Changes of Scots Pine Phyllosphere and Soil Fungal Communities during Outbreaks of Defoliating Insects


1
Soil Science of Temperate and Boreal Ecosystems, Georg-August University of Goettingen, Goettingen 37077, Germany
2
Molecular Phytopathology and Mycotoxin Research, Georg-August University of Goettingen, Goettingen 37077, Germany
*
Authors to whom correspondence should be addressed.
Forests 2017, 8(9), 316; https://doi.org/10.3390/f8090316
Submission received: 14 July 2017
/
Revised: 14 August 2017
/
Accepted: 26 August 2017
/
Published: 28 August 2017
Abstract
:Outbreaks of forest pests increase with climate change, and thereby may affect microbial communities and ecosystem functioning. We investigated the structure of phyllosphere and soil microbial communities during defoliation by the nun moth (Lymantria monacha L.) (80% defoliation) and the pine tree lappet (Dendrolimus pini L.) (50% defoliation) in Scots pine forests (Pinus sylvestris L.) in Germany. Ribosomal RNA genes of fungi and bacteria were amplified by polymerase chain reaction (PCR), separated by denaturing gradient gel electrophoresis (DGGE), and subsequently sequenced for taxonomic assignments. Defoliation by both pests changed the structure of the dominant fungal (but not bacterial) taxa of the phyllosphere and the soil. The highly abundant ectomycorrhizal fungal taxon (Russula sp.) in soils declined, which may be attributed to insufficient carbohydrate supply by the host trees and increased root mortality. In contrast, potentially pathogenic fungal taxa in the phyllosphere increased during pest outbreaks. Our results suggest that defoliation of pines by insect pest, change the structure of fungal communities, and thereby indirectly may be contributing to aggravation of tree health.
1. Introduction
Over the last decades, biotic disturbances like insect outbreaks increased dramatically worldwide [1]. There is evidence that increasing intensity, frequency, duration, and extent of insect outbreaks are closely related to climate and global change [1,2]. Pest insects like the nun moth (Lymantria monacha L.) can evidently benefit from these changing environmental conditions [3]. For example, in Brandenburg, eastern Germany, about 5800 ha of forest were infested with nun moth, including 366 ha of completely defoliated area in 2013, and more than 11,000 ha of forest had to be treated with insecticides [4]. Besides substantial economic losses and expenditure for forest protection, outbreaks exhibit tremendous impact on forest ecosystem functions such as carbon sink potential and net ecosystem production [5,6,7].
The impacts of forest pest outbreaks on microbial community are not well understood. In recent years, our knowledge of the composition, diversity, and function of phyllosphere microbial communities expanded by the use of culture-independent methods like denaturing gradient gel electrophoresis (DGGE) [8]. Next generation sequencing (NGS) technologies offer the highest sampling depth, allowing the detection of low-abundant taxa [9], but the costs for the analysis of large numbers of sample sets are prohibitive. DGGE enables the monitoring of dominant taxa in a large number of samples at a fraction of the costs of NGS. The technique has been successfully used to characterize microbial communities of various environments (e.g., [10,11,12,13,14,15]). In a pioneering study by Yang et al. [15], DGGE revealed that microbial populations inhabiting the phyllosphere are much more complex than previously assumed. Studies on honeydew-excreting aphids suggest that C-rich insect excreta promote the growth of phyllosphere bacteria, filamentous fungi, and yeasts [16,17]. Beside excreta, the growth of microorganisms inhabiting the phyllosphere is enhanced by nutrient leaching from leaves damaged by folivorous insects [18,19,20,21].
During an outbreak of pine tree lappet, C and N canopy-to-soil fluxes as well as the C/N ratio in the throughfall solution in Scots pine forests in northern Germany increased significantly [22]. Total net C and N canopy-to-soil fluxes of sessile oak (Quercus petraea L.) forests during infestation with winter moth (Operophtera brumata L.) and mottled umber (Erannis defoliaria L.) were 30- and 18-fold higher compared to uninfested forests [23]. Some studies show stimulation of the soil microbial activity accompanied by alterations in soil nutrient cycling during insect outbreaks [22,23]. For example, increased input of easily biodegradable organic matter with a narrow C/N ratio during pine beauty (Panolis flammea D.) and pine tree lappet infestations led to changes in the soil bacterial community of a Scots pine forest in south-western Poland [24]. In addition to matter input, canopy damage by insect defoliation can affect belowground microbial communities, particularly microorganisms that form symbiosis with damaged trees. Soil fungal communities are generally expected to decline in abundance and richness during outbreak events, particularly those of tree symbiotic ectomycorrhizal (EM) fungi [25,26], while saprophytic and endophytic fungi species may benefit [26].
The effects of outbreaks of single pests either on phyllosphere or soil microbial communities have been studied already (e.g., [21,24,27,28,29,30,31,32,33,34,35]), but these data did not simultaneously compare the effect of pest outbreaks on phyllosphere and soil microbial communities and did not differentiate between the effects of different pest insects in the same forest ecosystem. Here, we analyzed the effect of outbreaks of two defoliating insects, nun moth or pine tree lappet, in Scots pine stands in eastern Germany on the bacterial and fungal community structure in the soil and the phyllosphere using DGGE. We hypothesize that defoliation of pine trees by pest insects would reduce the abundance of tree symbiotic microorganisms (e.g., rhizobacteria and mycorrhizal fungi), and that bacterial and fungal communities of soils and the phyllosphere would respond differentially to defoliation.
2. Materials and Methods
2.1. Study Design and Site Characteristics
The bacterial and fungal community structure of phyllosphere and soil was investigated on infested and adjacent, uninfested control forest sites in 2014 and 2015. All sites were located in the federal state of Brandenburg, Germany. In 2014, a nun moth infested site (80% defoliation) and control site were located in a 65-year-old Scots pine stand located 3 km north of Märkisch Buchholz (52°8′38″ N, 13°45′14″ E, 42 m a.s.l), Germany, and 10 km west of Teupitz (52°9′29″ N, 13°36′47″ E, 35 m a.s.l.), Germany, respectively. In 2015, the infested (52°17′42″ N, 12°19′32″ E, 47 m a.s.l.) and control (52°17′19″ N, 12°20′46″ E, 50 m a.s.l.) sites were located in a 92-year-old scot pine stand in Ziesar. The infested site showed abundant infestation (50% defoliation) with pine tree lappet. All soil types at all sites in both years were classified as podzols with a pH (1:10 in H2O) ranging from 3.2 to 3.9 in the Ah horizon and a fine to medium sand of glacial origin as parent material. The annual mean temperature was 10.8 and 10.6 °C and the total average annual precipitation was 474 and 496 mm (DWD: German Federal Meteorological Service (DWD) and Climate Data Center (CDC), Weather Station Lindenberg (ID:3015), 16.02.2017) for the sites in 2014 and 2015, respectively.
Three plots with comparable site conditions were selected for sampling, designated plot I–III for the infested and plot IV–VI for the control site. To represent the phyllosphere bacterial and fungal community in the canopy, samples of one to three-year-old pine needles were taken from five individual branches with a minimum length of 20 cm in the south exposed tree crown at 11 m height (average tree height was 13 m with the tree crown starting at approximately 9 m) of three individual trees per site plot. We collected all needles of sampled branches to achieve a representative sample. On infested sites, particularly damaged branches were sampled, whereas on the control sites branches without visible insect damage were sampled. Needle samples of each site plot were pooled after DNA extraction to obtain one mixed sample per site plot. Two replicate soil samples of 200 g fresh weight were taken from each of four soil organic layers (L Ol, Of, Oh) as well as the mineral soil horizon (Ah). After DNA extraction the replicates were pooled to obtain one mixed sample per layer and horizon at each site plot. Since the forest floor vegetation was exclusively characterized by mosses and lichens, soil samples were not sieved and tree roots were not removed so as to include tree root associated microorganisms. Further, soil samples were taken at similar distances from the trees to ensure a similar proportion of root biomass in all samples. Immediately after sampling, soil samples were dried at 40 °C for 24 h and ground; needle samples were freeze-dried for 72 h and ground.
Related to the developmental stages of the insect pests, samples were taken in early May (pre-defoliation), late May (main defoliation), and early October (post-defoliation and tree recovery period), whereas needle samples in 2014 were only available for late May and early October.
2.2. DNA Extraction from Soil and Needles
Total DNA extraction was performed using a cetyltrimethyl ammonium bromide (CTAB) extraction protocol with polyethylene glycol precipitation [36] downscaled to 50 mg material followed by phenol extraction [37]. Finely ground soil or needles were suspended in 1 mL of CTAB buffer (20 mM Na2EDTA, 10 mM Tris, 0.13 M sorbitol, 0.03 M N-laurylsarcosine, 0.02 M hexadecyltrimethylammonium bromide, 0.8 M NaCl, 1% (v/v) polyvinylpolypyrolidone, adjusted to pH 8.0 with NaOH), 1 μL 2-Mercaptoethanol, and 1 μL of Proteinase K solution (20 mg/mL) were added. After gentle vortexing, samples were incubated at 42 °C for 10 min and subsequently at 65 °C for 10 min, inverting the tubes frequently. Following adding 800 μL of chloroform/isoamylalcohol (24:1 (v/v)), the mixture was shaken, incubated on ice for 10 min, and centrifuged at 7380× g for 10 min. We mixed 700 μL of the supernatant thoroughly with 700 μL phenol and centrifuged at 7380× g for 10 min. The supernatant (650 μL) was extracted with the same amount of chloroform/isoamylalcohol (24:1 (v/v)) in the same manner. Following this, 600 μL of the liquid phase were transferred to a new Eppendorf tube containing 200 μL of polyethylene glycol (PEG) 6000 (30% (w/v)) and 100 μL of NaCl (5 M). After mixing the content thoroughly, the tubes were incubated at room temperature for 20 min and centrifuged at 16,000× g for 15 min to obtain DNA pellets. The supernatant was discarded and the pellet was washed with 500 μL of 70% (v/v) ethanol twice. The remaining ethanol in the tubes was evaporated under vacuum, the pellets were resuspended in 50 μL TE buffer (10 mM Tris, 1 mM Na2EDTA, adjusted to pH 8.0 with HCl) and incubated at 42 °C for 2 h to facilitate dissolving the DNA.
