Natural and Synthetic Hydrophilic Polymers Enhance Salt and Drought Tolerance of Metasequoia glyptostroboides Hu and W.C.Cheng Seedlings


Abstract
:1. Introduction
2. Materials and Methods
2.1. Hydrophilic Polymers
2.2. Plant Material and Treatments
- (1)
- Control (Non-polymer), Control + Stockosorb, Control + Luquasorb, Control + KGM;
- (2)
- NaCl (Non-polymer), NaCl + Stockosorb, NaCl + Luquasorb, NaCl + KGM;
- (3)
- Drought (Non-polymer), Drought + Stockosorb, Drought + Luquasorb, Drought + KGM;
- (4)
- Drought + NaCl (Non-polymer), Drought + NaCl + Stockosorb, Drought + NaCl + Luquasorb, Drought + NaCl + KGM.
2.3. Shoot Height Measurement
2.4. Whole-Plant Water Consumption
2.5. Leaf Gas-Exchange
2.6. Chlorophyll a Fluorescence
2.7. Leaf Membrane Permeability
2.8. Plant Harvest
2.9. Ion Analysis of Leaves and Roots
2.10. Ion Analysis of Soil
2.11. Data Analysis
3. Results
3.1. Occurrence of Leaf Injury under Water and Salt Stress
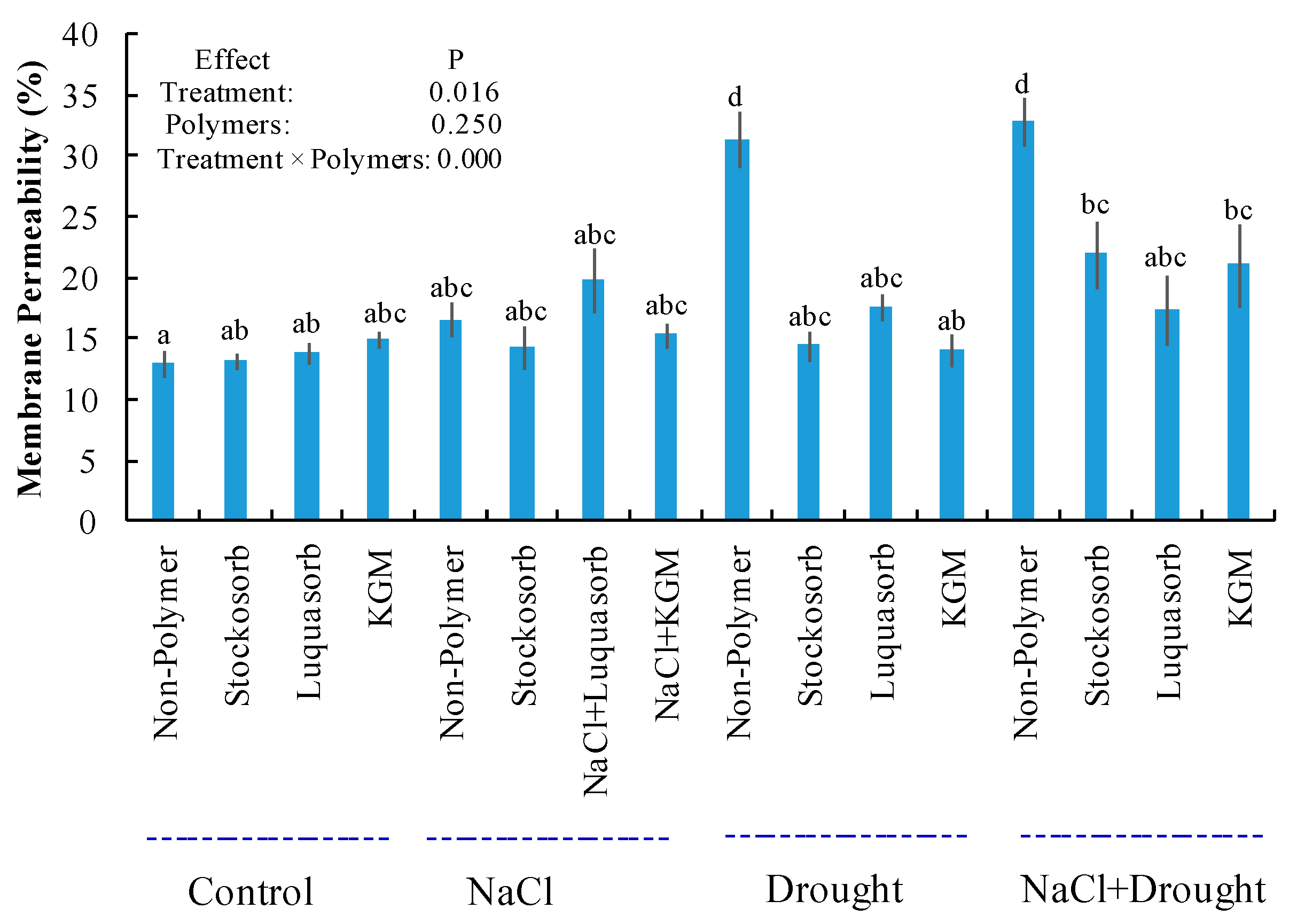
3.2. Leaf Membrane Permeability
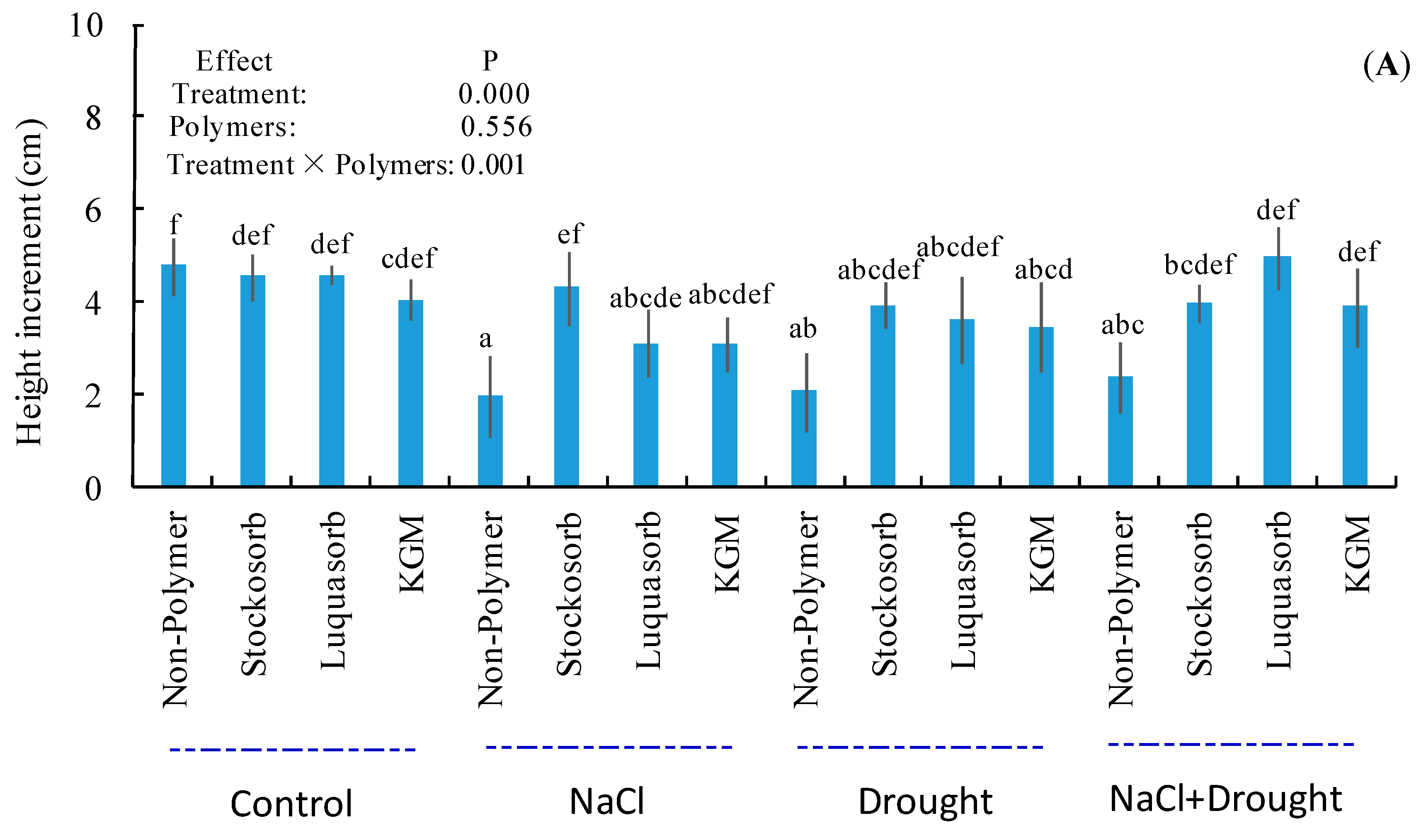
3.3. Shoot Height Increment
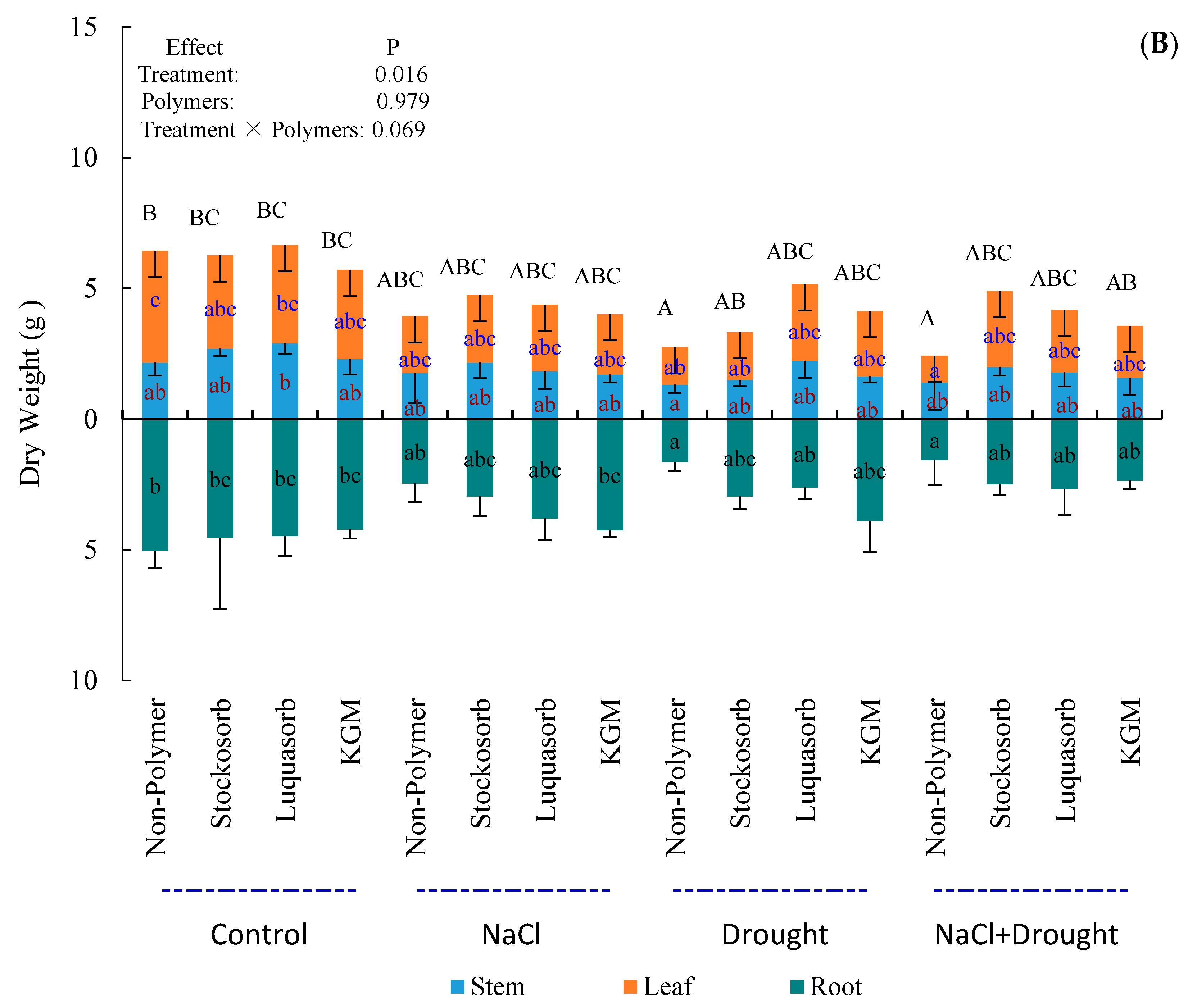
3.4. Plant Dry Weight
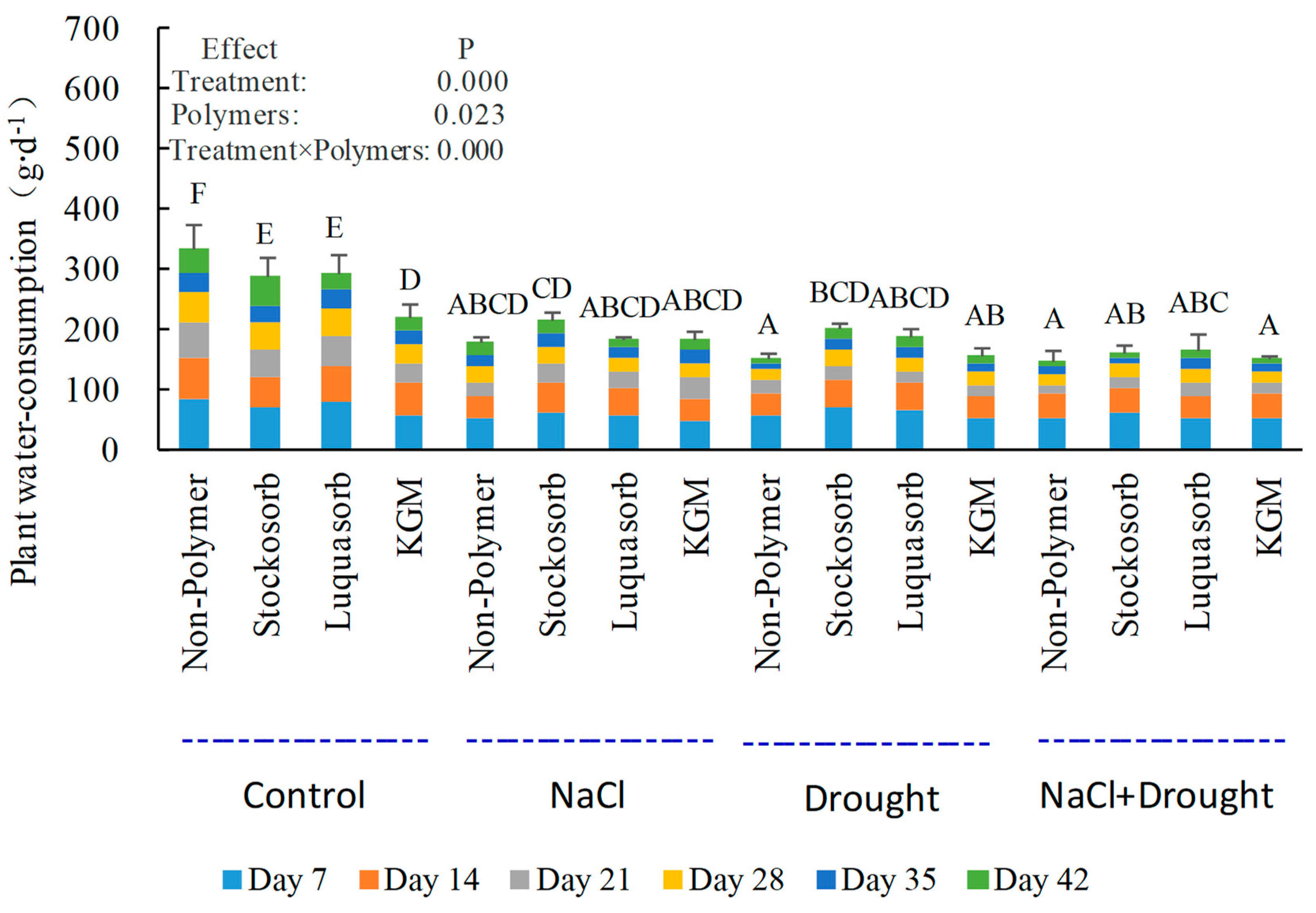
3.5. Whole-Plant Water-Consumption
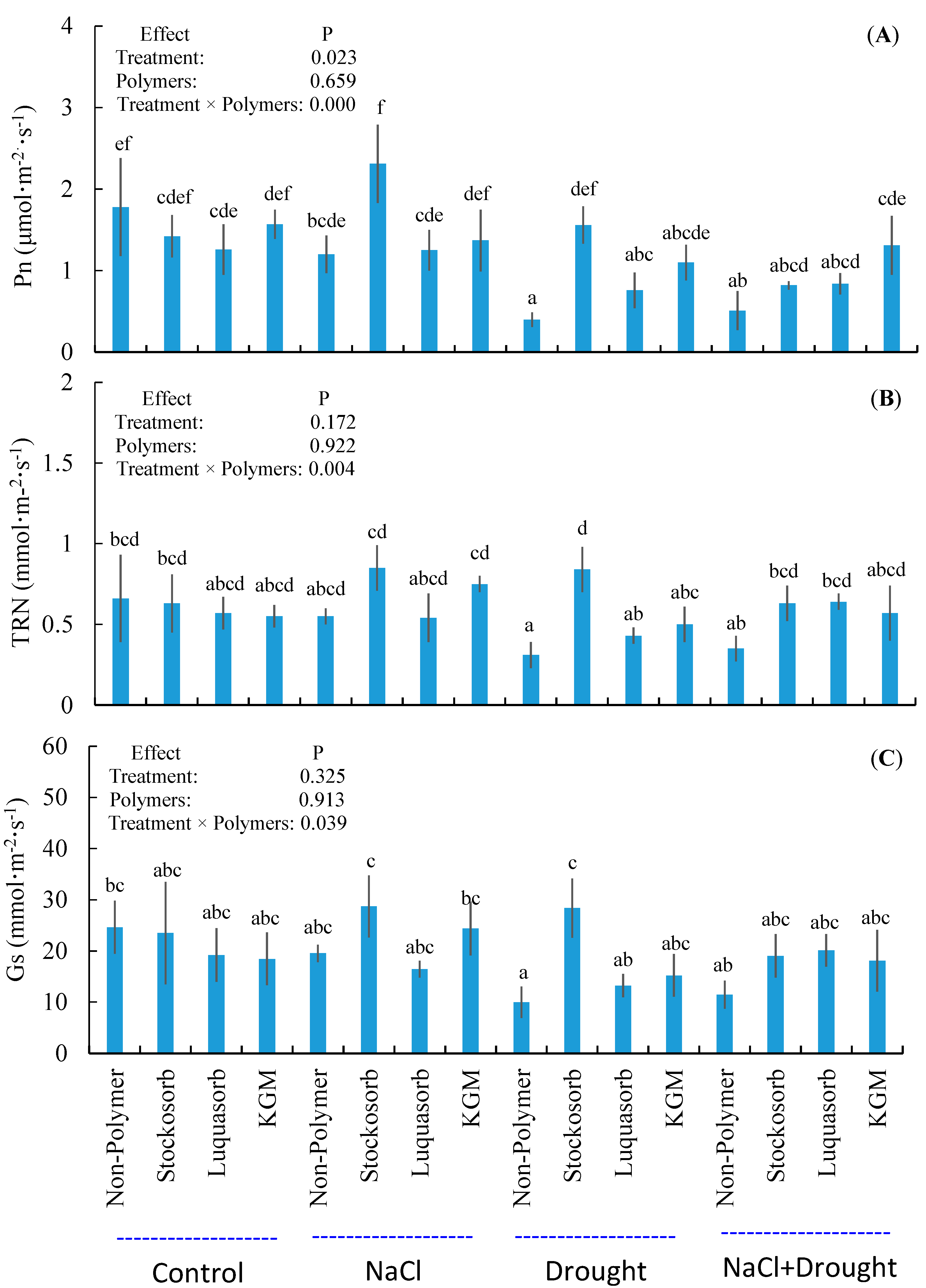
3.6. Leaf Gas-Exchange
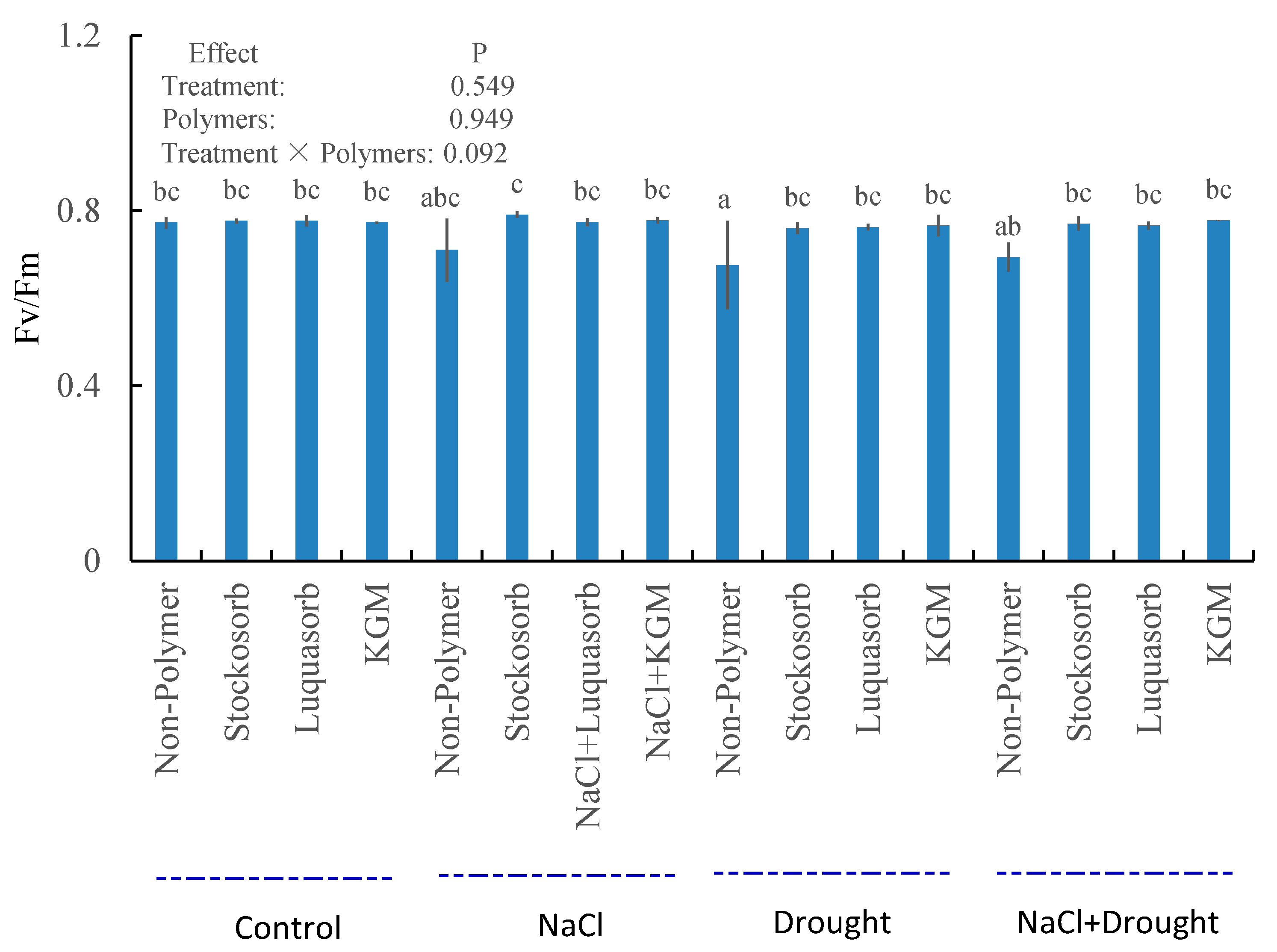
3.7. Chlorophyll a Fluorescence
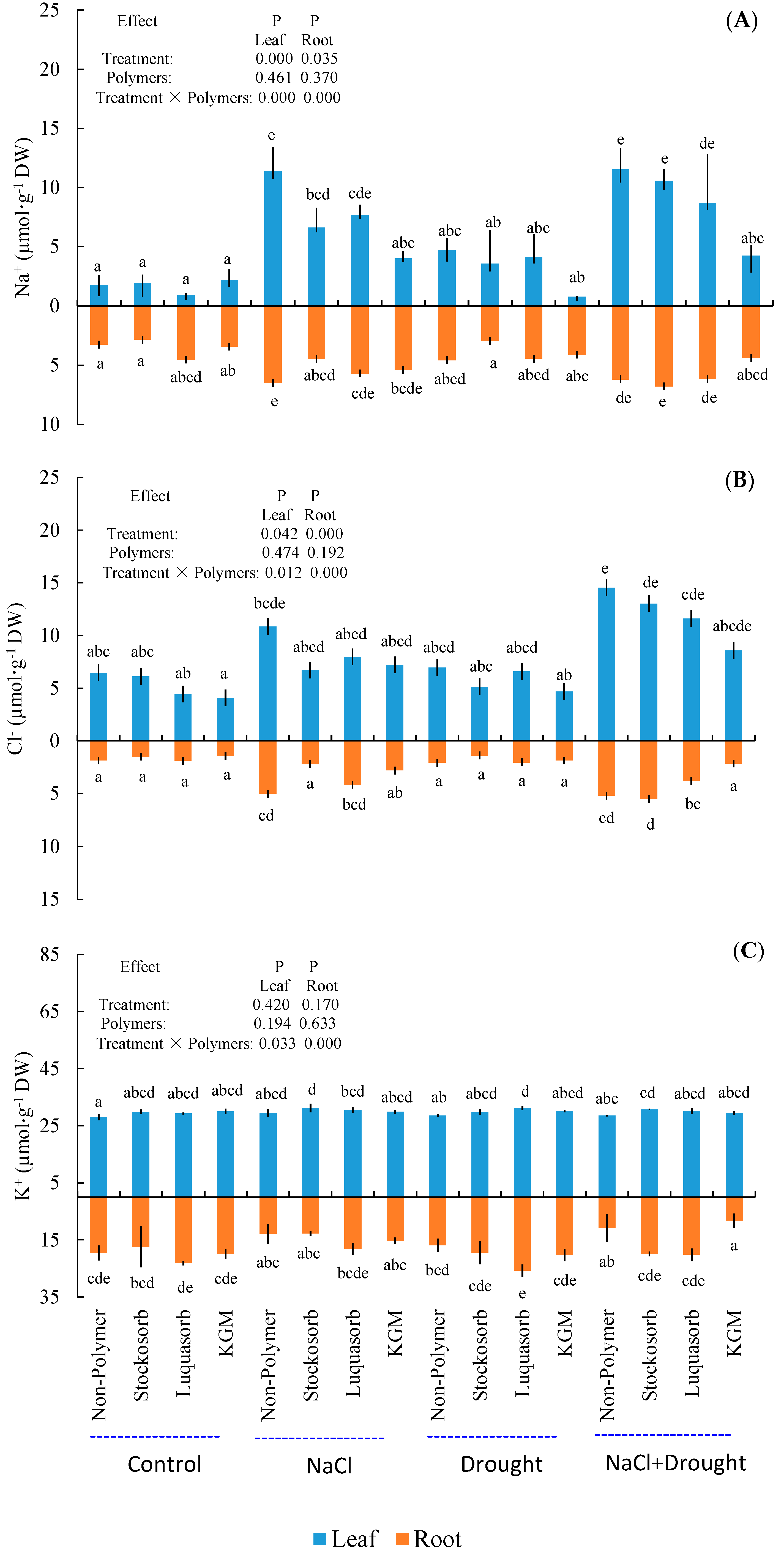
3.8. Ion Concentrations in Roots and Leaves
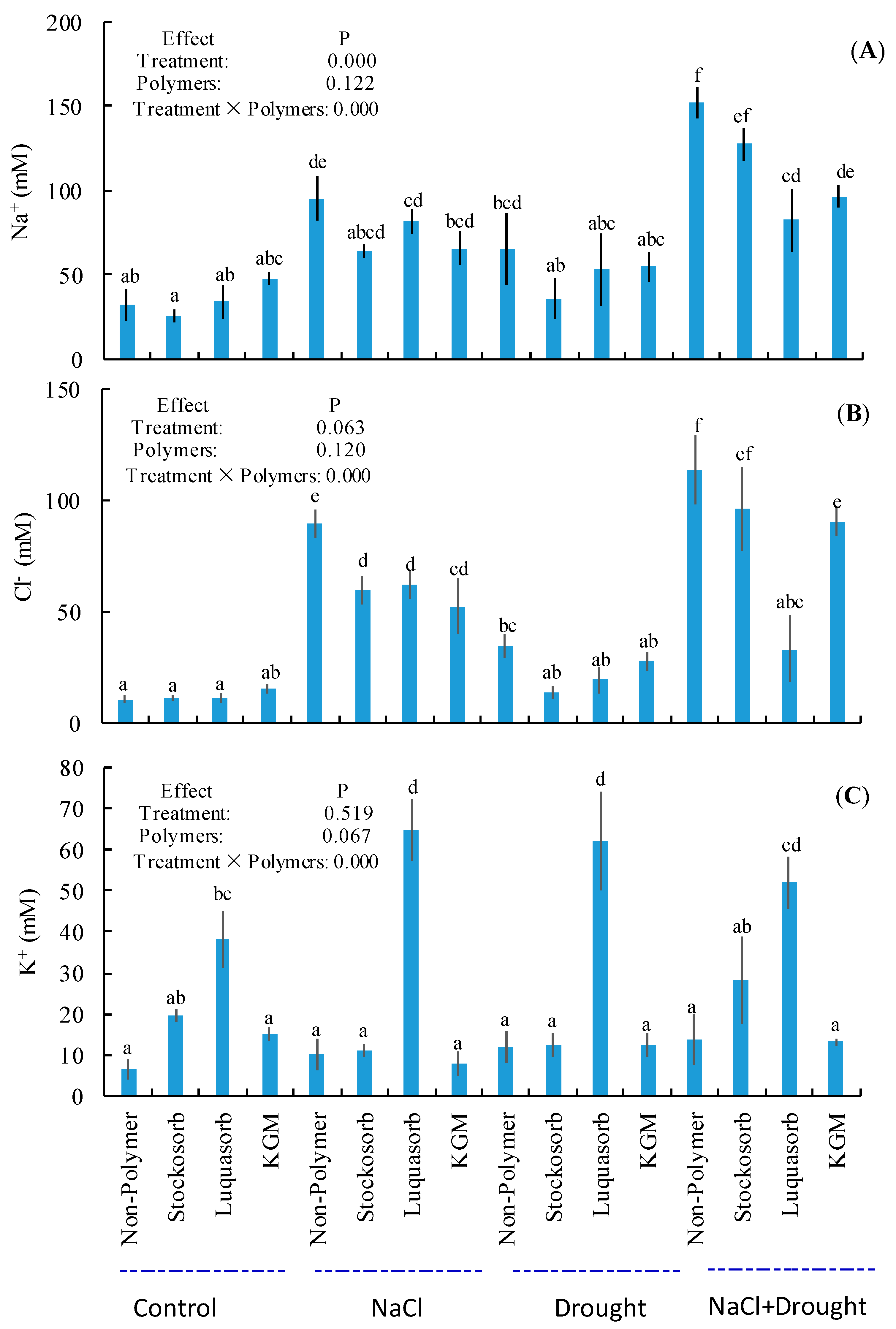
3.9. Ion Concentrations in Soils
4. Discussion
4.1. Hydrophilic Polymers Increase Water Supply under Drought Stress
4.2. Hydrophilic Polymers Alleviate Salt Stress
