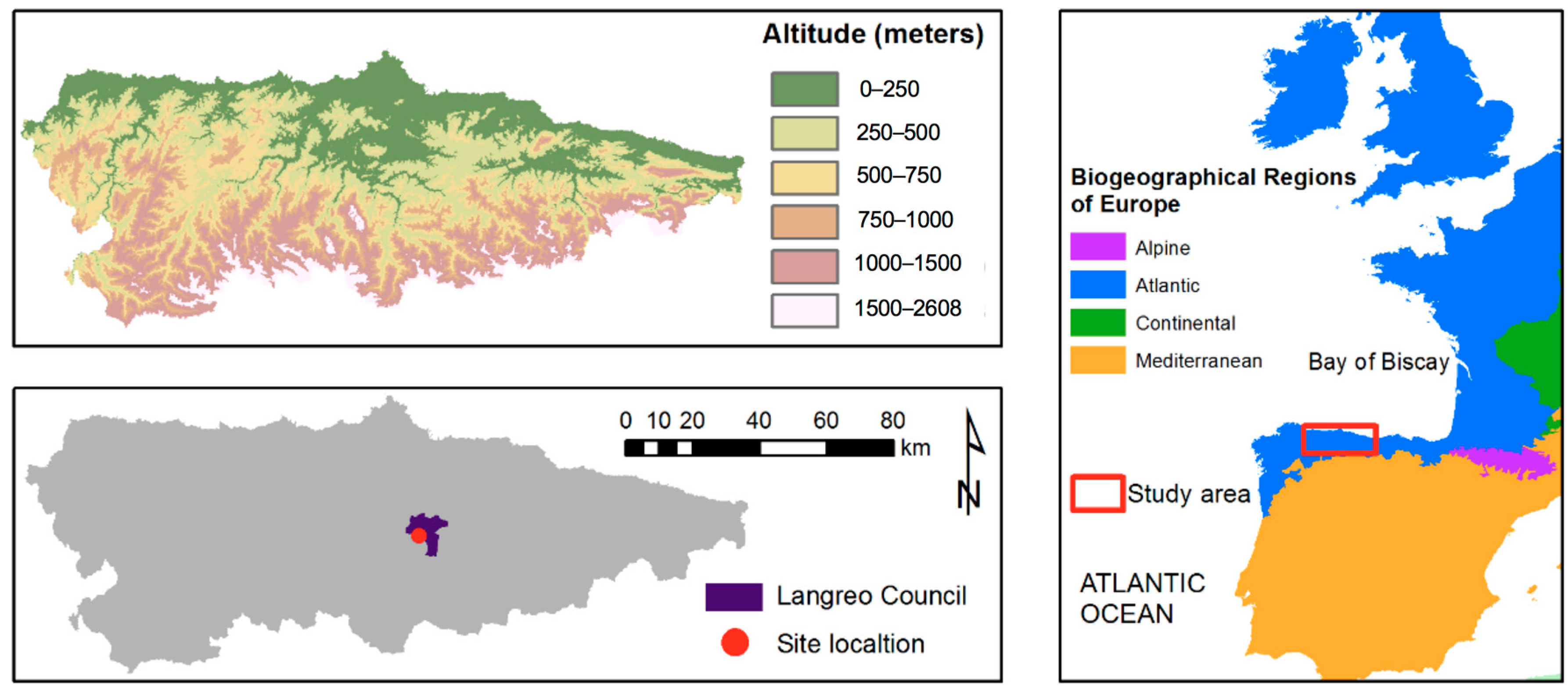
2.1. Site Description
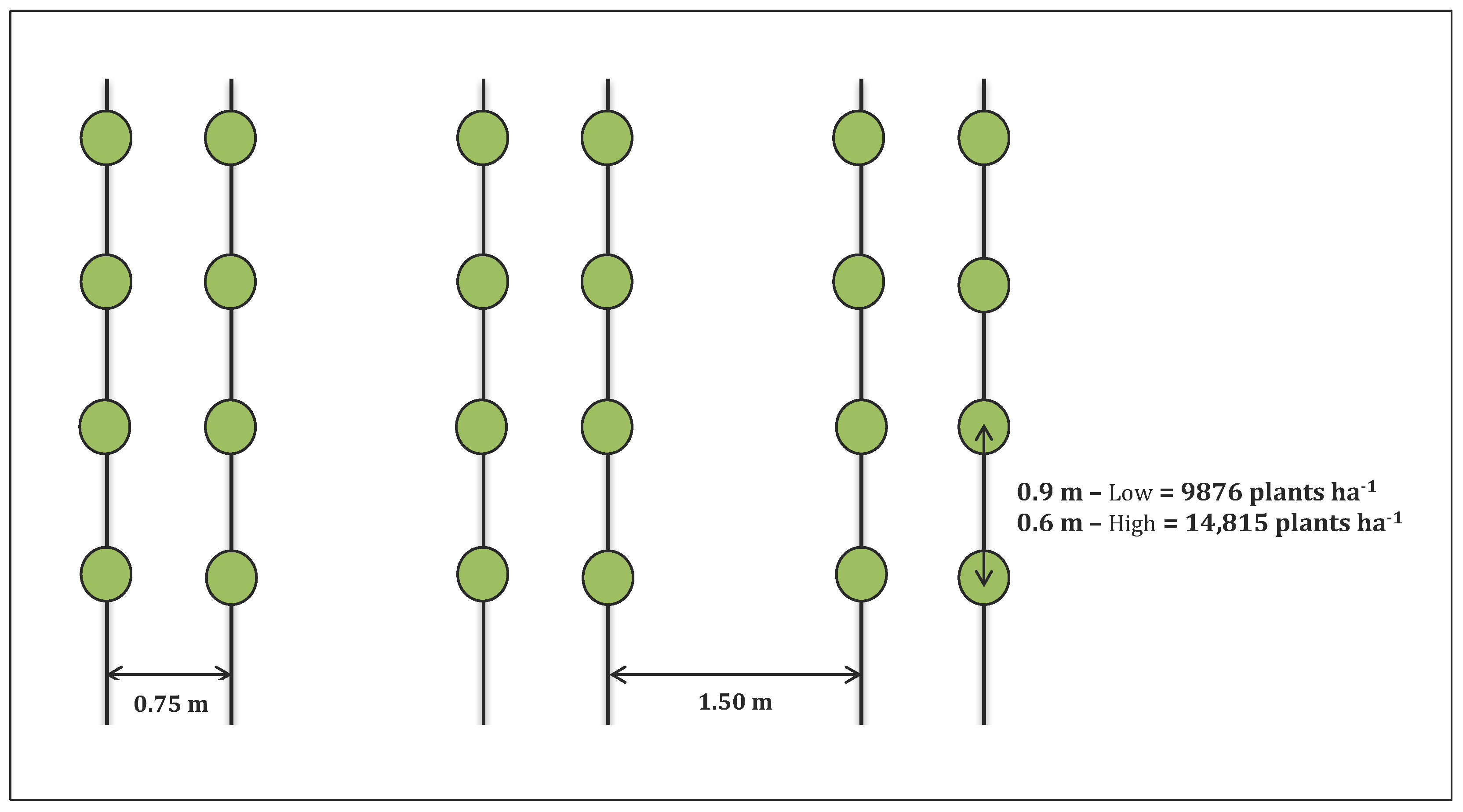
2.2. Experimental Design
2.3. Field Measurements and Yield Estimates
2.4. Statistical Analyses
3. Results
3.1. Number of Shoots per Stool
3.2. Tree Dimensions (H and D20)
3.3. Mortality
3.4. Yield (Dry Weight)
4. Discussion
4.1. Clone Factor
4.2. Planting Density Response
4.3. Response to Fertilization and Herbicide
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- European Commission. Energy 2020. A strategy for Competitive, Sustainable and Secure Energy. 2010. Available online: https://ec.europa.eu/energy/en/topics/energy-strategy/2020-energy-strategy (accessed on 18 November 2017).
- Edenhofer, O.; Madruga, R.P.; Sokona, Y.; Seyboth, K.; Matschoss, K.P.; Kadner, S.; Zwickel, T.; Eickemeier, P.; Hansen, G.; Schlömer, S.; et al. Renewable Energy Sources and Climate Change Mitigation: Special Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2011. [Google Scholar]
- Bentsen, N.S.; Felby, C. Biomass for energy in the European Union—A review of bioenergy resource assessments. Biotechnol. Biofuels 2012, 5, 25. [Google Scholar] [CrossRef] [PubMed]
- Rosso, L.; Facciotto, G.; Bergante, S.; Vietto, L.; Nervo, G. Selection and testing of Populus alba and Salix spp. as bioenergy feedstock: Preliminary results. Appl. Energy 2013, 102, 87–92. [Google Scholar] [CrossRef]
- Paredes-Sánchez, J.P.; García-Elcoro, V.E.; Rosillo-Calle, F.; Xiberta-Bernat, J. Assessment of forest bioenergy potential in a coal-producing area in Asturias (Spain) and recommendations for setting up a Biomass Logistic Centre (BLC). Appl. Energy 2016, 171, 133–141. [Google Scholar] [CrossRef]
- Castaño-Díaz, M.; Álvarez-Álvarez, P.; Tobin, B.; Nieuwenhuis, M.; Afif-Khouri, E.; Cámara-Obregón, A. Evaluation of the use of low-density LiDAR data to estimate structural attributes and biomass yield in a short-rotation willow coppice: An example in a field trial. Ann. For. Sci. 2017, 74, 69. [Google Scholar] [CrossRef]
- Instituto para la Diversificación y el Ahorro de la Energía (IDEA). Biomasa: Cultivos Energéticos; Instituto para la Diversificación y el Ahorro de la Energía (IDAE): Madrid, Spain, 2007. [Google Scholar]
- Styles, D.; Jones, M. Energy crops in Ireland: Quantifying the potential life-cycle greenhouse gas reductions of energy-crop electricity. Biomass Bioenergy 2007, 31, 759–772. [Google Scholar] [CrossRef]
- Keoleian, G.A.; Volk, T.A. Renewable energy from willow biomass crops: Life cycle energy, environmental and economic performance. Crit. Rev. Plant Sci. 2005, 24, 385–406. [Google Scholar] [CrossRef]
- Avelin, A.; Skvaril, J.; Aulin, R.; Odlare, M.; Dahlquist, E. Forest biomass for bioenergy production—Comparison of different forest species. Energy Procedia 2014, 61, 1820–1823. [Google Scholar] [CrossRef]