The concentration and quality of DNA was assessed by agarose gel electrophoresis. A volume of 5 μL DNA solution were mixed with 2 μL loading buffer (100 mM EDTA, 50% (v/v) glycerol, 0.025% (v/v) bromphenol blue) and loaded into a 0.8% agarose gel in 1× TAE buffer (40 mM Tris, 20 mM sodium acetate, 1 mM Na2EDTA, adjusted to pH 7.6)). Gel electrophoresis was carried out at 4.6 V/cm for 60 min, gels were stained with ethidium bromide solution (1 mg/L (w/v)) for 12 min and rinsed with double distilled water for 12 min before visualizing DNA by fluorescence in UV light. Replicate samples from each site plot were pooled for each sampling date and stored at −20 °C until analysis.
2.3. PCR-DGGE
Polymerase chain reaction (PCR) was performed in a peqSTAR 96 universal gradient thermocycler (PEQLAB, Erlangen, Germany) using 1:100 dilutions of the pooled DNA samples in a total reaction volume of 15 μL.
Bacterial 16S rDNA gene fragments were amplified using the primer set F968-GC and R1401-1a that included the hypervariable regions V6–V8 [14]. PCR mixtures were composed of 16 mM (NH4)2SO4, 67 mM Tris-HCI, 0.01% Tween-20, pH 8.8 (NH4 reaction buffer, Bioline, Luckenwalde, Germany), 3 mM MgCl2, 200 μM of each deoxyribonucleoside triphosphate, 0.8 μM of each primer, 1 mg/mL bovine serum albumin (BSA), 0.25 u Biotaq DNA-polymerase (Bioline, Luckenwalde, Germany) and 1 μL template DNA solution. PCR conditions were: initial denaturation for 5 min at 95 °C; 10 touchdown cycles of 60 s denaturation at 95 °C, 60 s annealing beginning at 60 °C with a decrease of annealing temperature by 0.5 °C per cycle, and 2 min extension at 72 °C; 30 cycles consisting of 60 s at 95 °C, 60 s at 55 °C, and 2 min at 72 °C; and final extension for 30 min at 72 °C.
Fungal 18S rDNA gene fragments were amplified using the primer set FR1-GC and FF390 that included hypervariable regions V7 and V8 [38]. The composition of PCR mixture was identical with PCR conditions for 16S rDNA except for 1.5 mM MgCl2, 0.5 μM of each primer, and 0.05 u Biotaq DNA-polymerase (Bioline, Luckenwalde, Germany). The thermocycle program consisted of an initial denaturation for 3 min at 95 °C; 10 touchdown cycles of 30 s at 95 °C, 45 s annealing beginning at 55 °C with a decrease by 0.5 °C per cycle, and 2 min at 72 °C; 30 cycles of 30 s at 95 °C, 45 s at 50 °C, and 2 min at 72 °C; and final extension for 10 min at 72 °C.
Amplified bacterial 16S rDNA and fungal 18S rDNA gene fragments were separated on a 7.5% (w/v) acrylamide/bisacrylamide gel (37:5:1) gel with a 30–70% denaturing gradient (100% denaturant correspond to 7 M urea, 40% (v/v) formamide) in TAE running buffer (40 mM Tris, 20 mM sodium acetate, 1 mM Na2EDTA, adjusted to pH 7.4 with glacial acetic acid). Gel electrophoresis was carried out using an INGENY phorU-2 system (Ingeny, International BV, Goes, The Netherlands) at 70 V (2.6 V/cm) for 16 h at 58 °C and 60 °C for fungal 18S rDNA and bacterial 16S rDNA gene fragments, respectively. PCR products were inspected on 1.7% (w/v) agarose electrophoresis gels.
2.4. Silver Staining and Gel Drying
A slightly modified protocol of the ProMega (Madison, USA) SILVER SEQUENCE™ DNA silver staining protocol [39] was used. All of the following steps were performed under continuous gentle shaking in plastic trays and all solutions were prepared with double distilled water at 4–10 °C. The use of deionized (or demineralized) water instead of doubly-distilled water for staining DGGE gels led to in-gel precipitation and a high background, which was most likely caused by organic traces leaking from the ion exchanger. To avoid these effects, double distilled water was used for all staining steps. Glass plates were removed, gels were fixed in 10% (v/v) glacial acetic acid for 20 min, rinsed two times with double distilled water for 2 min each and stained using a 0.1% (w/v) silver nitrate, 0.06% (v/v) formaldehyde solution for 35 min. Immediately after staining, the gels were rinsed with double distilled water for 30 s and transferred to a developer solution (3% (w/v) sodium carbonate, 0.06% (v/v) formaldehyde, 0.0002% (w/v) sodium thiosulfate) and the development of the image was monitored. Once DNA bands became visible, the reaction was stopped by adding 2 volumes of 10% (v/v) glacial acetic acid. The gels were kept in the solution for 4 min and rinsed with double distilled water two times for 4 min each, soaked in 3 mL of a 3% (v/v) glycerol solution and dried in a frame between two hydrated cellophane foils at room temperature for 48 h.
2.5. Selection of Soil DGGE Gels and Profiles for Sequencing
Fungal DGGE profiles of infested and uninfested plots differed distinctly for samples from late May (main defoliation) but not for samples from early May (pre-defoliation) and October (post-defoliation). Several reasons are conceivable: high population density of pests and high defoliation rates during the main defoliation might have enhanced the effect of defoliators on the fungal communities in these samples. Therefore, we further analyzed only samples taken during main defoliation in late May. Additionally, exclusively DGGE gels of the upper soil organic layers (L and Ol) were selected for DNA sequencing since ordination analysis revealed that separation between clusters of infested and uninfested control plots decreased with the position of soil organic layers and mineral soil. Decreased fungal abundance and diversity in lower soil horizons is mainly caused by the decrease of oxygen level with soil depth, which may explain why the differences between fungal populations of infested and uninfested plots declined with soil depth [40]. DNA bands exhibiting largest differences in DGGE profiles from infested pines and uninfested controls during main defoliation in late May were selected for sequencing. When sequencing did not enable discrimination of taxa on family level, taxa were assigned to the next higher taxonomic level (order).
2.6. Band Excision, Reamplification, and Sequencing
Based on the NMDS, bands were excised from dried gels using a razor blade and placed in double distilled water for 20 s. Cellophane was removed and imbibed polyacrylamide was ground by sterile plastic cones in 50 μL double distilled water and incubated at 4 °C over night. Polyacrylamide was removed by centrifugation and supernatants were used to reamplify DNA fragments using primers identical to the primers used to generate DGGE samples but without GC-clamps; the same PCR conditions without the touchdown phase was used. PCR products were inspected on 1.7% (w/v) agarose gels; DNA was precipitated with isopropanol in a final concentration of 70% (v/v) at room temperature for 10 min and centrifuged at 16,160× g for 10 min. The pellet was washed with 70% (v/v) ethanol and dissolved in 12 μL double distilled water and send to Macrogen Europe (Macrogen Europe, Amsterdam, The Netherlands) for sequencing. To verify the identity of bands occurring at the same position on a DGGE gel but within different samples, at least two bands occurring at the same position were sequenced. The sequences were assigned to fungal and bacterial taxa as follows. Published sequences with ≥99% identity were retrieved from the GeneBank database at the National Center for Biotechnology Information (NCBI) using the software BLAST [41] with default settings. The origin of the strains from which these sequences originated and their taxonomic characterization were checked with the help of metadata attached to the accessions and sequences without reliable taxonomic assignment were discarded.
2.7. Data Analysis of DGGE Profiles
DGGE gels were placed on a light-table and photographed for visual band identification. Although the test design prevented inaccuracies in comparing profiles between two or more gels [37], gel images were first adjusted for exposure and contrast and then converted into black and white to achieve the maximum number of identifiable bands and minimize the bias of the staining intensity.
A matrix based on relative intensities of DGGE bands (0 = absent, 1 = least intense, 4 = most intense) was constructed for each gel for semiquantitative analysis [42,43]. Ordination analysis of the data was performed using non-metric multidimensional scaling ordination (NMDS) of Euclidean distance matrices. The analysis was executed using the R (Version 3.3.1 GUI 1.68 Mavericks build, R Foundation for Statistical Computing, Vienna, Austria, 2016) package vegan v.2.0-4 [44].
3. Results
3.1. Bacterial Communities
Ordination analysis as well as manual reviewing of DGGE profiles revealed no significant differences between infested and control plots in samples under insect outbreaks at any sampling date. Thus, fragments of bacterial DNA separated by DGGE were not sequenced.
