
On the Effect of Thinning on Tree Growth and Stand Structure of White Birch (Betula platyphylla Sukaczev) and Siberian Larch (Larix sibirica Ledeb.) in Mongolia

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
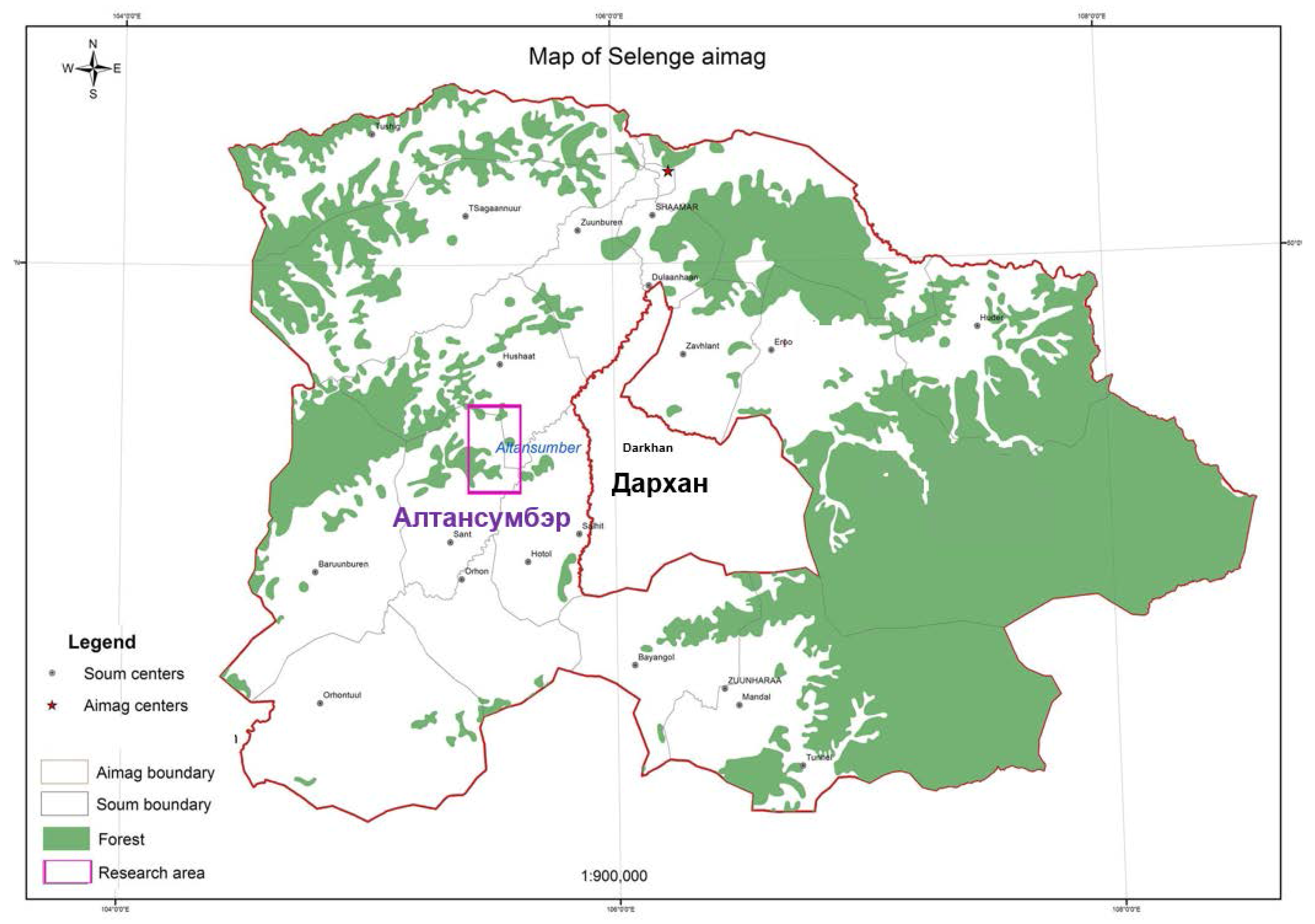