- Broeckx, L.S.; Verlinden, M.S.; Ceulemans, R. Establishment and two-year growth of a bio-energy plantation with fast-growing Populus trees in Flanders (Belgium): Effects of genotype and former land use. Biomass Bioenergy 2012, 42, 151–163. [Google Scholar] [CrossRef]
- Zurba, K.; Oertel, C.; Matschullat, J. CO2 emissions from willow and poplar short rotation forestry (SRF) on a derelict mining soil. In Proceedings of the Conference International Forum-Competition of Young Researchers “Topical Issues of Subsoil Usage”, Sankt-Petersburg, Russia, 1–4 April 2013. [Google Scholar]
- Kuzovkina, Y.A.; Quigley, M.F. Willows beyond wetlands: Uses of Salix L. species for environmental projects. Water. Air. Soil Pollut. 2005, 162, 183–204. [Google Scholar] [CrossRef]
- Jama, A.; Nowak, W. Willow (Salix viminalis L.) in purifying sewage sludge treated soils. Pol. J. Agron. 2012, 9, 3–6. [Google Scholar]
- Tilman, D.; Hill, J.; Lehman, C. Carbon—Negative biofuels from low-input high-diversity grassland biomass. Science 2006, 314, 1598–1600. [Google Scholar] [CrossRef] [PubMed]
- Gruenewald, H.; Brandt, B.K.V.; Schneider, B.U.; Bens, O.; Kendzia, G.; Hüttl, R.F. Agroforestry systems for the production of woody biomass for energy transformation purposes. Ecol. Eng. 2007, 29, 319–328. [Google Scholar] [CrossRef]
- Venturi, P.; Gigler, J.K.; Huisman, W. Economical and technical comparison between herbaceous (Miscanthus × giganteus) and woody energy crops (Salix viminalis). Renew. Energy 1999, 16, 1023–1026. [Google Scholar] [CrossRef]
- Tahvanainen, L.; Rytko, V. Biomass production of Salix viminalis in southern Finland and the effect of soil properties and climate conditions on its production and survival. Biomass Bioenergy 1999, 16, 103–117. [Google Scholar] [CrossRef]
- Weih, M. Intensive short rotation forestry in boreal climates: Present and future perspectives. Can. J. For. Res. 2004, 34, 1369–1378. [Google Scholar] [CrossRef]
- Hofmann-Schielle, C.; Jug, A.; Makeschin, F.; Rehfuess, K. Short-rotation plantations of balsam poplars, aspen and willows on former arable land in the Federal Republic of Germany. I. Site-growth relationships. For. Ecol. Manag. 1999, 121, 85–99. [Google Scholar] [CrossRef]
- Lindroth, A.; Båth, A. Assessment of regional willow coppice yield in Sweden on basis of water availability. For. Ecol. Manag. 1999, 121, 57–65. [Google Scholar] [CrossRef]
- Labrecque, M.; Teodorescu, T.I. High biomass yield achieved by Salix clones in SRIC following two 3-year coppice rotations on abandoned farmland in southern Quebec, Canada. Biomass Bioenergy 2003, 25, 135–146. [Google Scholar] [CrossRef]
- Bergkvist, P.; Ledin, S. Stem biomass yields at different planting designs and spacings in willow coppice systems. Biomass Bioenergy 1998, 14, 149–156. [Google Scholar] [CrossRef]
- Willebrand, E.; Ledin, S.; Verwijst, T. Willow coppice systems in short-rotation forestry effects of plant spacing, rotation length and clonal composition on biomass production. Biomass Bioenergy 1993, 4, 323–331. [Google Scholar] [CrossRef]