4.3. Hydrophilic Polymers Enhance Plant Tolerance of Combined Drought and Salt Stresses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kumar, M. Crop plants and abiotic stresses. J. Biomol. Res. Ther. 2013, 3, e125. [Google Scholar] [CrossRef]
- Harfouche, A.; Meilan, R.; Altman, A. Molecular and physiological responses to abiotic stress in forest trees and their relevance to tree improvement. Tree Physiol. 2014, 34, 1181–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polle, A.; Chen, S. On the salty side of life: Molecular, physiological and anatomical adaptation and acclimation of trees to extreme habitats. Plant Cell Environ. 2015, 38, 1794–1816. [Google Scholar] [CrossRef] [PubMed]
- Rennenberg, H.; Loreto, F.; Polle, A.; Brilli, F.; Fares, S.; Beniwal, R.S.; Gessler, A. Physiological responses of forest trees to heat and drought. Plant Biol. 2006, 8, 556–571. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.; Zhang, J.; Ma, Y.; Pan, X.; Dong, C.; Pang, S.; He, S.; Deng, L.; Yi, S.; Zheng, Y.; et al. Combined analysis of mRNA and miRNA identifies dehydration and salinity responsive key molecular players in citrus roots. Sci. Rep. 2017, 7, 42094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.; Choi, J.; An, G.; Kim, S.R. Ectopic expression of OsSta2 enhances salt stress tolerance in rice. Front. Plant Sci. 2017, 8, 316. [Google Scholar] [CrossRef] [PubMed]
- Fu, R.; Zhang, M.; Zhao, Y.; He, X.; Ding, C.; Wang, S.; Feng, Y.; Song, X.; Li, P.; Wang, B. Identification of salt tolerance-related microRNAs and their targets in Maize (Zea mays L.) using high-throughput sequencing and degradome analysis. Front. Plant Sci. 2017, 8, 864. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Polle, A. Salinity tolerance of Populus. Plant Biol. 2010, 12, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Hawighorst, P.; Sun, J.; Polle, A. Salt tolerance in Populus: Significance of stress signaling networks, mycorrhization, and soil amendments for cellular and whole-plant nutrition. Environ. Exp. Bot. 2014, 107, 113–124. [Google Scholar] [CrossRef]
- Kumar, M.; Lee, S.C.; Kim, J.Y.; Kim, S.J.; Aye, S.S.; Kim, S.R. Over-expression of dehydrin gene, OsDhn1, improves drought and salt stress tolerance through scavenging of reactive oxygen species in rice (Oryza sativa L.). J. Plant Biol. 2014, 57, 383–393. [Google Scholar] [CrossRef]
- Azzam, R.A. Polymeric conditioner gels for desert soils. Commun. Soil Sci. Plant Anal. 1983, 14, 739–760. [Google Scholar] [CrossRef]
- Woodhouse, J.; Johnson, M.S. Effect of superabsorbent polymers on survival and growth of crop seedlings. Agric. Water Manag. 1991, 20, 63–70. [Google Scholar] [CrossRef]
- Allahdadi, I.; Moazzen-Ghamsari, B.; Akbari, G.A.; Zohorianfar, M.J. Investigating the Effect of Different Rates of Superabsorbent Polymer (Superab A200) and Irrigation on the Growth and Yield of Zea Mays, 3rd Specialized Training Course and Seminar on the Application of Superabsorbent Hydrogels in Agriculture; IPPI: Tehran, Iran, 2005; pp. 52–56. [Google Scholar]
- Al-Humaid, A.I.; Moftah, A.E. Effects of hydrophilic polymers on the survival of buttonwood seedlings grown under drought stress. J. Plant Nutr. 2007, 30, 53–66. [Google Scholar] [CrossRef]
- Guiwei, Q.; Varennes, A.D.; Cunha-Queda, C. Remediation of a mine soil with insoluble polyacrylate polymers enhances soil quality and plant growth. Soil Use Manag. 2008, 24, 350–356. [Google Scholar] [CrossRef]