3.2. Fungal Communities
3.2.1. Phyllosphere Fungi
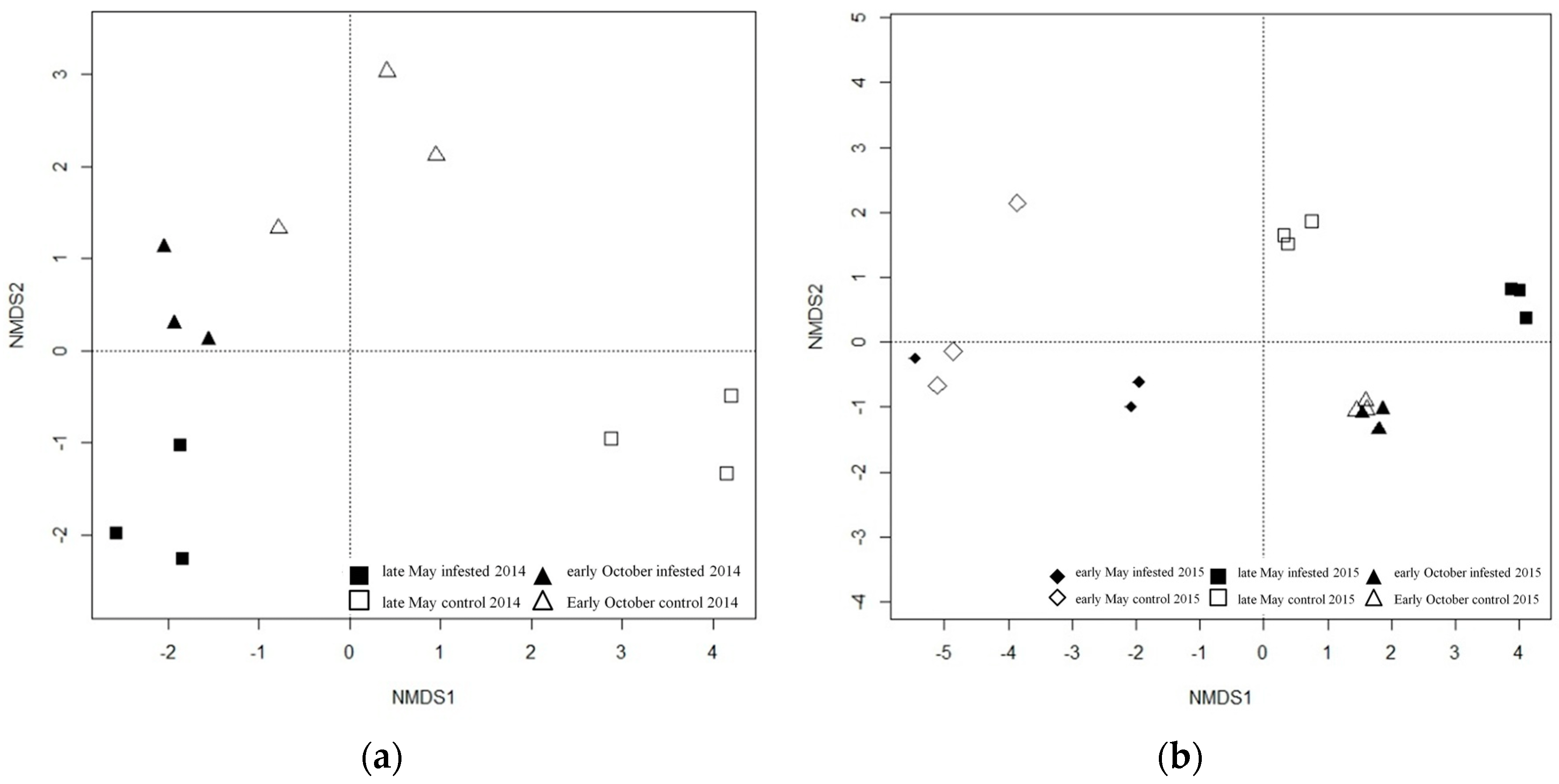
The fungal community of the phyllosphere showed well separated clustering between infested and uninfested control forest plots during the main period of defoliation in late May compared to early May and October 2014 and 2015 (Figure 1). Taxonomic assignments based on the sequences of the phyllosphere community in both years revealed that all fungi were assigned to the phylum Ascomycota (Figure 2, Table 1 and Table 2).
Products assigned to Hypocreales (family: Nectriaceae) and Dothideales (family: Dothideaceae) were present in both sample types from pine trees in 2014 but they differed in their relative intensities: the taxon assigned to Nectriaceae and one of the two taxa belonging to Dothideaceae were more abundant in uninfested control samples while the other taxon of Dothideaceae was more abundant in infested pine tree samples. One amplicon, assigned to Xylariales (family: Amphisphaeriaceae), occurred in both years in both sample types but was more abundant in the samples from infested pine trees. The majority of taxa were assigned to Capnodiales, occurring in both years with different relative intensities: in 2014 these taxa occurred either only in infested (1 taxon) or uninfested (4 taxa) phyllosphere samples; in samples taken in 2015 taxa assigned to Capnodiales (family: Mycosphaerellaceae) were either only detected in infested (2 taxa) or uninfested control samples (1 taxon) but also exhibited different abundance between infested and uninfested samples (5 taxa). Both PCR products assigned to Pleosporales (family: Pleosporaceae) occurred only in the samples of infested phyllosphere from 2015.
3.2.2. Soil Fungal Community
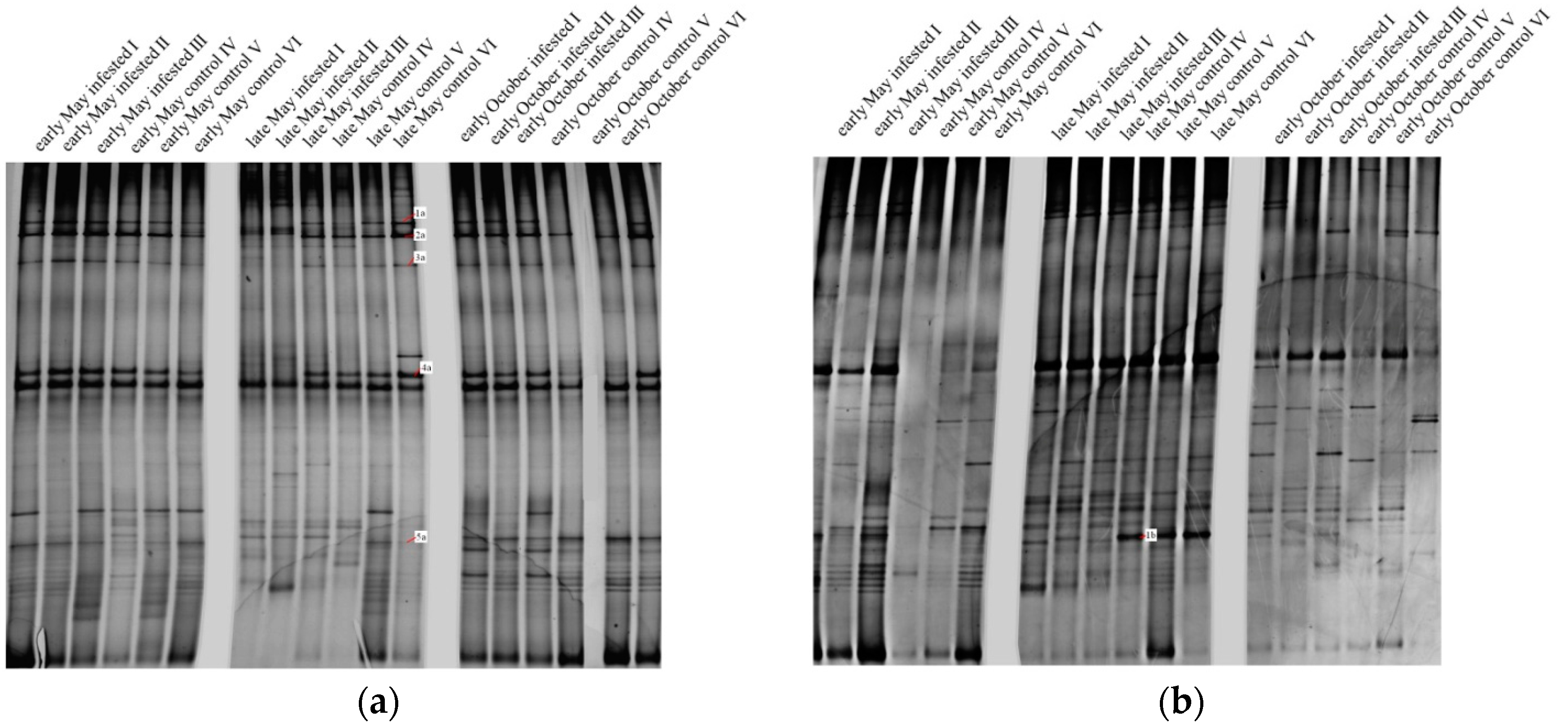
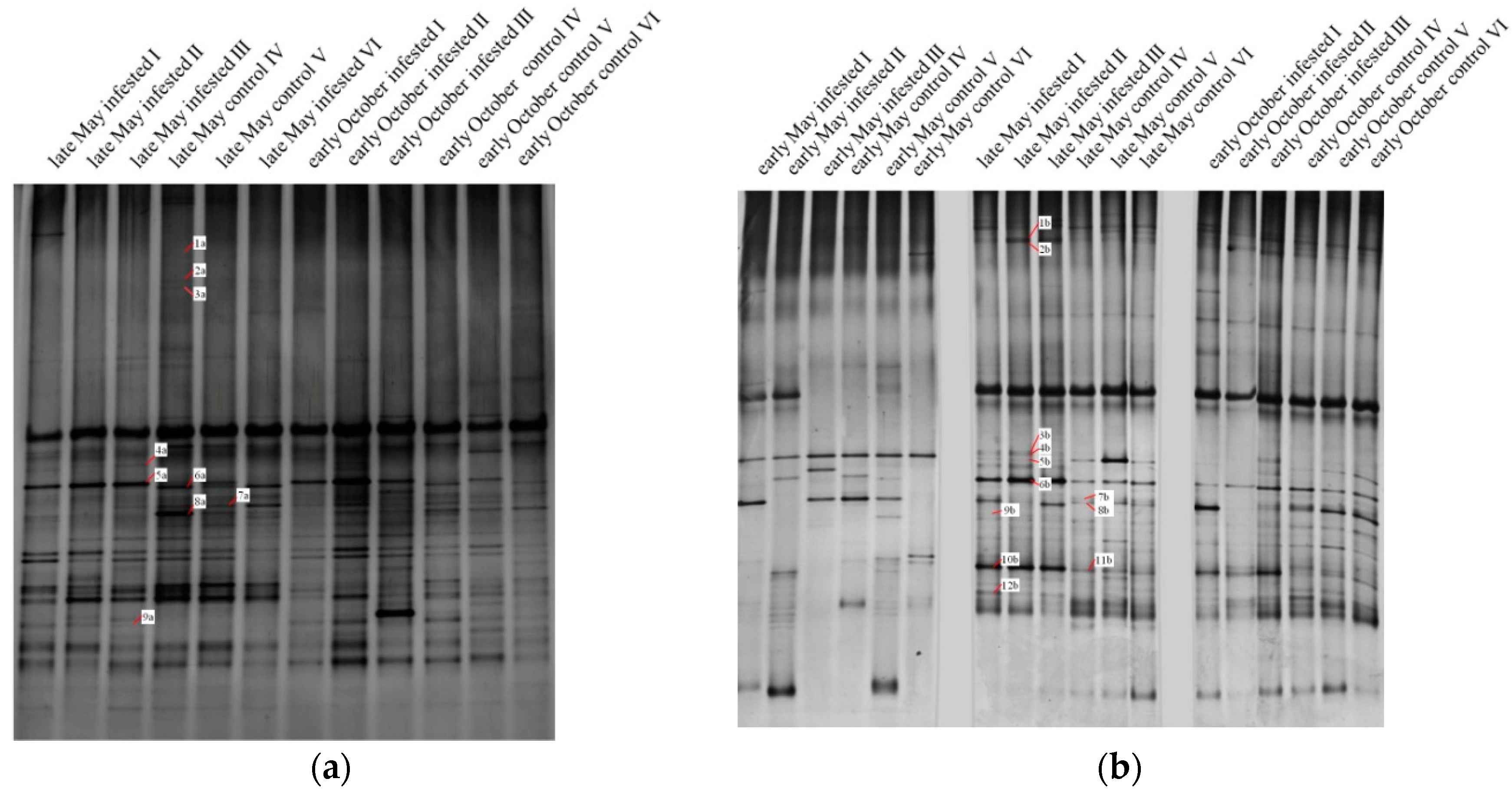
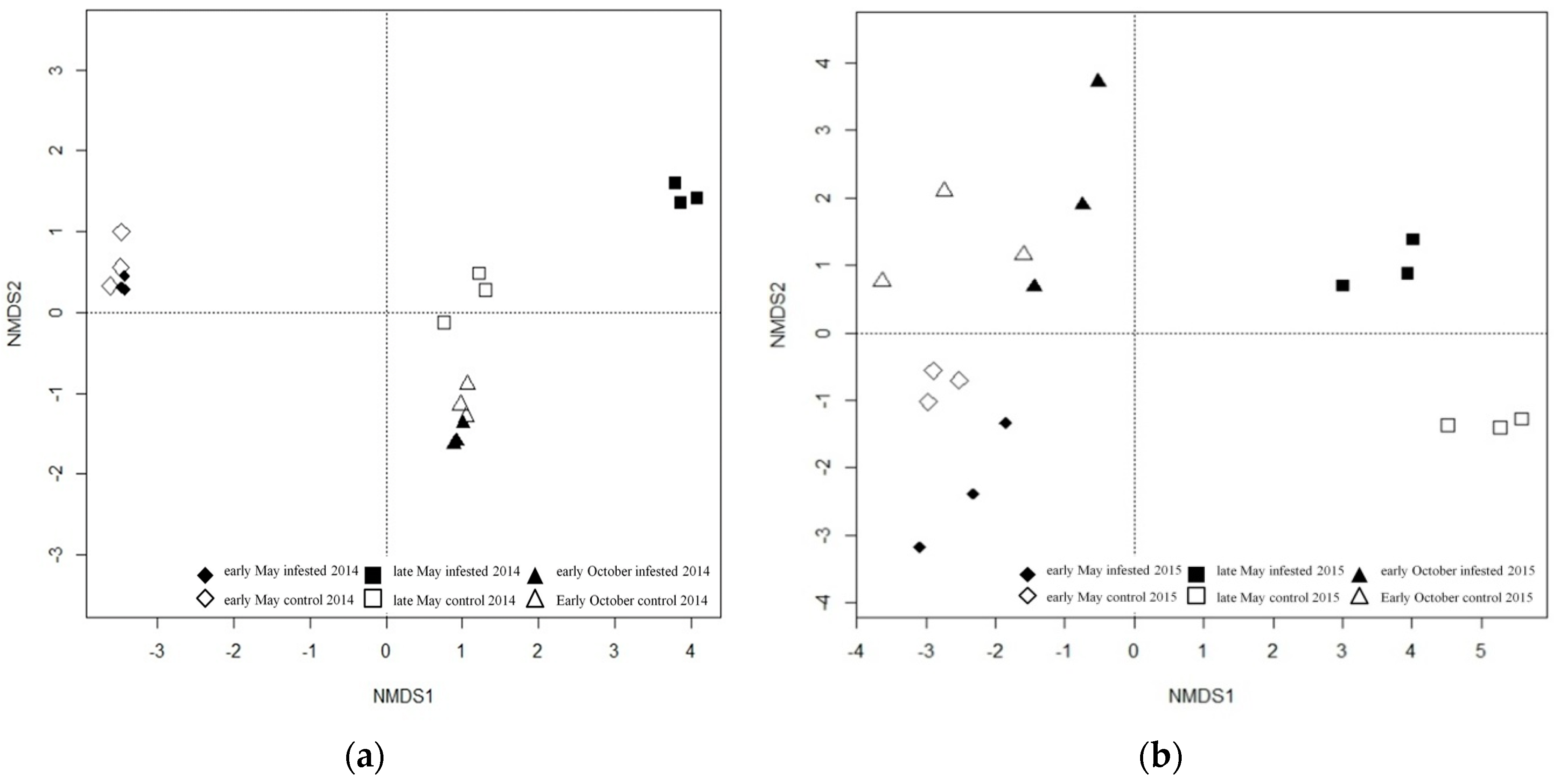
In accordance with the fungal community of the phyllosphere, the soil fungal community showed separation between clusters of infested and uninfested forest plots in late May compared to early May and October 2014 and 2015 (Figure 3). Cluster separation between infested and uninfested plots during the main defoliation in late May decreased with increasing soil depth (data not shown). Therefore, only DNA bands exhibiting the largest differences in DGGE profiles of L and Ol layers in late May were selected for sequencing (Figure 4); the L layer was analyzed using samples from 2014 and the Ol layer using samples from 2015.
A taxon, assigned to Trichocomaceae, was only present in two of the three samples of the uninfested control plots in 2014 (Table 3). Taxa assignable to the genus Rhizomucor (family: Mucoraceae) accounted for the majority of sequenced bands in 2014 with varying relative intensities between infested and uninfested control samples: one of the three infested samples exhibited the same relative abundance pattern of Rhizomucor as the uninfested control samples which all show occurrences of these taxa; in the other two soil samples from the infested site only one of the four Rhizomucor taxa is present. One amplicon present in DGGE profiles of samples taken in 2015 was assigned to the genus Russula (family: Russulaceae) (Table 4). This taxon was only detected in uninfested plots, despite a relatively low abundance in one of the three infested plots.
4. Discussion
The analyses of bacterial and fungal community of soils and phyllospheres during insect pest outbreaks in Scots pine forests showed that defoliation changed the structure of dominant fungal communities of the phyllosphere and the soil, while bacterial communities were not affected. Moreover, infestation of pine trees with pests in the present study stimulated the colonization of needles with potential parasites and pathogens. Our results indicate that insect damages are accompanied by alterations in dominating fungal species. However, it should be noted that our analysis provides a picture that is limited by observation time, site conditions, as well as tree and insect species.
4.1. Bacterial Community of Pine Needles and Soil