2.1. Participatory Establishment of the Thinning Trials in the research area (RA) Altansumber
2.2. Methods
2.2.1. Characterisation of Thinning Type and Intensity—Non-Spatial Harvest Event Analysis
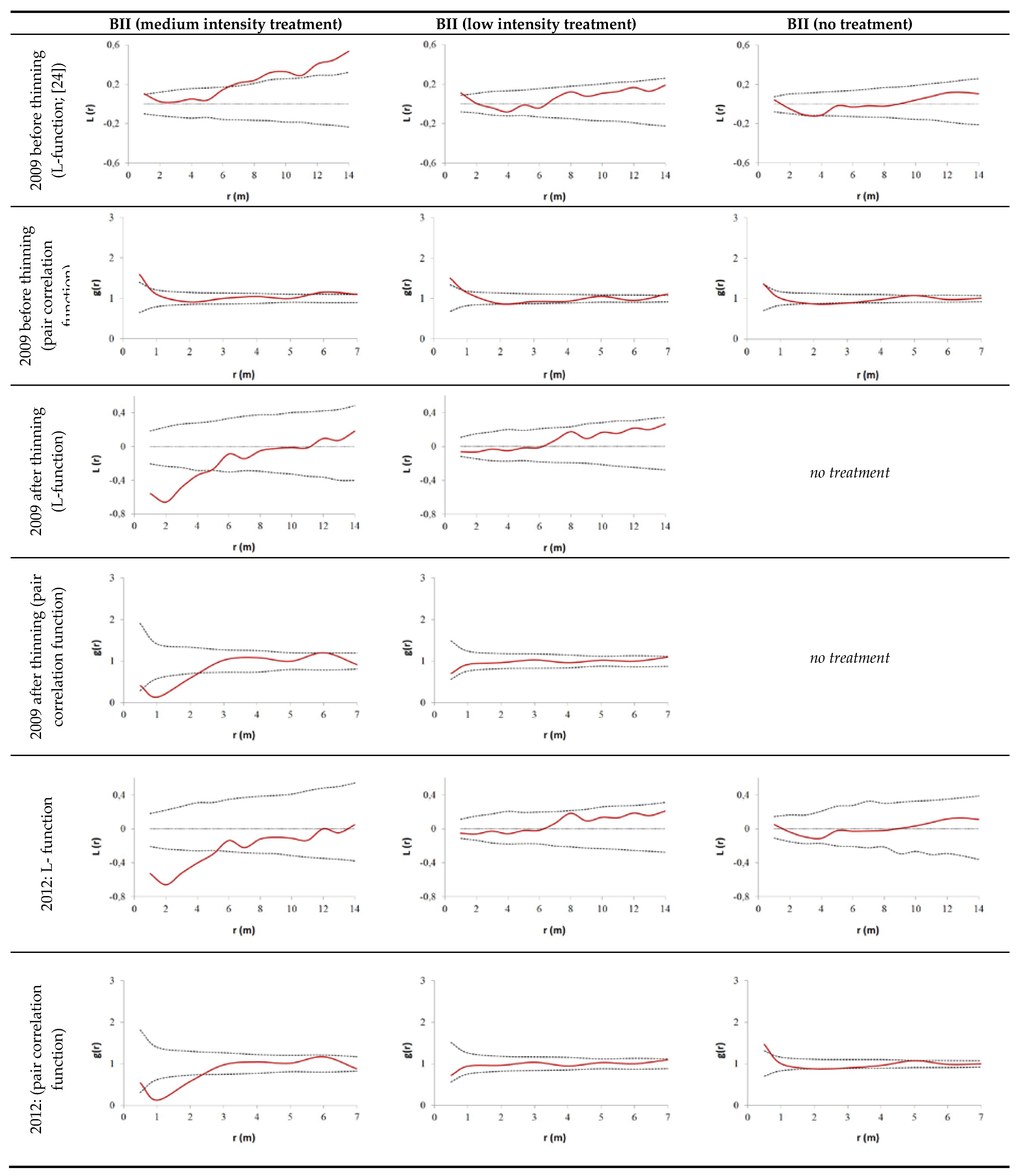
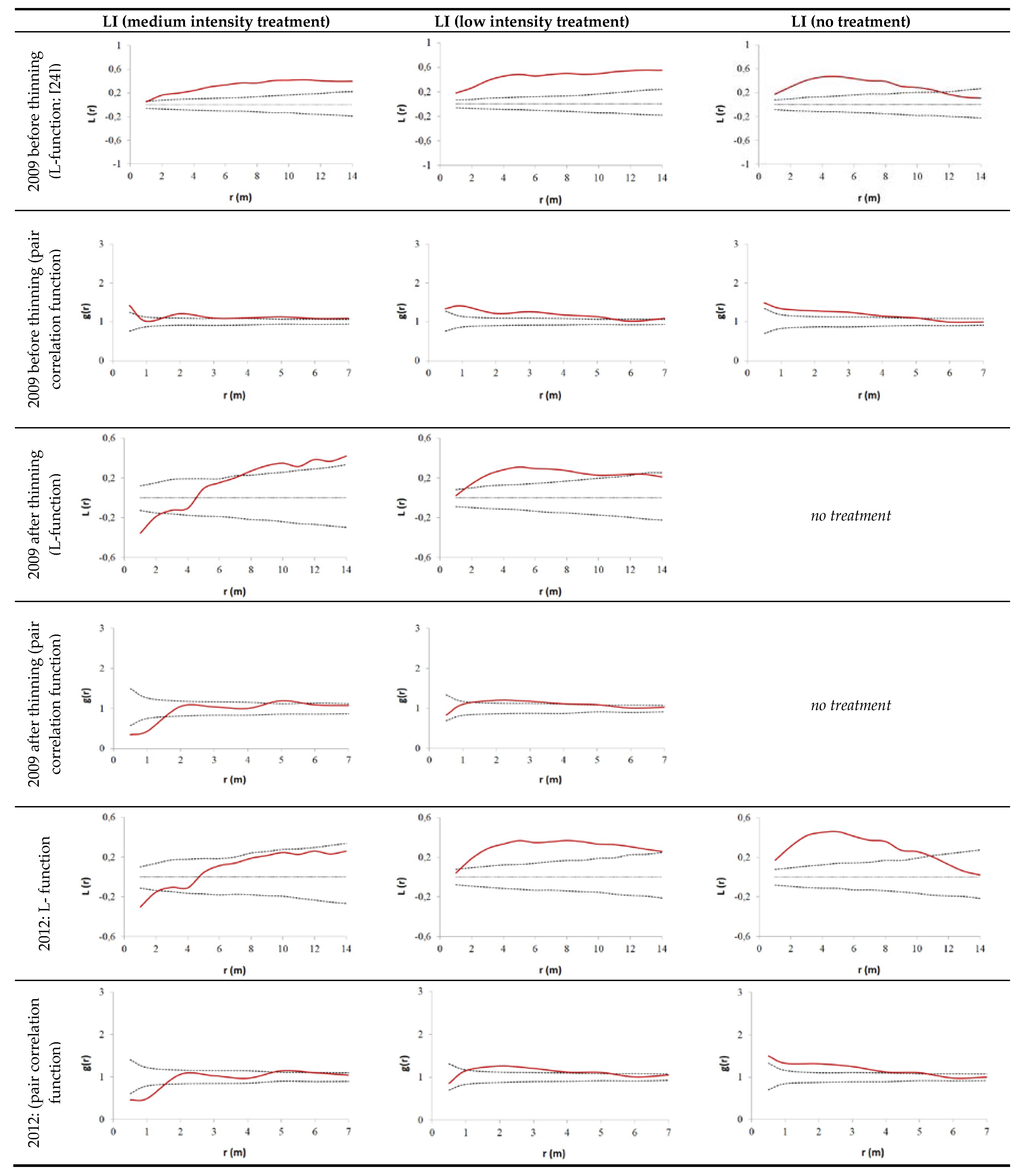
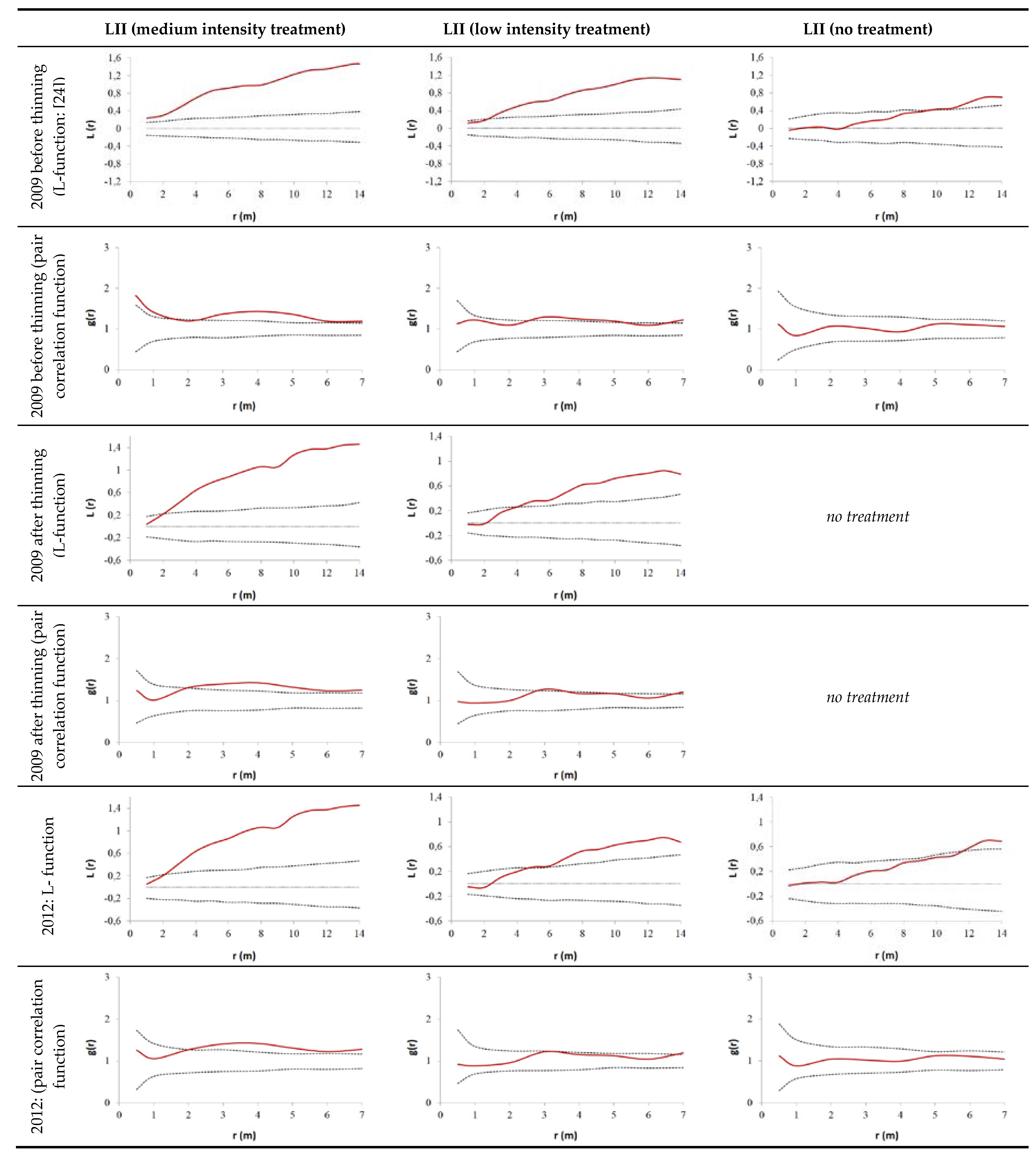