- El-Hady, O.A.; El-Kader, A.A.; Shafi, A.M. Physico-bio-chemical properties of sandy soil conditioned with acrylamide hydrogels after cucumber plantation. Aust. J. Basic Appl. Sci. 2009, 3, 3145–3151. [Google Scholar]
- Jobin, P.; Caron, J.; Bernier, P.Y.; Dansereau, B. Impact of two hydrophilic acrylic-based polymers on the physical properties of three substrates and the growth of Petunia hybrida ‘Brilliant Pink’. J. Am. Soc. Hort. Sci. 2004, 129, 449–457. [Google Scholar]
- Hüttermann, A.; Orikiriza, L.J.B.; Agaba, H. Application of superabsorbent polymers for improving the ecological chemistry of degraded or polluted lands. Clean-Soil Air Water 2009, 37, 517–526. [Google Scholar] [CrossRef]
- Apostol, K.G.; Jacobs, D.F.; Dumroese, R.K. Root desiccation and drought stress responses of bare root Quercus rubra seedlings treated with a hydrophilic polymer root dip. Plant Soil 2009, 315, 229–240. [Google Scholar] [CrossRef]
- Abedi-Koupai, J.; Sohrab, F. Evaluating the application of superabsorbent polymers on soil water capacity and potential on three soil textures. Iran. J. Polym. Sci. Technol. 2004, 17, 163–173. [Google Scholar]
- Guterres, J.; Rossato, L.; Pudmenzky, A.; Doley, D.; Whittaker, M.; Schmidt, S. Micron-size metal-binding hydrogel particles improve germination and radicle elongation of Australian metallophyte grasses in mine waste rock and tailings. J. Hazard. Mater. 2013, 248–249, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, A.K.; Shainberg, I.; Goldstein, D.; Warrington, D.N.; Levy, G.J. Water retention and hydraulic of cross-linked polyacrylamides in sandy soils. Soil Sci. Soc. Am. J. 2007, 71, 406–412. [Google Scholar] [CrossRef]