Infestation of pine trees with foliar feeding pests neither affected the bacterial communities of needles nor soils in our study. Nonetheless, results documented in the literature on effects of insect damage on bacteria colonizing the phyllosphere remain controversial. It is known that defoliating insects contribute to the dispersal of bacteria on the phyllosphere [45]. Studies on honeydew-excreting aphids showed that C-rich insect excreta significantly increased the growth of phyllosphere bacteria, filamentous fungi and yeasts [16,17]. For example, heavy infestations of eastern hemlock (Tsuga canadensis L.) with hemlock wooly adelgid (Adelges tsugae A.) strongly stimulated bacteria, filamentous fungi, and yeasts in the phyllosphere [46]. A study conducted by Müller et al. [21] investigated the effects of lepidopterous larvae infesting European beech (Fagus sylvatica L.) and sessile oak on culturable phyllosphere bacteria. Defoliation of sessile oak increased the proportion of bacteria utilizing mineral N [21]. Growth-promoting effects on culturable bacteria, filamentous fungi, and yeasts of the phyllosphere were also observed on European beech and sessile oak during moth infestation [20]. Thus, changes in bacterial community structures in the phyllosphere appear to be related to species-specific insect pest-tree interactions.
In accordance with other studies, soil bacterial community structure appeared to be relatively stable with no changes under insect infestation. Similarly, culture-independent pyrosequencing in a 5-year investigation of bark beetle (Ips typographus L.) induced mortality of limber pines (Pinus flexilis E.) did not reveal substantial changes in the bacterial community of soil during outbreaks [27]. In contrast, altered soil bacterial community structure was found to be coupled with altered soil edaphic properties under heavily (85% tree mortality) bark beetle-impacted lodgepole pine (Pinus contorta) trees [33]. Further, outbreaks of pine beauty and pine tree lappet on Scots pine increased the abundance of culturable actinobacteria, which was attributed to litterfall of easily biodegradable organic matter with a narrow C/N ratio [24]. An increase of N content in needles as a compensatory response to defoliation was often observed [47,48,49]. Needles with a higher N content, which get into the soil via litterfall, stimulate decomposition later in the season and are introduced into soil in increased rates during pest outbreaks [50]. In addition, the extent of infestation was found to determine the biogeochemical responses as well as the soil bacterial communities [33,35]. Mikkelson et al. [34] elegantly highlighted the importance of rare bacterial taxa on the community dynamics and presumably biogeochemical cycling under bark beetle-induced tree mortality. Because DGGE visualizes only amplicons originating from dominant species, the effect of defoliation on rare taxa might have remained undetected, although they might be critical in maintaining ecosystem functioning.
4.2. Fungal Community of Pine Needles
Pine needle fungi indicated considerable changes in community structure, with a high abundance of Ascomycota in the phyllosphere during outbreaks (cf. [30,31]). The phyllosphere of Norway spruce (Picea abies L.) stands following a spruce bud scale outbreak (Physokermes piceae S.) in Lithuania showed also high abundances of plant pathogenic fungi (Rhizosphaera kalkhoffii, Exobasidium bisporum, Phialophora sessilis) [30]. These are known to cause damage to plant tissues. Therefore, alteration of fungal communities during pest outbreaks, additionally may threat tree health [30]. In our study, a taxon assigned to the family Amphisphaeriaceae occurred in all plots with higher abundance in the infested plots. Many Amphisphaeriaceae are opportunistic pathogens colonizing damaged trees in temperate zones [51,52]. One taxon of the family Dothideaceae occurred only in infested plots, while another Dothideaceae taxon was predominantly found in uninfested plots. Dothideaceae are cosmopolitans that can be endophytic, biotrophic, necrothropic or saprobic on various plants [53]. Some taxa (e.g., Scirrhia pini F. & P.) are known to cause economically relevant damage to conifers [54,55]. A taxon of the family Nectriaceae was less abundant during insect defoliation in 2014. Taxa belonging to the family Nectriaceae are cosmopolitans and show association with plant necromass or occur as pathogens of other fungi [56,57]. Two taxa assigned to the family Pleosporaceae were only detected in infested plots. The occurrence was related to outbreaks of the pine tree lappet. Pleosporaceae include necrotrophs and saprophytes [58,59], including economical important pathogens that can threaten tree health. Fungal taxa of the family Mycosphaerellaceae varied in response to insect infestation. Mycosphaerellaceae exhibit as diverse lifestyles as Dothideaceae [60], and may have contributed to the different abundances.