- Bullard, M.J.; Mustill, S.J.; Carver, P.; Nixon, P.M.I. Yield improvements through modification of planting density and harvest frequency in short rotation coppice Salix spp.—2. Resource capture and use in two morphologically diverse varieties. Biomass Bioenergy 2002, 22, 27–39. [Google Scholar] [CrossRef]
- Stolarski, M.; Szczukowski, S.; Tworkowski, J.; Klasa, A. Productivity of seven clones of willow coppice in annual and quadrennial cutting cycles. Biomass Bioenergy 2008, 32, 1227–1234. [Google Scholar] [CrossRef]
- Sage, R.B. Weed competition in willow coppice crops: The cause and extent of yield losses. Weed Res. 1999, 39, 399–411. [Google Scholar] [CrossRef]
- Karp, A.; Hanley, S.J.; Trybush, S.O.; Macalpine, W.; Pei, M.; Shield, I. Genetic Improvement of Willow for Bioenergy and Biofuels. J. Integr. Plant Biol. 2011, 53, 151–165. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.; Bullard, M.J.; Christian, D.G.; Knight, J.D.; Lainsbury, M.A.; Parker, S.R. Commercial varieties from the Swedish willow breeding programme. Asp. Appl. Biol. 2001, 65, 193–198. [Google Scholar]
- Lindegaard, K.N.; Cater, M.M.; McCracken, A.; Shield, I.F.; Macalpine, W.; Hinton Jones, M.; Valentine, J.; Larsson, S. Comparative trials of elite Swedish and UK biomass willow varieties 2001–2010. Asp. Appl. Biol. 2011, 112, 57–66. [Google Scholar]
- Adegbidi, H.G.; Volk, T.A.; White, E.H.; Abrahamson, L.P.; Briggs, R.D.; Bickelhaupt, D.H. Biomass and nutrient removal by willow clones in experimental bioenergy plantations in New York State. Biomass Bioenergy 2001, 20, 399–411. [Google Scholar] [CrossRef]
- Lazdina, D.; Lazdins, A.; Karins, Z.; Kaposts, V. Effect of sewage sludge fertilization in short-rotation willow plantations. J. Environ. Eng. Landsc. Manag. 2007, 15, 105–111. [Google Scholar] [CrossRef]
- Stolarski, M.; Wróblewska, H.; Szczukowski, S.; Tworkowski, J.; Kwiatkowski, J.; Cichy, W. Charakterystyka biomasy wierzby i slazowca pensylwanskiego jako potencjalnego surowca przemyslowego. Fragm. Agron. 2006, 3, 277–289. [Google Scholar]
- Sulima, P.; Przyborowski, J.A.; Stolarski, M. Ocena przydatnosci wybranych gatunkow wierzby do celow energetycznych. Fragm. Agron. 2006, 23, 290–299. [Google Scholar]
- Abrahamson, L.; Volk, T.; Smart, L.; Cameron, K. Shrub Willow Biomass Producer’s Handbook; College of Environmental Science and Forestry: Syracuse, NY, USA, 2010. [Google Scholar]
- European Environment Agency (EEA). Biogeographical Regions. 2011. Available online: http://www.eea.europa.eu/dataand-maps/data/biogeographical-regions-europe-1 (accessed on 9 October 2017).
- Gee, G.W.; Bauder, J.W. Particle-size analysis. In Methods of Soil Analysis. Part 1. Physical and Mineralogical Methods, 2nd ed.; Klute, A., Ed.; Soil Science Society of America, American Society of Agronomy (ASA–SSA): Madison, WI, USA, 1986; pp. 383–411. [Google Scholar]