- Agaba, H.; Orikiriza, L.J.B.; Obua, J.; Kabasa, J.D.; Worbes, M.; Hüttermann, A. Hydrogel amendment to sandy soil reduces irrigation frequency and improves the biomass of Agrostis stolonifera. Agric. Sci. 2011, 2, 544–550. [Google Scholar] [CrossRef]
- El-Sayed, H.; Kirkwood, R.C.; Graham, N.B. The effects of a hydrogel polymer on the growth of certain horticultural crops under saline conditions. J. Exp. Bot. 1991, 42, 891–899. [Google Scholar] [CrossRef]
- Szmidt, R.A.K.; Graham, N.B. The effect of poly (ethylene oxide) hydrogel on crop growth under saline conditions. Acta Hortic. 1991, 287, 211–218. [Google Scholar] [CrossRef]
- Chen, S.; Zommorodi, M.; Fritz, E.; Wang, S.; Hüttermann, A. Hydrogel modified uptake of salt ions and calcium in Populus euphratica under saline conditions. Trees 2004, 18, 175–183. [Google Scholar]
- Shao, J.; Chen, S.; Wang, R.; Zhang, X. Enhancement of hydrogel on salt resistance of Populus popularis ‘35–44′ and its mechanism. J. Beijing For. Univ. 2007, 29, 79–84. (In Chinese) [Google Scholar]
- Shi, Y.; Li, J.; Shao, J.; Deng, S.; Wang, R.; Li, N.; Sun, J.; Zhang, H.; Zhu, H.; Zhang, Y.; et al. Effects of Stockosorb and Luquasorb polymers on salt and drought tolerance of Populus popularis. Sci. Hortic. 2010, 124, 268–273. [Google Scholar] [CrossRef]
- Dorraji, S.S.; Golchin, A.; Ahmadi, S. The effects of hydrophilic polymer and soil salinity on corn growth in sandy and loamy soils. Clean-Soil Air Water 2010, 38, 584–591. [Google Scholar] [CrossRef]
- Koroskenyi, B.; Mccarthy, S.P. Synthesis of acetylated konjac glucomannan and effect of degree of acetylation on water absorbency. Biomacromolecules 2001, 2, 824–826. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, J.; Yang, F. Konjac glucomannan, a promising polysaccharide for OCDDS. Carbohyd. Polym. 