2.2.2. Evaluation of Spatial Tree Distribution on the Plots during the Observation Period
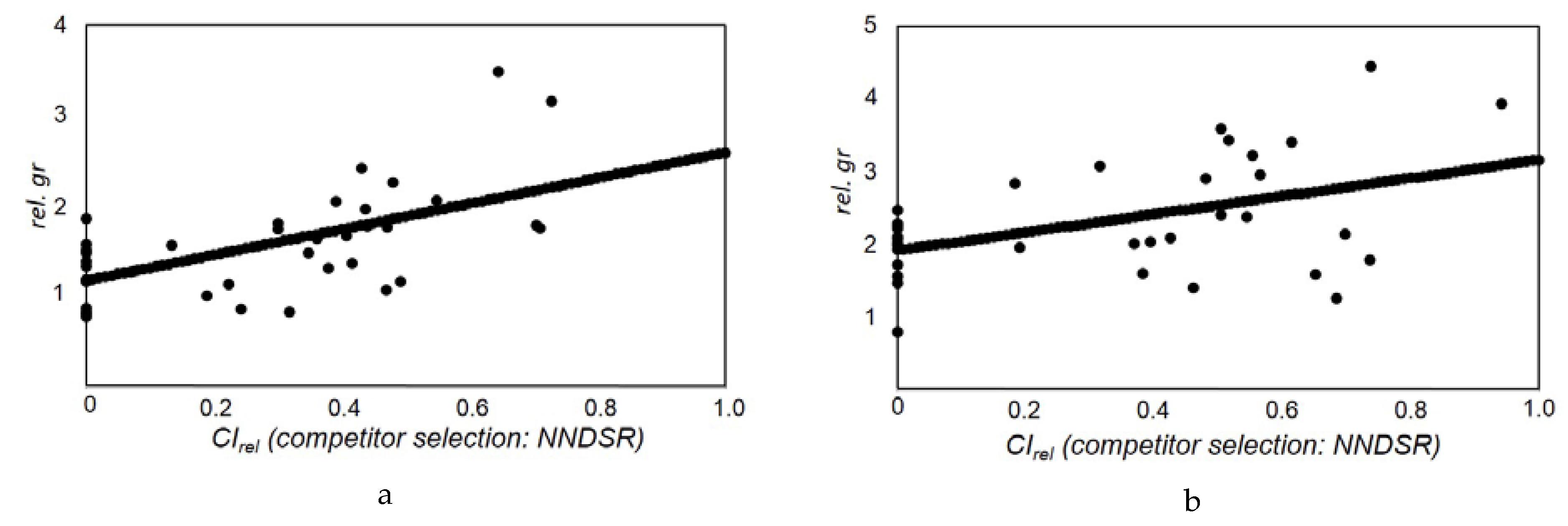
2.2.3. Evaluation of Single Tree Growth Response
3. Results
3.1. Characterisation of the Thinning Impact—Non-Spatial Harvest Event Analysis
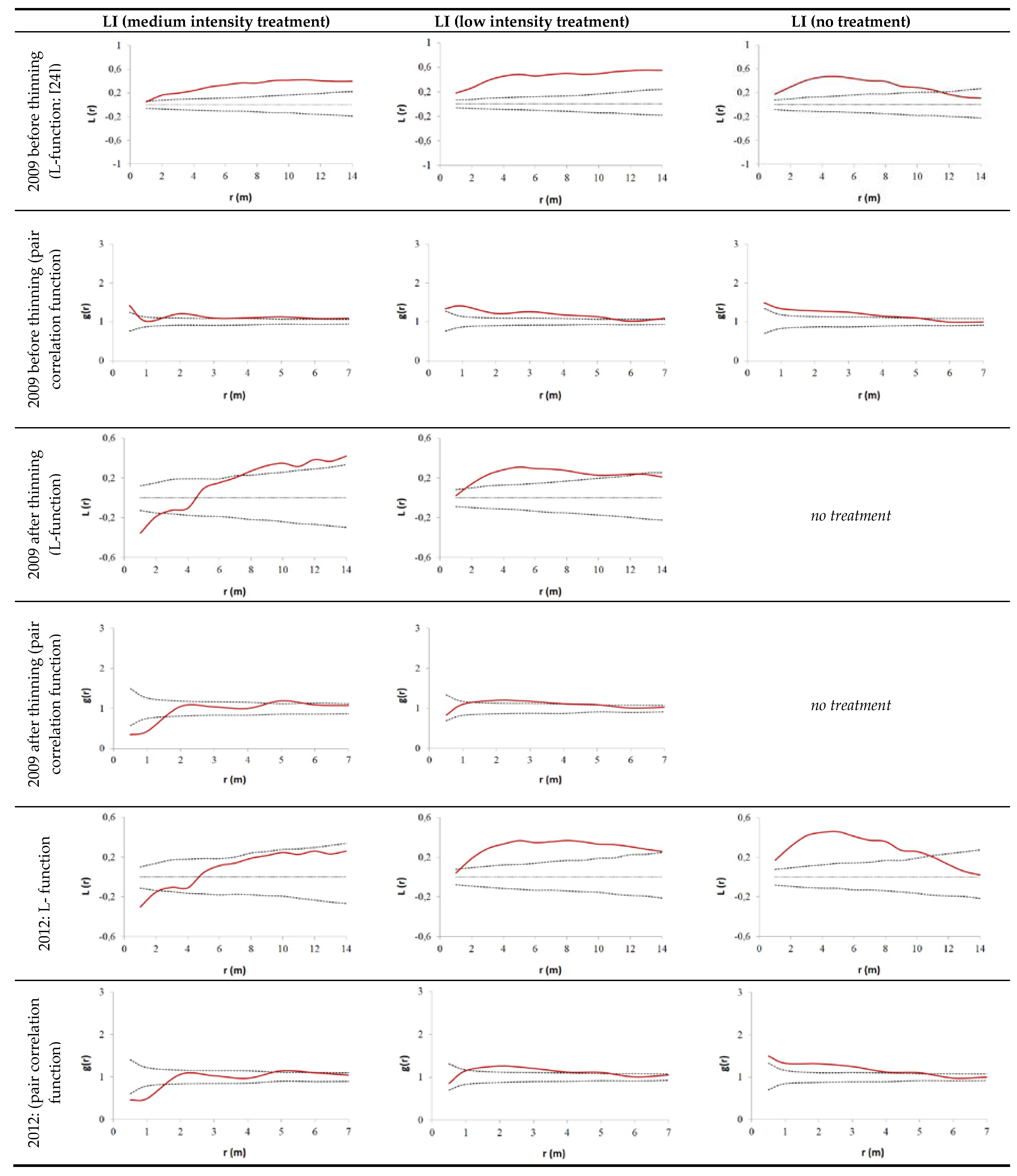