- Klute, A. Nitrogen-total. In Methods of Soil Analyses, Part 1; Klute, A., Ed.; Soil Science Society of America, American Society of Agronomy (ASA–SSA): Madison, WI, USA, 1996; pp. 595–624. [Google Scholar]
- Mehlich, A. Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant. Soil Sci. Plant Anal. 1985, 15, 1409–1416. [Google Scholar] [CrossRef]
- Lindegaard, K.N.; Barker, J.H.A. Breeding willows for biomass. Asp. Appl. Biol. 1996, 49, 155–162. [Google Scholar]
- Larsson, S. Genetic improvement of willow for short- rotation coppice. Biomass Bioenergy 1998, 15, 23–26. [Google Scholar] [CrossRef]
- Forest Research. Mensurational variables protocol. In Yield Models for Energy Coppice of Poplar and Willow; Forest Research: Farnham, UK, 2003. [Google Scholar]
- IBM Corp. Released, IBM SPSS Statistics forWindows, Version 23.0; IBM Corp: Armonk, NT, USA, 2015. [Google Scholar]
- Hytönen, J.; Nurmi, J. Heating value and ash content of intensively managed stands. Wood Res. 2015, 60, 71–82. [Google Scholar]
- Morhart, C.; Sheppard, J.; Seidl, F.; Spiecker, H. Influence of different tillage systems and weed treatments in the establishment year on the final biomass production of short rotation coppice poplar. Forests 2013, 4, 849–867. [Google Scholar] [CrossRef]
- Bungart, R.; Huttl, R.F. Production of biomass for energy in post-mining landscapes and nutrient dynamics. Biomass Bioenergy 2001, 20, 181–187. [Google Scholar] [CrossRef]
- Grünewald, H.; Böhm, C.; Quinkenstein, A.; Grundmann, P.; Eberts, J.; von Wühlisch, G. Robinia pseudoacacia L.: A lesser known tree species for biomass production. Bioenergy Res. 2009, 2, 123–133. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Szczukowski, S.; Tworkowski, J.; Klasa, A. Willow biomass production under conditions of low-input agriculture on marginal soils. For. Ecol. Manag. 2011, 262, 1558–1566. [Google Scholar] [CrossRef]
- Nissim, W.G.; Pitre, F.E.; Teodorescu, T.I.; Labrecque, M. Long-term biomass productivity of willow bioenergy plantations maintained in southern Quebec, Canada. Biomass Bioenergy 2013, 56, 361–369. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Szczukowski, S.; Tworkowski, J.; Klasa, A. Yield, energy parameters and chemical composition of short-rotation willow biomass. Ind. Crops Prod. 2013, 46, 60–65. [Google Scholar] [CrossRef]
- Volk, T.; Kiernan, B.D.; Kopp, R.; Abrahamson, L. First-and second-rotation yields of willow clones at two sites in New York State. Fifth Biomass Conf. Am. 2001, 13210, 17–21. [Google Scholar]
- Ceulemans, R.; McDonald, A.J.S.; Pereira, J.S. A comparison among eucalypt, poplar and willow characteristics with particular reference to a coppice, growth-modelling approach. Biomass Bioenergy 1996, 11, 215–231. [Google Scholar] [CrossRef]
- Department for Environment, Food & Rural Affairs (DEFRA). Growing Short Rotation Coppic; DEFRA: London, UK, 2004; pp. 1–30.