2014, 104, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Li, J.; Wang, S.; Fritz, E.; Hüttermann, A.; Altman, A. Effects of NaCl on shoot growth, transpiration, ion compartmentation, and transport in regenerated plants of Populus euphratica and Populus tomentosa. Can. J. For. Res. 2003, 33, 967–975. [Google Scholar] [CrossRef]
- Wang, R.; Chen, S.; Deng, L.; Fritz, E.; Hüttermann, A.; Polle, A. Leaf photosynthesis, fluorescence response to salinity and the relevance to chloroplast salt compartmentation and anti-oxidative stress in two poplars. Trees 2007, 21, 581–591. [Google Scholar] [CrossRef]
- Storey, R. Salt tolerance, ion relations and the effects of root medium on the response of citrus to salinity. Aust. J. Plant Physiol. 1995, 22, 101–114. [Google Scholar] [CrossRef]
- Chen, S.; Li, J.; Wang, S.; Hüttermann, A.; Altman, A. Salt, nutrient uptake and transport and ABA of Populus euphratica; a hybrid in response to increasing soil NaCl. Trees 2001, 15, 186–194. [Google Scholar] [CrossRef]
- Syvertsen, J.P.; Dunlop, J.M. Hydrophilic gel amendments to sand soil can increase growth and nitrogen uptake efficiency of citrus seedlings. Hortic. Sci. 2004, 39, 267–271. [Google Scholar]
- Hüttermann, A.; Zommorodi, M.; Reise, K. Addition of hydrogels to soil for prolonging the survival of Pinus halepensis seedlings subjected to drought. Soil Till. Res. 1999, 50, 295–304. [Google Scholar] [CrossRef]
- Arbona, V.; Iglesias, D.J.; Jacas, J.; Primo-Millo, E.; Talon, M.; Gómez-Cadenas, A. Hydrogel substrate amendment alleviates drought effects on young citrus plants. Plant Soil 2005, 270, 73–82. [Google Scholar] [CrossRef]
- Chirino, E.; Vilagrosa, A.; Vallejo, V.R. Using hydrogel and clay to improve the water status of seedlings for dryland restoration. Plant Soil 2011, 344, 99–110. [Google Scholar] [CrossRef]