4.3. Soil Fungal Community
Soil fungal community structures responded significantly to insect outbreaks. The abundance of several taxa assigned to the genera Rhizomucor and the family Trichocomaceae was distinctly reduced in infested plots. Rhizomucor and Trichocomaceae are saprophytic and saprotrophic fungi, respectively, with global distribution in soils and decomposing vegetation [61,62,63]. The reduced abundance of saprophytic and saprotrophic fungi in soils from infested Scots pine plots stands is a new finding, and contrasts earlier studies (e.g., [26,28]) on Norway spruce and subarctic mountain birch (Betula pubescens ssp. czerepanovii H.-A.). Apparently, species-specific interactions and/or the biogeographical effects related to composition region appear to contribute to structuring of these fungal communities.
The abundance of an ectomycorrhizal fungus assigned to the genus Russula was strongly reduced in soil samples collected from infested trees during infestation in the L layer in 2015, supporting the hypothesis that defoliation would suppress the abundance of these fungi. This finding is in line with the results of an artificial defoliation experiment in Scots pine by Kuikka et al. [25] reporting reduced production of sporocarps as well as reduced diversity in ectomycorrhizal fungi under defoliated trees (see also [28,29,32]). For example, in lodgepole pine (Pinus contorta Dougl. ex. Loud. var. latifolia Engelm.) pine beetle (Dendroctonus ponderosae Hopkins) outbreaks led to a decline in species richness [32] as well as in the abundance of ectomycorrhizal fungi [29]. Bark beetle outbreaks in Norway spruce stands also reduced ectomycorrhizal fungi, but increased saprophytic fungi [28]. Successive defoliation of subarctic mountain birch by autumnal (Epirrita autumnata B.) and winter moth (Operophtera brumata L.) larvae in northern Finland led to a decline of the abundance and richness of ectomycorrhizal fungi by 70–80%, but benefited saprophytic and endophytic fungi [26]. Ectomycorrhizal fungi depend on the supply of carbohydrates via host roots [64,65]. It is likely that high losses of needle biomass during defoliation decrease photosynthesis and consequently carbohydrate supply of this symbiotic association. Additionally, it is known that an impaired nutritional status can cause increased root mortality and depressed growth of fine roots [66]. Therefore, reduced root biomass in soil samples of infested sites may contribute to the observed lower abundance of ectomycorrhizal fungi.
5. Conclusions
The interactions of species in the course of biotic disturbances in forests need to be reconsidered. Defoliation of Scots pine by insects changed the structure of phyllosphere and soil fungal communities but not those of bacterial communities. In particular, the highly abundant symbiotic association of an ectomycorrhizal taxon (Russula sp.) declined during an outbreak of pine tree lappet, and thereby may affect ecosystem functioning. Since pest outbreaks are short term events (2–3 years) in the life span of a forest ecosystem, observed adverse changes of fungal communities appear to occur rather rapidly, emphasizing the importance of monitoring and management of forest pest.
Acknowledgments
The authors thank Anke Sirrenberg for her dedication during troubleshooting with PCR-DGGE. In addition, Sebastian Hesse and Jonas Watterott for participating in the laboratory work and Annett Reinhardt and Franziska Germeshausen for the field work. The authors thank Carsten Thies for linguistic revision. Further, the authors thank the two anonymous reviewers for their valuable input. This study was founded by the Germany Research Foundation (DFG) (DFG Project No. ME 3861/2-1).
Author Contributions
A.l.-M.-A. and P.K. conceived and designed the experiments; L.B. and M.M.G. performed the experiments; L.B. analyzed the data; L.B. and M.M.G. wrote the manuscript; P.K. revised the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Van Lierop, P.; Lindquist, E.; Sathyapala, S.; Franceschini, G. Global forest area disturbance from fire, insect pests, diseases and severe weather events. For. Ecol. Manag. 2015, 352, 78–88. [Google Scholar] [CrossRef]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J.; et al. Climate change and forest disturbances. BioScience 2001, 51, 723–734. [Google Scholar] [CrossRef]
- Majunke, C.; Möller, K.; Funke, M. Die Nonne. Waldschutz-Merkblatt 52, 3rd ed.; Umweltschutz und Raumordnung des Landes Brandenburg; Landesforstanstalt Eberswalde und Ministerium für Landwirtschaft: Eberswalde, Germany, 2008; p. 10.
- Möller, K.; Heydeck, P. Situationsbericht zum Auftreten von Schaderregern und Schäden im Land Brandenburg; Aktuelle Waldschutzsituation 07/2013; Ministerium für Infrastruktur und Landwirtschaft des Landes Brandenburg: Eberswalde, Germany, 2013; p. 3.
- Langström, B.; Annila, E.; Hellqvist, C.; Varama, M.; Niemelä, P. Tree mortality, needle biomass recovery and growth losses in Scots pine following defoliation by Diprion pini (L.) and subsequent attack by Tomicus piniperda (L.). Scand. J. For. Res. 2001, 16, 342–353. [Google Scholar] [CrossRef]
- Armour, H.; Straw, N.; Day, K. Interactions between growth, herbivory and long-term foliar dynamics of Scots pine. Trees Struct. Funct. 2003, 17, 70–80. [Google Scholar] [CrossRef]
- Hicke, J.A.; Allen, C.D.; Desai, A.R.; Dietze, M.C.; Hall, R.J.; Kashian, D.M.; Moore, D.; Raffa, K.F.; Sturrock, R.N.; Vogelmann, J. Effects of biotic disturbances on forest carbon cycling in the United States and Canada. Glob. Chang. Biol. 2012, 18, 7–34. [Google Scholar] [CrossRef]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Knief, C. Analysis of plant microbe interactions in the era of next generation sequencing technologies. Front. Plant Sci. 2014, 5, 216. [Google Scholar] [CrossRef] [PubMed]
- Nakatsu, C.H.; Torsvik, V.; Øvreås, L. Soil community analysis using DGGE of 16S rDNA polymerase chain reaction products. Soil Sci. Soc. Am. J. 2000, 64, 1382–1388. [Google Scholar] [CrossRef]
- Dı́ez, B.; Pedrós-Alió, C.; Marsh, T.L.; Massana, R. Application of denaturing gradient gel electrophoresis (DGGE) to study the diversity of marine picoeukaryotic assemblages and comparison of DGGE with other molecular techniques. Appl. Environ. Microbiol. 2001, 67, 2942–2951. [Google Scholar] [CrossRef] [PubMed]
- González-Toril, E.; Llobet-Brossa, E.; Casamayor, E.O.; Amann, R.; Amils, R. Microbial ecology of an extreme acidic environment, the Tinto River. Appl. Environ. Microbiol. 2003, 69, 4853–4865. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Y.; He, L.M.; Wu, J.; Jiang, Q. Bacterial community diversity associated with four marine sponges from the South China Sea based on 16S rDNA-DGGE fingerprinting. J. Exp. Mar. Biol. Ecol. 2006, 329, 75–85. [Google Scholar] [CrossRef]
- Brons, J.K.; Van Elsas, J.D. Analysis of bacterial communities in soil by use of denaturing gradient gel electrophoresis and clone libraries, as influenced by different reverse primers. Appl. Environ. Microbiol. 2008, 74, 2717–2727. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.H.; Crowley, D.E.; Borneman, J.; Keen, N.T. Microbial phyllosphere populations are more complex than previously realized. Proc. Natl. Acad. Sci. USA 2001, 98, 3889–3894. [Google Scholar] [CrossRef] [PubMed]
- Dik, A.J.; van Pelt, J.A. Interaction between phyllosphere yeasts, aphid honeydew and fungicide effectiveness in wheat under field conditions. Plant Pathol. 1992, 41, 661–675. [Google Scholar] [CrossRef]
- Stadler, B.; Müller, T. Aphid honeydew and its effect on the phyllosphere microflora of Picea abies (L.) Karst. Oecologia 1996, 108, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Kimmins, J.P. Relative contributions of leaching, litter-fall and defoliation by Neodiprion sertifer (Hymenoptera) to the removal of cesium-134 from red pine. Oikos 1972, 226–234. [Google Scholar] [CrossRef]
- Schowalter, T.D.; Hargrove, W.; Crossley, D.A., Jr. Herbivory in forested ecosystems. Ann. Rev. Entomol. 1986, 31, 177–196. [Google Scholar] [CrossRef]
- Stadler, B.; Müller, T. Effects of aphids and moth caterpillars on epiphytic microorganisms in canopies of forest trees. Can. J. For. Res. 2000, 30, 631–638. [Google Scholar] [CrossRef]
- Müller, T.; Müller, M.; Behrendt, U.; Stadler, B. Diversity of culturable phyllosphere bacteria on beech and oak: The effects of lepidopterous larvae. Microbiol. Res. 2003, 158, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Le Mellec, A.; Habermann, M.; Michalzik, B. Canopy herbivory altering C to N ratios and soil input patterns of different organic matter fractions in a Scots pine forest. Plant Soil 2009, 325, 255–262. [Google Scholar] [CrossRef]
- L-M-Arnold, A.; Grüning, M.; Simon, J.; Reinhardt, A.B.; Lamersdorf, N.; Thies, C. Forest defoliator pests alter carbon and nitrogen cycles. R. Soc. Open Sci. 2016, 3, 160361. [Google Scholar] [CrossRef] [PubMed]
- Stremińska, M.A.; Blaszczyk, M.; Kolk, A. Microbial Abundance and Some of Their Physiological Activities in Soil Organic Horizon of Pine Forest Affected by Insect Herbivory. Pol. J. Environ. Stud. 2006, 15, 905–914. [Google Scholar]
- Kuikka, K.; Härmä, E.; Markkola, A.; Rautio, P.; Roitto, M.; Saikkonen, K.; Ahonen-Jonnarth, U.; Finlay, R.; Tuomi, J. Severe defoliation of Scots pine reduces reproductive investment by ectomycorrhizal symbionts. Ecology 2003, 84, 2051–2061. [Google Scholar] [CrossRef]