- Mola-Yudego, B.; Aronsson, P. Yield models for commercial willow biomass plantations in Sweden. Biomass Bioenergy 2008, 32, 829–837. [Google Scholar] [CrossRef]
- McIvor, I.; Snowdon, K.; Nicholas, I.D. Effect of species and management on root development in SRC willow. In IEA Task 30 Conference Paper 2009; International Energy Agency: Paris, France, 2009. [Google Scholar]
- Wilkinson, J.; Evans, E.; Bilsborrow, P.; Wright, C.; Hewison, W.; Pilbeam, D. Yield of willow cultivars at different planting densities in a commercial short rotation coppice in the north of England. Biomass Bioenergy 2007, 31, 469–474. [Google Scholar] [CrossRef]
- DeBell, D.S.; Clendenen, G.W.; Harrington, C.A.; Zasada, J.C. Tree growth and stand development in short-rotation Populus plantings: 7-year results for two clones at three spacings. Biomass Bioenergy 1996, 11, 253–269. [Google Scholar] [CrossRef]
- Hansen, E.; Gobakken, T.; Solberg, S.; Kangas, A.; Ene, L.; Mauya, E.; Næsset, E. Relative efficiency of ALS and InSAR for biomass estimation in a Tanzanian rainforest. Remote Sens. 2015, 7, 9865–9885. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Lehto, T.; Piirainen, S.; Kukkonen, J.V.K.; Pelkonen, P. Effects of planting orientation and density on the soil solution chemistry and growth of willow cuttings. Biomass Bioenergy 2012, 46, 165–173. [Google Scholar] [CrossRef]
- Tubby, I.; Armstrong, A. Establishment and management of short rotation coppice. In Practice Note; Forestry Commission: Bristol, UK, 2002. [Google Scholar]
- Cao, Y.; Lehto, T.; Repo, T.; Silvennoinen, R.; Pelkonen, P. Effects of planting orientation and density of willows on biomass production and nutrient leaching. New For. 2011, 41, 361–377. [Google Scholar] [CrossRef]
- Senelwa, K.; Sims, R.E.H. Fuel characteristics of short rotation forest biomass. Biomass Bioenergy 1999, 17, 127–140. [Google Scholar] [CrossRef]
- Szczukowski, S.; Stolarski, M.; Tworkowski, J.; Przyborowski, J.; Klasa, A. Productivity of willow coppice plants grown in short rotations. Plant Soil Environ. 2005, 51, 423–430. [Google Scholar] [CrossRef]
- Ferm, A.; Vuori, J. Effects of spacing and nitrogen fertilization on the establishment and biomass production of short rotation poplar in Finland. Biomass 1989, 18, 95–108. [Google Scholar] [CrossRef]
- Klasnja, B.; Kopitovic, S.; Orlovic, S. Wood and bark of some poplar and willow clones as fuelwood. Biomass Bioenergy 2010, 23, 427–432. [Google Scholar] [CrossRef]
- DeBell, D.; Clendenen, G.; Zasadat, J. Growing Populus biomass: Comparison of woodgrass versus wider-spaced short-rotation systems. Biomass Bioenergy 1993, 4, 305–313. [Google Scholar] [CrossRef]
- Johnstone, W.D. The effects of initial spacing and rectangularity on the early growth of hybrid poplar. West. J. Appl. For. 2008, 23, 189–196. [Google Scholar]
- Strong, T.; Hansen, E. Hybrid poplar spacing/productivity relations in short rotation intensive culture plantations. Biomass Bioenergy 1993, 4, 255–261. [Google Scholar] [CrossRef]
- Proe, M.F.; Gri, J.H.; Craig, J. Effects of spacing, species and coppicing on leaf area, light interception and photosynthesis in short rotation forestry. Biomass Bioenergy 2002, 23, 315–326. [Google Scholar] [CrossRef]
- Miller, R.O.; Bender, B.A.; Irving, P.N.; Zuidema, K.T. Common short rotation poplar growth patterns observed in ten trials over 18 years in Michigan, USA. In Proceedings of the 25th International Poplar Symposium, Berlin, Germany, 13–16 September 2016; pp. 1–13. [Google Scholar]