- Narjary, B.; Aggarwal, P.; Singh, A.; Chakraborty, D.; Singh, R. Water availability in different soils in relation to hydrogel application. Geoderma 2012, 187–188, 94–101. [Google Scholar] [CrossRef]
- Tatirat, O.; Charoenrein, S. Physicochemical properties of konjac glucomannan extracted from konjac flour by a simple centrifugation process. LWT-Food Sci. Technol. 2011, 44, 2059–2063. [Google Scholar] [CrossRef]
- Chen, S.; Li, J.; Fritz, E.; Wang, S.; Hüttermann, A. Sodium and chloride distribution in roots and transport in three poplar genotypes under increasing NaCl stress. For. Ecol. Manag. 2002, 168, 217–230. [Google Scholar] [CrossRef]
- Chen, S.; Li, J.; Wang, T.; Wang, S.; Polle, A.; Hüttermann, A. Osmotic stress and ion-specific effects on xylem abscisic acid and the relevance to salinity tolerance in poplar. J. Plant Growth Regul. 2002, 21, 224–233. [Google Scholar] [CrossRef]
- Sun, J.; Chen, S.; Dai, S.; Wang, R.; Li, N.; Shen, X.; Zhou, X.; Lu, C.; Zheng, X.; Hu, Z.; et al. NaCl-induced alternations of cellular and tissue ion fluxes in roots of salt-resistant and salt-sensitive poplar species. Plant Physiol. 2009, 149, 1141–1153. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Dai, S.; Wang, R.; Chen, S.; Li, N.; Zhou, X.; Lu, C.; Shen, X.; Zheng, X.; Hu, Z.; et al. Calcium mediates root K+/Na+ homeostasis in poplar species differing in salt tolerance. Tree Physiol. 2009, 29, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wang, M.; Ding, M.; Deng, S.; Liu, M.; Lu, C.; Zhou, X.; Shen, X.; Zheng, X.; Zhang, Z.; et al. H2O2 and cytosolic Ca2+ signals triggered by the PM H+-coupled transport system mediate K+/Na+ homeostasis in NaCl-stressed Populus euphratica cells. Plant Cell Environ. 