- Saravesi, K.; Aikio, S.; Wäli, P.R.; Ruotsalainen, A.L.; Kaukonen, M.; Huusko, K.; Suokas, M.; Brown, S.P.; Jumpponen, A.; Tuomi, J.; et al. Moth outbreaks alter root-associated fungal communities in subarctic mountain birch forests. Microb. Ecol. 2015, 69, 788–797. [Google Scholar] [CrossRef] [PubMed]
- Ferrenberg, S.; Knelman, J.E.; Jones, J.M.; Beals, S.C.; Bowman, W.D.; Nemergut, D.R. Soil bacterial community structure remains stable over a 5-year chronosequence of insect-induced tree mortality. Front. Microbiol. 2014, 5, 681. [Google Scholar] [CrossRef] [PubMed]
- Štursová, M.; Šnajdr, J.; Cajthaml, T.; Bárta, J.; Šantrůčková, H.; Baldrian, P. When the forest dies: The response of forest soil fungi to a bark beetle-induced tree dieback. ISME J. 2014, 8, 1920–1931. [Google Scholar] [CrossRef]
- Treu, R.; Karst, J.; Randall, M.; Pec, G.J.; Cigan, P.W.; Simard, S.W.; Cooke, J.E.K.; Erbilgin, N.; Cahill, J.F. Decline of ectomycorrhizal fungi following a mountain pine beetle epidemic. Ecology 2014, 95, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Menkis, A.; Marciulynas, A.; Gedminas, A.; Lynikien, J. High-throughput sequencing reveals drastic changes in fungal communities in the phyllosphere of Norway spruce (Picea abies) following invasion of the spruce bud scale (Physokermes piceae). Microb. Ecol. 2015, 70, 904. [Google Scholar] [CrossRef] [PubMed]
- Abdelfattah, A.; Wisniewski, M.; Nicosia, M.G.L.D.; Cacciola, S.O.; Schena, L. Metagenomic analysis of fungal diversity on strawberry plants and the effect of management practices on the fungal community structure of aerial organs. PLoS ONE 2016, 11, e0160470. [Google Scholar] [CrossRef] [PubMed]
- Pec, G.J.; Karst, J.; Taylor, D.L.; Cigan, P.W.; Erbilgin, N.; Cooke, J.E.; Simard, S.W.; Cahill, J.F. Change in soil fungal community structure driven by a decline in ectomycorrhizal fungi following a mountain pine beetle (Dendroctonus ponderosae) outbreak. New Phytol. 2017, 213, 864–873. [Google Scholar] [CrossRef] [PubMed]
- Mikkelson, K.M.; Lozupone, C.A.; Sharp, J.O. Altered edaphic parameters couple to shifts in terrestrial bacterial community structure associated with insect-induced tree mortality. Soil Biol. Biochem. 2016, 95, 19–29. [Google Scholar] [CrossRef]
- Mikkelson, K.M.; Bokman, C.M.; Sharp, J.O. Rare taxa maintain microbial diversity and contribute to terrestrial community dynamics throughout bark beetle infestation. Appl. Environ. Microbiol. 2016, 82, 6912–6919. [Google Scholar] [CrossRef] [PubMed]
- Brouillard, B.M.; Mikkelson, K.M.; Bokman, C.M.; Berryman, E.M.; Sharp, J.O. Extent of localized tree mortality influences soil biogeochemical response in a beetle-infested coniferous forest. Soil Biol. Biochem. 2017, 114, 309–318. [Google Scholar] [CrossRef]
- Brandfass, C.; Karlovsky, P. Upscaled CTAB-based DNA extraction and real-time PCR assays for Fusarium culmorum and F. graminearum DNA in plant material with reduced sampling error. Int. J. Mol. Sci. 2008, 9, 2306–2321. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Vainio, E.J.; Hantula, J. Direct analysis of wood-inhabiting fungi using denaturing gradient gel electrophoresis of amplified ribosomal DNA. Mycol. Res. 2000, 104, 927–936. [Google Scholar] [CrossRef]
- SILVER SEQUENCE™ DNA Sequencing System. Available online: https://www.promega.de/-/media/files/resources/protocols/technical-manuals/0/silver-sequence-dna-sequencing-system-protocol.pdf (accessed on 8 July 2017).
- Voříšková, J.; Brabcová, V.; Cajthaml, T.; Baldrian, P. Seasonal dynamics of fungal communities in a temperate oak forest soil. New Phytol. 2014, 201, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Basic Local Alignment Search Tool. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 8 July 2017).
- Fromin, N.; Hamelin, J.; Tarnawski, S.; Roesti, D.; Jourdain-Miserez, K.; Forestier, N.; Teyssier-Cuvelle, S.; Gillet, F.; Aragna, M.; Rossi, P. Statistical analysis of denaturing gel electrophoresis (DGE) fingerprinting patterns. Environ. Microbiol. 2002, 4, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Fry, J.C.; Webster, G.; Cragg, B.A.; Weightman, A.J.; Parkes, R.J. Analysis of DGGE profiles to explore the relationship between prokaryotic community composition and biogeochemical processes in deep subseafloor sediments from the Peru Margin. FEMS Microbiol. Ecol. 2006, 58, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simspons, G.L.; Solymos, P.; Stevens, M.H.; Szoecs, E.; et al. Community Ecology Package 2013. Version 2.0-4. Available online: https://cran.ism.ac.jp/web/packages/vegan/vegan.pdf (accessed on 8 July 2017).
- Lilley, A.K.; Hails, R.S.; Cory, J.S.; Bailey, M.J. The dispersal and establishment of pseudomonad populations in the phyllosphere of sugar beet by phytophagous caterpillars. FEMS Microbiol. Ecol. 1997, 24, 151–157. [Google Scholar] [CrossRef]
- Stadler, B.; Müller, T.; Orwig, D.; Cobb, R. Hemlock woolly adelgid in New England forests: Canopy impacts transforming ecosystem processes and landscapes. Ecosystems 2005, 8, 233–247. [Google Scholar] [CrossRef]
- Gómez, S.; Orians, C.M.; Preisser, E.L. Exotic herbivores on a shared native host: Tissue quality after individual, simultaneous, and sequential attack. Oecologia 2012, 169, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- Rubino, L.; Charles, S.; Sirulnik, A.G.; Tuininga, A.R.; Lewis, J.D. Invasive insect effects on nitrogen cycling and host physiology are not tightly linked. Tree Physiol. 2015, 35, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Grüning, M.M.; Simon, J.; Rennenberg, H.; l-M-Arnold, A. Defoliating Insect Mass Outbreak Affects Soil N Fluxes and Tree N Nutrition in Scots Pine Forests. Front. Plant Sci. 2017, 8, 954. [Google Scholar] [CrossRef] [PubMed]
- Vestgarden, L.S. Carbon and nitrogen turnover in the early stage of Scots pine (Pinus sylvestris L.) needle litter decomposition: Effects of internal and external nitrogen. Soil Biol. Biochem. 2001, 33, 465–474. [Google Scholar] [CrossRef]
- Kang, J.C.; Hyde, K.D.; Kong, R.Y. Studies on Amphisphaeriales: The Amphisphaeriaceae (sensu stricto). Mycol. Res. 1999, 103, 53–64. [Google Scholar] [CrossRef]
- Jeewon, R.E.C.Y.; Liew, E.C.Y.; Hyde, K.D. Phylogenetic evaluation of species nomenclature of Pestalotiopsis in relation to host association. Fungal Divers. 2004, 17, 39–55. [Google Scholar]
- Cannon, P.F.; Carmarán, C.C.; Romero, A.I. Studies on biotrophic fungi from Argentina: Microcyclus porlieriae, with a key to South American species of Microcyclus. Mycol. Res. 1995, 99, 353–356. [Google Scholar] [CrossRef]
- Funk, A.; Parker, A.K. Scirrhia pini n. sp., the perfect state of Dothistroma pini Hulbary. Can. J. Bot. 1966, 44, 1171–1176. [Google Scholar] [CrossRef]
- Butin, H. Teleomorph-und anamorph-Entwicklung von Scirrhia pini Funk & Parker auf Nadeln von Pinus nigra Arnold. Sydowia 1985, 38, 20–27. [Google Scholar]
- Dobbeler, P. Ascospore diversity of bryophilous Hypocreales and two new hepaticolous Nectria species. Mycologia 2005, 97, 924–934. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.J.; Geiser, D.M.; Proctor, R.H.; Rooney, A.P.; O’Donnell, K.; Trail, F.; Gardiner, D.M.; Manners, J.M.; Kazan, K. Fusarium pathogenomics. Ann. Rev. Microbiol. 2013, 67, 399–416. [Google Scholar] [CrossRef] [PubMed]
- Berbee, M.L.; Pirseyedi, M.; Hubbard, S. Cochliobolus phylogenetics and the origin of known, highly virulent pathogens, inferred from ITS and glyceraldehyde-3-phosphate dehydrogenase gene sequences. Mycologia 1999, 91, 964–977. [Google Scholar] [CrossRef]
- Zhang, Y.; Schoch, C.L.; Fournier, J.; Crous, P.W.; De Gruyter, J.; Woudenberg, J.H.C.; Hirayama, K.; Tanaka, K.; Pointing, S.B.; Spatafora, J.W.; et al. Multi-locus phylogeny of Pleosporales: A taxonomic, ecological and evolutionary re-evaluation. Stud. Mycol. 2009, 64, 85–102. [Google Scholar] [CrossRef] [PubMed]
- Simon, U.K.; Groenewald, J.Z.; Crous, P.W. Cymadothea trifolii, an obligate biotrophic leaf parasite of Trifolium, belongs to Mycosphaerellaceae as shown by nuclear ribosomal DNA analyses. Persoonia 2009, 22, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Shweta, M.M. Onychomycosis due to Rhizomucor in psoriatic patient with HIV infection. Indian J. Dermatol. 2013, 58, 242. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, E.; Visser, L.; Nijssen, A.; van Veluw, G.J.; Wösten, H.A.B.; de Vries, R.P. Analysis of regulation of pentose utilisation in Aspergillus niger reveals evolutionary adaptations in Eurotiales. Stud. Mycol. 2011, 69, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Houbraken, J.; Samson, R.A. Phylogeny of Penicillium and the segregation of Trichocomaceae into three families. Stud. Mycol. 2011, 70, 1–51. [Google Scholar] [CrossRef] [PubMed]
- Nehls, U.; Grunze, N.; Willmann, M.; Reich, M.; Kuester, H. Sugar for my honey: Carbohydrate partitioning in ectomycorrhizal symbiosis. Phytochemistry 2007, 68, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Gieger, T.; Thomas, F.M. Effects of defoliation and drought stress on biomass partitioning and water relations of Quercus robur and Quercus petraea. Basic Appl. Ecol. 2002, 3, 171–181. [Google Scholar] [CrossRef]
Figure 1.