- Verwijst, T.; Telenius, B. Biomass estimation procedures in short rotation forestry. For. Ecol. Manag. 1999, 121, 137–146. [Google Scholar] [CrossRef]
- Willebrand, E.; Verwijst, T. Willow coppice systems in short rotation forestry: The influence of plant spacing and rotation length on the sustainability of biomass. In Proceedings of the 7th European Conference on Biomass for Energy and Environment, Agriculture and Industry, Florence, Italy, 5–9 October 1992; pp. 472–477. [Google Scholar]
- Hytönen, J. Effects of liming on the growth of birch and Willow on cut-away peat substrates in greenhouse. Balt. For. 2005, 11, 68–74. [Google Scholar]
- Barrio, M.; Montoto, J.L.; Pérez, J.; Mazón, P.; Ciria, P.; Sixto, H. Influence of fertilization and weed control following the first growth period in multiclonal poplar plantations in central Spain. In Proceedings of the 15th European Biomass Conference and Exhibition, Berlin, Germany, 7–11 May 2007; pp. 736–740. [Google Scholar]
- Mitchell, C.P.; Stevens, E.A.; Watters, M.P. Short-rotation forestry—Operations, productivity and costs based on experience gained in the UK. For. Ecol. Manag. 1999, 121, 123–136. [Google Scholar] [CrossRef]
- Bergkvist, P.; Nordh, N.; Ledin, S.; Olsson, T. Plant material for short rotation forestry. In Handbook on How to Grow Short Rotation Forests; Willebrand, E., Ed.; Swedish University of Agricultural Sciences, Department of Short Rotation Forestry: Uppsala, Sweden, 1996; pp. 1–11. ISBN 9157651558, 9789157651556. [Google Scholar]
- Sevel, L.; Ingerslev, M.; Nord-Larsen, T.; Jørgensen, U.; Holm, P.E.; Schelde, K.; Raulund-Rasmussen, K. Fertilization of SRC Willow, II: Leaching and Element Balances. Bioenergy Res. 2014, 7, 338–352. [Google Scholar] [CrossRef]
- Alriksson, B.; Ledin, S.; Seeger, P. Effect of nitrogen fertilization on growth in a Salix viminalis stand using a response surface experimental design. Scand. J. For. Res. 1997, 12, 321–327. [Google Scholar] [CrossRef]
- Kopp, R.F.; Abrahamson, L.P.; White, E.H.; Volk, T.A.; Nowak, C.A.; Fillhart, R.C. Willow biomass production during ten successive annual harvests. Biomass Bioenergy 2001, 20, 1–7. [Google Scholar] [CrossRef]
- Heinsoo, K.; Sild, E.; Koppel, A. Estimation of shoot biomass productivity in Estonian Salix plantations. For. Ecol. Manag. 2002, 170, 67–74. [Google Scholar] [CrossRef]
- Sevel, L.; Nord-Larsen, T.; Ingerslev, M.; Jørgensen, U.; Raulund-Rasmussen, K. Fertilization of SRC Willow, I: Biomass Production Response. Bioenergy Res. 2014, 7, 319–328. [Google Scholar] [CrossRef]
- Georgiadis, P.; Sevel, L.; Raulund-Rasmussen, K.; Stupak, I. Fertilization of willow coppice over three consecutive 2-year rotations—Effects on biomass production, soil nutrients and water. Bioenergy Res. 2017, 10, 728–739. [Google Scholar] [CrossRef]
- Holm, B.; Heinsoo, K. Municipal wastewater application to Short Rotation Coppice of willows—Treatment efficiency and clone response in Estonian case study. Biomass Bioenergy 2013, 57, 126–135. [Google Scholar] [CrossRef]
- Aronsson, P.; Rosenqvist, H.; Dimitriou, I. Impact of nitrogen fertilization to short-rotation willow coppice plantations grown in Sweden on yield and economy. Bioenergy Res. 2014, 7, 993–1001. [Google Scholar] [CrossRef]
- Böhm, C.; Quinkenstein, A.; Freese, D.; Hüttl, R.F. Assessing the short rotation woody biomass production on marginal post-mining areas. J. For. Sci. 2011, 57, 303–311. [Google Scholar] [CrossRef]