2010, 33, 943–958. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Han, G.; Jiang, Y.; Liu, W. Research on keep water properties of Konjac Powder. Acad. Period. Farm Prod. Process 2005, 42, 16–18. (In Chinese) [Google Scholar]
- Shabala, S.; Cuin, T.A. Potassium transport and plant salt tolerance. Physiol. Plant. 2008, 133, 651–669. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Ann. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Non-Polymer | Stockosorb | Luquasorb | KGM |
|---|---|---|---|---|
| Control | NI | NI | NI | NI |
| NaCl | 26 ± 2 | NI | 36 ± 2 | NI |
| Drought | 17 ± 1 | 34 ± 1 | 30 ± 2 | 46 ± 1 |
| Drought + NaCl | 16 ± 1 | 34 ± 2 | 30 ± 2 | 46 ± 1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Ma, X.; Sa, G.; Zhou, D.; Zheng, X.; Zhou, X.; Lu, C.; Lin, S.; Zhao, R.; Chen, S. Natural and Synthetic Hydrophilic Polymers Enhance Salt and Drought Tolerance of Metasequoia glyptostroboides Hu and W.C.Cheng Seedlings. Forests 2018, 9, 643. https://doi.org/10.3390/f9100643
Li J, Ma X, Sa G, Zhou D, Zheng X, Zhou X, Lu C, Lin S, Zhao R, Chen S. Natural and Synthetic Hydrophilic Polymers Enhance Salt and Drought Tolerance of Metasequoia glyptostroboides Hu and W.C.Cheng Seedlings. Forests. 2018; 9(10):643. https://doi.org/10.3390/f9100643
Chicago/Turabian StyleLi, Jing, Xujun Ma, Gang Sa, Dazhai Zhou, Xiaojiang Zheng, Xiaoyang Zhou, Cunfu Lu, Shanzhi Lin, Rui Zhao, and Shaoliang Chen. 2018. "Natural and Synthetic Hydrophilic Polymers Enhance Salt and Drought Tolerance of Metasequoia glyptostroboides Hu and W.C.Cheng Seedlings" Forests 9, no. 10: 643. https://doi.org/10.3390/f9100643