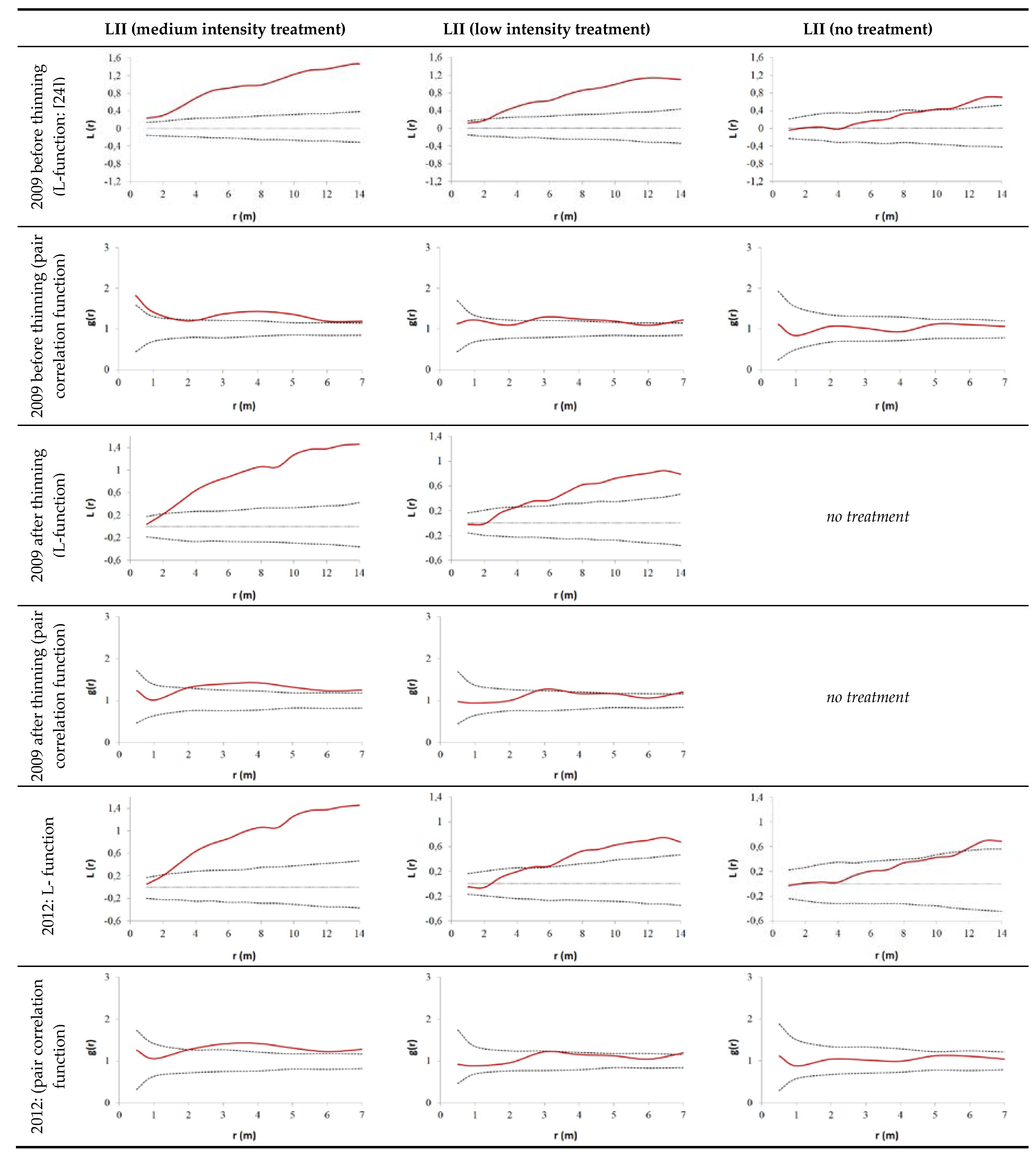
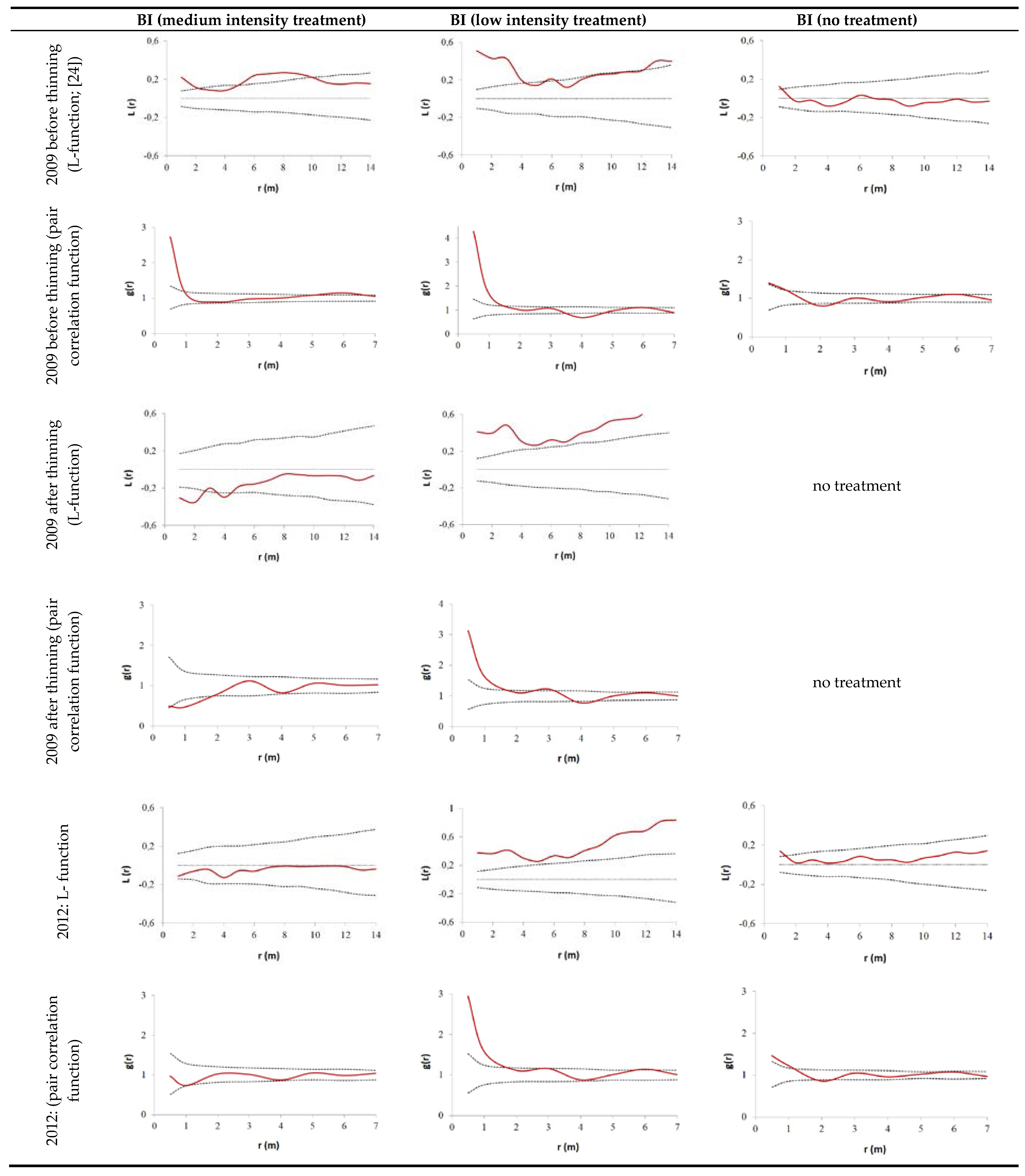
3.2. Thinning Impact on Spatial Tree Distribution Pattern
3.3. Quantification of Thinning Impact on Growth of Birch and Larch Trees
4. Discussion
4.1. Stand Level: Non-Spatial Forest Structure
4.2. Stand Level: Spatial Forest Structure