Non-metric multidimensional scaling ordination (NMDS) of Euclidean distance matrices of 18S rDNA denaturing gradient gel electrophoresis (DGGE) profiles of the phyllosphere fungal community in Scots pine (Pinus sylvestris L.) forests in 2014 (a) and 2015 (b).
Figure 1.
Non-metric multidimensional scaling ordination (NMDS) of Euclidean distance matrices of 18S rDNA denaturing gradient gel electrophoresis (DGGE) profiles of the phyllosphere fungal community in Scots pine (Pinus sylvestris L.) forests in 2014 (a) and 2015 (b).
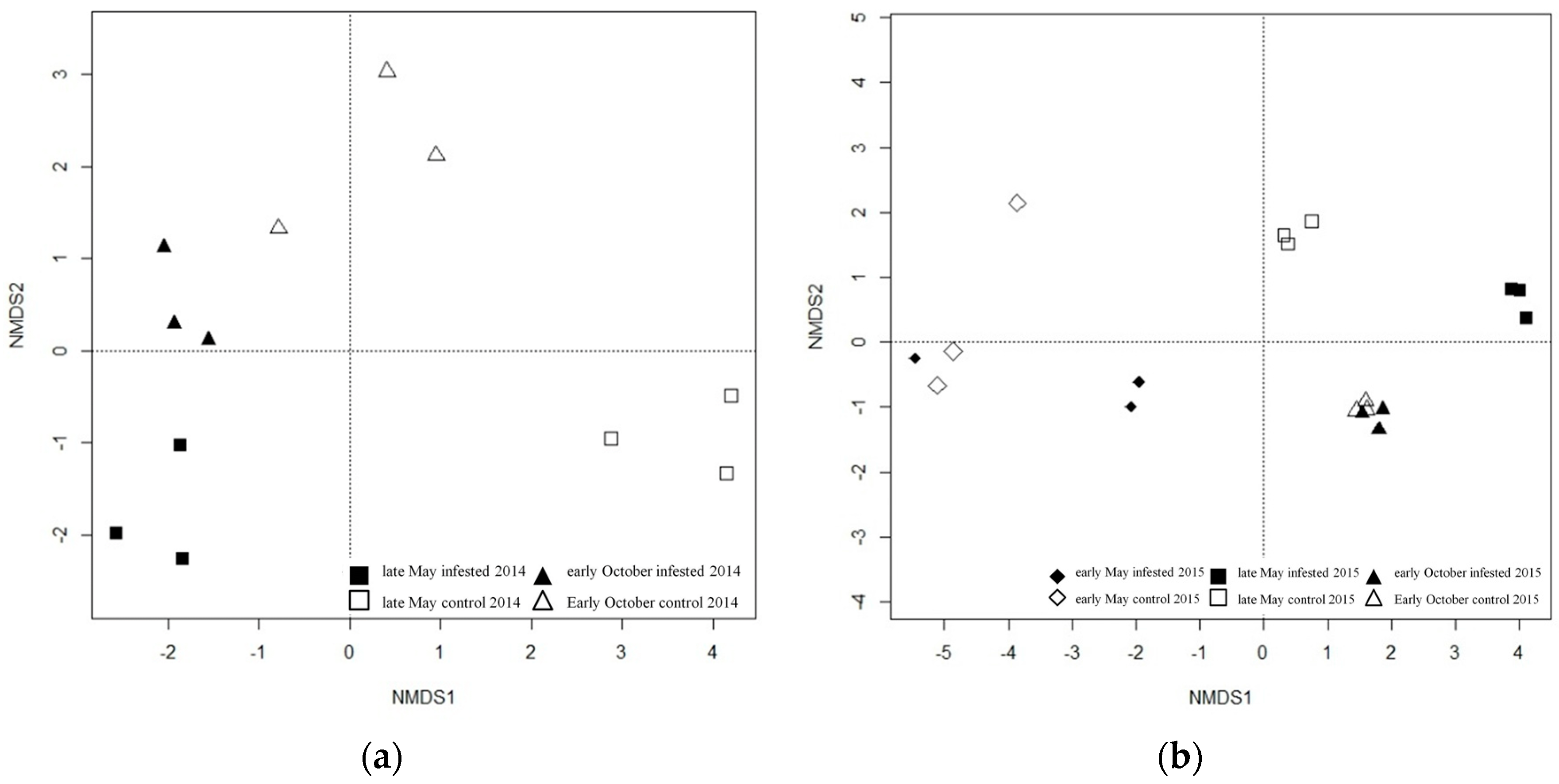
Figure 2.
Denaturing gradient gel electrophoresis (DGGE) profiles of 18S rDNA gene fragments of the phyllosphere fungal community in Scots pine (Pinus sylvestris L.) forests in 2014 (a) and 2015 (b). Labelled bands were excised for sequencing.
Figure 2.
Denaturing gradient gel electrophoresis (DGGE) profiles of 18S rDNA gene fragments of the phyllosphere fungal community in Scots pine (Pinus sylvestris L.) forests in 2014 (a) and 2015 (b). Labelled bands were excised for sequencing.
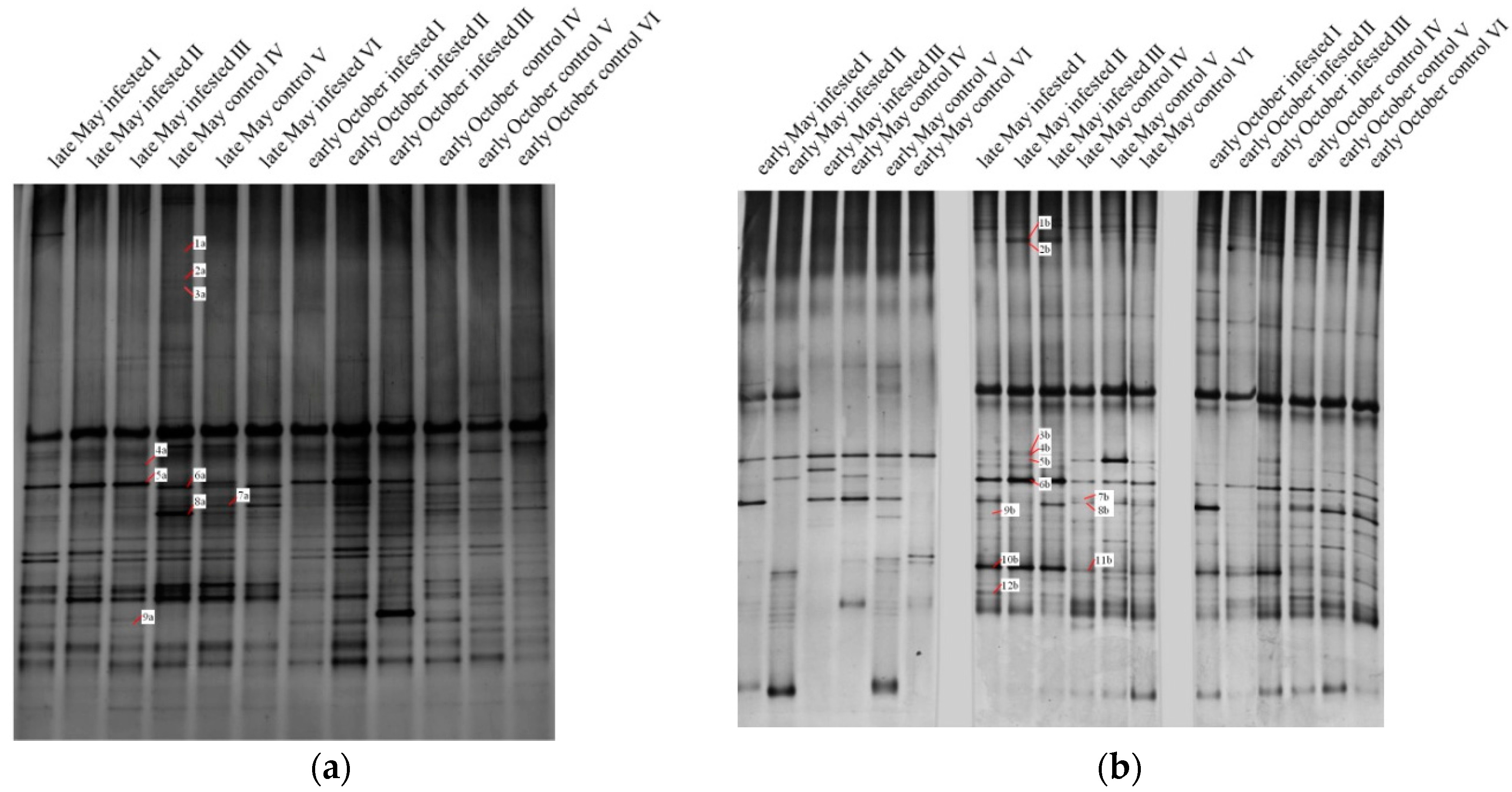
Figure 3.
Non-metric multidimensional scaling ordination (NMDS) of Euclidean distance matrices of 18S rDNA denaturing gradient gel electrophoresis (DGGE) profiles of the phyllosphere fungal community of the L soil organic layer in 2014 (a) and Ol soil organic layer in 2015 (b).
Figure 3.
Non-metric multidimensional scaling ordination (NMDS) of Euclidean distance matrices of 18S rDNA denaturing gradient gel electrophoresis (DGGE) profiles of the phyllosphere fungal community of the L soil organic layer in 2014 (a) and Ol soil organic layer in 2015 (b).