- Mosseler, A.; Zsuffa, L.; Stoehr, M.U.; Kenney, W.A. Variation in biomass production, moisture content, and specific gravity in some North American willows (Salix L.). Can. J. For. Res. 1988, 18, 1535–1540. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Site Description | Parameters | Units | Mozquita Trial |
|---|---|---|---|
| Site conditions | Location | Mozquita (Langreo-Asturias) | |
| Latitude | 43°16′15.90″ N | ||
| Longitude | 5°41′55.75″ N | ||
| Elevation | m | 597 | |
| Mean slope | % | 19 | |
| Mean soil depth | m | 0.36 | |
| Previous crops | Herbaceous | ||
| Annual average temperature | °C | 13 | |
| Annual max. temperature | °C | 18 | |
| Annual min. temperature | °C | 7 | |
| Annual precipitation | mm | 1115 | |
| Initial soil properties | pH | H2O (1:2.5) | 6.68 |
| Electrical conductivity | dS m−1 | 0.23 | |
| Sand | % | 35.34 | |
| Clay | % | 27.32 | |
| Organic Matter | % | 1.61 | |
| N total | % | 0.11 | |
| C/N ratio | 8.80 | ||
| Available P (Mehlich 3) | mg kg−1 | 7.34 | |
| Extractable Na | cmol(+) kg−1 | 1.48 | |
| Extractable K | cmol(+) kg−1 | 0.39 | |
| Extractable Ca | cmol(+) kg−1 | 5.71 | |
| Extractable Mg | cmol(+) kg−1 | 3.57 | |
| Extractable Al | cmol(+) kg−1 | 1.75 | |
| ECEC | cmol(+) kg−1 | 12.91 |
| Characteristics | Mozquita Trial | |||
|---|---|---|---|---|
| Total area | 2.3 ha | |||
| Experimental design | Randomized complete blocks | |||
| Number of replicates | 3 | |||
| Species | Salix spp. | |||
| Clones tested | Björn (B), Inger (I) and Olof (O) | |||
| Origin of clones tested | Sweden | |||
| Progenitor | B: Salix schwerinii E. Wolf × Salix viminalis L. | |||
| I: Salix triandra L. × Salix viminalis L. | ||||
| O: Salix viminalis L. × (Salix schwerinii E. Wolf × Salix viminalis L.) | ||||
| Treatments | Planting density | Low | High | |
| 9876 plants·ha−1 | 14,815 plants·ha−1 | |||
| F0 (control) | F1 | F2 | ||
| Fertilization | None | N-P-K 6:20:12 300 kg ha−1 a−1 | N-P-K 6:20:12 600 kg ha−1 a−1 | |
| Herbicide | None | Application of glyphosate 4 l ha−1 a−1 | Application of glyphosate 4 l ha−1 a−1 | |
| Clone /Variables | n | Minimum | Maximum | Mean | Standard Deviation |
|---|---|---|---|---|---|
| Björn | |||||
| Shoots per stool (n) | 18 | 1.0 | 3.4 | 1.5 | 0.5 |
| H (cm) | 18 | 99 | 397 | 228 | 97 |
| D20 (cm) | 18 | 0.8 | 3.2 | 1.7 | 0.7 |
| Mortality rate (%) | 18 | 2.5 | 100.0 | 42.5 | 25.5 |
| DW (Mg·ha−1) | 18 | 0.2 | 10.2 | 3.9 | 3.1 |
| Inger | |||||
| Shoots per stool (n) | 18 | 1.3 | 3.2 | 2.3 | 0.5 |
| H (cm) | 18 | 67 | 417 | 241 | 87 |
| D20 (cm) | 18 | 0.6 | 2.6 | 1.8 | 0.5 |
| Mortality rate (%) | 18 | 0.0 | 77.5 | 17.7 | 21.0 |
| DW (Mg·ha−1) | 18 | 0.2 | 15.4 | 6.8 | 4.8 |
| Olof | |||||
| Shoots per stool (n) | 18 | 1.1 | 1.8 | 1.4 | 0.2 |
| H (cm) | 18 | 177 | 697 | 468 | 164 |
| D20 (cm) | 18 | 1.3 | 3.8 | 2.6 | 0.8 |
| Mortality rate (%) | 18 | 0.0 | 27.5 | 11.7 | 7.6 |
| DW (Mg·ha−1) | 18 | 1.1 | 30.8 | 14.1 | 9.1 |
| Shoots per Stool (n) | H (cm) | D20 (cm) | Mortality Rate (%) | DW (Mg·ha−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | |
| Clone | 27.797 | <0.001 ** | 14.806 | <0.001 ** | 7.337 | 0.002 ** | 12.309 | <0.001 ** | 9.084 | <0.001 ** |
| Planting density | 0.036 | 0.850 | 0.115 | 0.736 | 0.188 | 0.667 | 0.503 | 0.481 | 0.041 | 0.840 |
| Treatment | 3.317 | 0.044 * | 8.763 | <0.001 ** | 10.644 | <0.001 ** | 0.519 | 0.598 | 10.070 | <0.001 ** |
| Block | 0.446 | 0.643 | 0.745 | 0.480 | 0.798 | 0.456 | 2.814 | 0.069 | 0.506 | 0.606 |
| Clone × planting density | 11.173 | <0.001 ** | 6.393 | <0.001 ** | 3.545 | 0.008 ** | 0.519 | 0.630 | 3.699 | 0.007 ** |
| Clone × treatment | 10.303 | <0.001 ** | 17.721 | <0.001 ** | 9.646 | <0.001 ** | 0.540 | 0.711 | 8.108 | <0.001 ** |
| Planting density × treatment | 1.952 | 0.104 | 3.546 | 0.008 ** | 4.538 | 0.002 ** | 0.399 | 0.695 | 4.008 | 0.004 ** |
| Shoots per Stool (n) | H (cm) | D20 (cm) | Mortality Rate (%) | DW (Mg·ha−1) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | M | G | F | M | G | F | M | G | F | M | G | F | M | G | |
| Clone | B | 1.4 | b | B | 206 | b | B | 1.6 | b | B | 42.5 | b | B | 1.3 | b |
| I | 2.2 | a | I | 222 | b | I | 1.7 | b | I | 17.9 | a | I | 3.6 | ab | |
| O | 1.3 | b | O | 434 | a | O | 2.5 | a | O | 11.9 | a | O | 8.6 | a | |
| Treatment | F0 | 1.4 | a | F0 | 181 | b | F0 | 1.3 | b | - | - | - | F0 | 1.1 | b |
| F1 | 1.7 | a | F1 | 315 | a | F1 | 2.1 | a | - | - | - | F1 | 5.4 | a | |
| F2 | 1.8 | a | F2 | 344 | a | F2 | 2.3 | a | - | - | - | F2 | 6.6 | a | |
| Clone × Density | BN1 | 1.3 | b | BN1 | 236 | b | BN1 | 1.8 | a | - | - | - | BN1 | 1.6 | b |
| IN1 | 2.3 | a | IN1 | 218 | b | IN1 | 1.6 | a | - | - | - | IN1 | 3.3 | ab | |
| ON1 | 1.0 | b | ON1 | 394 | a | ON1 | 2.3 | a | - | - | - | ON1 | 7.0 | a | |
| BN2 | 1.5 | b | BN2 | 177 | b | BN2 | 1.3 | b | - | - | - | BN2 | 1.0 | b | |
| IN2 | 2.1 | a | IN2 | 226 | b | IN2 | 1.7 | b | - | - | - | IN2 | 3.8 | ab | |
| ON2 | 1.4 | b | ON2 | 478 | a | ON2 | 2.6 | a | - | - | - | ON2 | 10.7 | a | |
| Clone × Treatment | BF0 | 1.2 | a | BF0 | 142 | b | BF0 | 1.1 | b | - | - | - | BF0 | 0.4 | b |
| BF1 | 1.4 | a | BF1 | 236 | ab | BF1 | 1.7 | ab | - | - | - | BF1 | 1.3 | ab | |
| BF2 | 1.7 | a | BF2 | 290 | a | BF2 | 2.1 | a | - | - | - | BF2 | 3.3 | a | |
| IF0 | 1.8 | b | IF0 | 174 | a | IF0 | 1.4 | a | - | - | - | IF0 | 2.2 | b | |
| IF1 | 2.4 | a | IF1 | 276 | a | IF1 | 1.9 | a | - | - | - | IF1 | 8.6 | a | |
| IF2 | 2.6 | a | IF2 | 274 | a | IF2 | 2.0 | a | - | - | - | IF2 | 6.8 | ab | |
| OF0 | 1.2 | b | OF0 | 275 | b | OF0 | 1.8 | b | - | - | - | OF0 | 3.1 | b | |
| OF1 | 1.6 | a | OF1 | 571 | a | OF1 | 3.1 | a | - | - | - | OF1 | 17.6 | a | |
| OF2 | 1.4 | ab | OF2 | 559 | a | OF2 | 3.0 | a | - | - | - | OF2 | 16.8 | a | |
| Density × Treatment | - | - | - | N1F0 | 180 | b | N1F0 | 1.4 | b | - | - | - | N1F0 | 0.9 | b |
| - | - | - | N1F1 | 324 | a | N1F1 | 2.3 | a | - | - | - | N1F1 | 6.7 | a | |
| - | - | - | N1F2 | 347 | a | N1F2 | 2.4 | a | - | - | - | N1F2 | 5.7 | a | |
| - | - | - | N2F0 | 183 | a | N2F0 | 1.3 | b | - | - | - | N2F0 | 1.2 | b | |
| - | - | - | N2F1 | 287 | a | N2F1 | 1.9 | ab | - | - | - | N2F1 | 4.4 | a | |
| - | - | - | N2F2 | 366 | a | N2F2 | 2.3 | a | - | - | - | N2F2 | 7.6 | a | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castaño-Díaz, M.; Barrio-Anta, M.; Afif-Khouri, E.; Cámara-Obregón, A. Willow Short Rotation Coppice Trial in a Former Mining Area in Northern Spain: Effects of Clone, Fertilization and Planting Density on Yield after Five Years. Forests 2018, 9, 154. https://doi.org/10.3390/f9030154
Castaño-Díaz M, Barrio-Anta M, Afif-Khouri E, Cámara-Obregón A. Willow Short Rotation Coppice Trial in a Former Mining Area in Northern Spain: Effects of Clone, Fertilization and Planting Density on Yield after Five Years. Forests. 2018; 9(3):154. https://doi.org/10.3390/f9030154
Chicago/Turabian StyleCastaño-Díaz, María, Marcos Barrio-Anta, Elías Afif-Khouri, and Asunción Cámara-Obregón. 2018. "Willow Short Rotation Coppice Trial in a Former Mining Area in Northern Spain: Effects of Clone, Fertilization and Planting Density on Yield after Five Years" Forests 9, no. 3: 154. https://doi.org/10.3390/f9030154