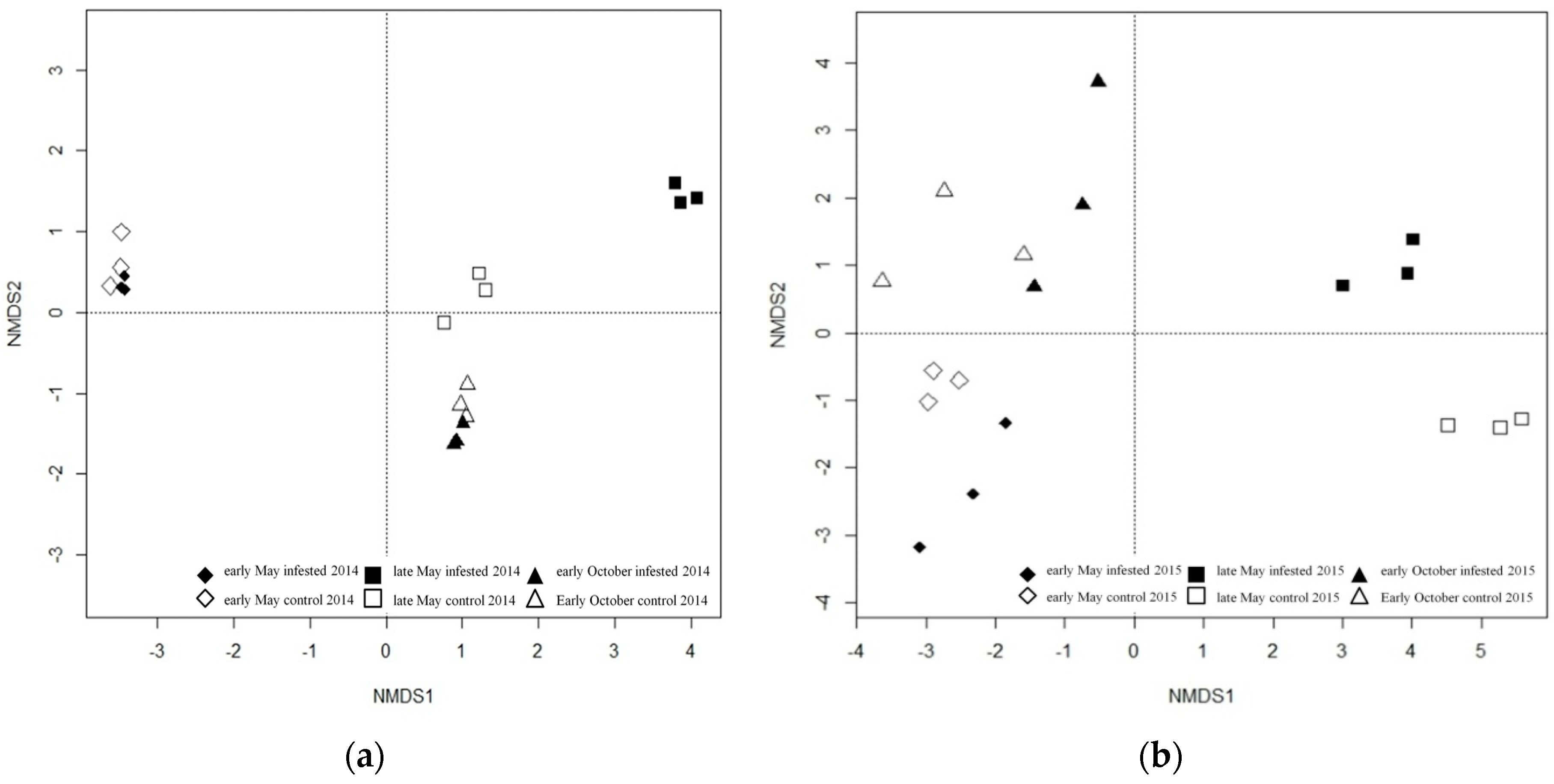
Figure 4.
Denaturing gradient gel electrophoresis (DGGE) profiles of 18S rDNA gene fragments of the soil fungal community in Scots pine (Pinus sylvestris L.) forests of the L soil organic layer in 2014 (a) and the Ol soil organic layer in 2015 (b). Labelled bands were excised for sequencing.
Figure 4.
Denaturing gradient gel electrophoresis (DGGE) profiles of 18S rDNA gene fragments of the soil fungal community in Scots pine (Pinus sylvestris L.) forests of the L soil organic layer in 2014 (a) and the Ol soil organic layer in 2015 (b). Labelled bands were excised for sequencing.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Taxonomic assignment of fungal DNA sequences obtained from the needles of Scots pine (Pinus sylvestris L.) in late May 2014. Phylum and order of the taxa are listed. Positions of the corresponding denaturing gradient gel electrophoresis (DGGE) bands are shown in Figure 2a.
Table 1.
Taxonomic assignment of fungal DNA sequences obtained from the needles of Scots pine (Pinus sylvestris L.) in late May 2014. Phylum and order of the taxa are listed. Positions of the corresponding denaturing gradient gel electrophoresis (DGGE) bands are shown in Figure 2a.
| Band | Phylum | Order | Infested Plots | Control Plots | ||||
|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | |||
| 1a | Ascomycota | Capnodiales | − | − | − | + | + | + |
| 2a | Ascomycota | Capnodiales | − | − | − | + | + | + |
| 3a | Ascomycota | Capnodiales | − | − | − | + | + | + |
| 4a | Ascomycota | Dothideales | − | + | + | − | − | − |
| 5a | Ascomycota | Xylariales | +++ | +++ | +++ | + | ++ | − |
| 6a | Ascomycota | Capnodiales | − | − | − | +++ | ++ | +++ |
| 7a | Ascomycota | Dothideales | − | − | + | ++ | ++ | +++ |
| 8a | Ascomycota | Hypocreales | + | + | − | +++ | ++ | + |
| 9a | Ascomycota | Capnodiales | + | + | + | − | − | − |
+, ++ and +++ indicate increasing relative band intensity; − indicates absence of bands.
Table 2.
Taxonomic assignment of fungal DNA sequences obtained from the needles of Scots pine (Pinus sylvestris L.) in late May 2015. Phylum and order of the taxa are listed. Positions of the corresponding denaturing gradient gel electrophoresis (DGGE) bands are shown in Figure 2b.
Table 2.
Taxonomic assignment of fungal DNA sequences obtained from the needles of Scots pine (Pinus sylvestris L.) in late May 2015. Phylum and order of the taxa are listed. Positions of the corresponding denaturing gradient gel electrophoresis (DGGE) bands are shown in Figure 2b.
| Band | Phylum | Order | Infested Plots | Control Plots | ||||
|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | |||
| 1b | Ascomycota | Capnodiales | + | ++ | ++ | + | + | + |
| 2b | Ascomycota | Capnodiales | + | ++ | ++ | + | + | + |
| 3b | Ascomycota | Capnodiales | + | + | + | − | − | − |
| 4b | Ascomycota | Capnodiales | + | + | + | − | − | − |
| 5b | Ascomycota | Capnodiales | + | + | + | ++ | +++ | ++ |
| 6b | Ascomycota | Xylariales | +++ | +++ | +++ | ++ | ++ | ++ |
| 7b | Ascomycota | Capnodiales | + | + | + | + | + | + |
| 8b | Ascomycota | Capnodiales | ++ | − | − | + | ++ | + |
| 9b | Ascomycota | Pleosporales | + | + | + | − | − | − |
| 10b | Ascomycota | Pleosporales | +++ | +++ | +++ | − | − | − |
| 11b | Ascomycota | Capnodiales | − | − | − | + | + | + |
| 12b | Ascomycota | Capnodiales | + | + | + | − | − | − |
+, ++ and +++ indicate increasing relative band intensity; − indicates absence of bands.
Table 3.
Taxonomic assignment of fungal DNA sequences obtained in the organic L layer of soil in Scots pine (Pinus sylvestris L.) forests in late May 2014. Phylum and family of the taxa are listed. Positions of the corresponding denaturing gradient gel electrophoresis (DGGE) bands are shown in Figure 4a.
Table 3.
Taxonomic assignment of fungal DNA sequences obtained in the organic L layer of soil in Scots pine (Pinus sylvestris L.) forests in late May 2014. Phylum and family of the taxa are listed. Positions of the corresponding denaturing gradient gel electrophoresis (DGGE) bands are shown in Figure 4a.
| Band | Phylum | Family | Infested Plots | Control Plots | ||||
|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | |||
| 1a | Zygomycota | Mucoraceae | − | − | ++ | ++ | ++ | +++ |
| 2a | Zygomycota | Mucoraceae | − | − | +++ | +++ | +++ | +++ |
| 3a | Zygomycota | Mucoraceae | + | + | ++ | ++ | ++ | ++ |
| 4a | Zygomycota | Mucoraceae | − | − | +++ | +++ | +++ | +++ |
| 5a | Ascomycota | Trichocomaceae | − | − | − | − | ++ | + |
+, ++ and +++ indicate increasing relative band intensity. − indicates absence of bands.
Table 4.
Taxonomic assignment of fungal DNA sequences obtained in the L layer of soil in Scots pine (Pinus sylvestris L.) forests in late May 2015 Phylum and family of the taxa are listed. Positions of the corresponding denaturing gradient gel electrophoresis (DGGE) bands are shown in Figure 4b.
Table 4.
Taxonomic assignment of fungal DNA sequences obtained in the L layer of soil in Scots pine (Pinus sylvestris L.) forests in late May 2015 Phylum and family of the taxa are listed. Positions of the corresponding denaturing gradient gel electrophoresis (DGGE) bands are shown in Figure 4b.
| Band | Phylum | Genus | Infested Plots | Control Plots | ||||
|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | |||
| 1b | Basidiomycota | Russula | − | + | − | +++ | +++ | +++ |
+, indicates low relative band intensity and +++ high relative band intensity; – indicates absence of bands.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Beule, L.; Grüning, M.M.; Karlovsky, P.; L-M-Arnold, A. Changes of Scots Pine Phyllosphere and Soil Fungal Communities during Outbreaks of Defoliating Insects. Forests 2017, 8, 316. https://doi.org/10.3390/f8090316
AMA Style
Beule L, Grüning MM, Karlovsky P, L-M-Arnold A. Changes of Scots Pine Phyllosphere and Soil Fungal Communities during Outbreaks of Defoliating Insects. Forests. 2017; 8(9):316. https://doi.org/10.3390/f8090316
Chicago/Turabian StyleBeule, Lukas, Maren Marine Grüning, Petr Karlovsky, and Anne L-M-Arnold. 2017. "Changes of Scots Pine Phyllosphere and Soil Fungal Communities during Outbreaks of Defoliating Insects" Forests 8, no. 9: 316. https://doi.org/10.3390/f8090316
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.