4.3. Single Tree Level
4.4. Management Issues: Development of a Mongolian Silviculture
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A. Pictures from the RA Altansumber




References
- Tsogtbaatar, J. Forest Policy Development in Mongolia. IUFRO Task Force Science/Policy Interface. p. 11. 2008. Available online: http://iufro-archive.boku.ac.at/iufro/taskforce/tfscipol/chennai-papers/ftsogtbaatar.pdf (accessed on 15 March 2016).
- Ykhanbai, H. Mongolian Forestry Outlook Study; Asia-Pacific Forestry Sector Outlook Study II; Working Paper Series; FAO: Bangkok, Thailand, 2010; p. 49. [Google Scholar]
- Gradel, A.; Petrow, W. Forstpolitische Entwicklungen im Transformationsland Mongolei. AFZ-DerWald. 2014, 17, 36–39. (In German) [Google Scholar]
- Yunatov, A.A. The Main Features of the Vegetation Cover of the Mongolian People’s Republic; Proceedings of the Mongolian Commission of the Academy of Sciences of the USSR: Moscow, Russia, 1950; p. 223. (In Russian) [Google Scholar]
- Dulamsuren, Ch. Floristische Diversität, Vegetation und Standortbedingungen in der Gebirgstaiga des Westkhentej, Nordmongolei; Universität Göttingen: Göttingen, Germany, 2004; p. 290. (In German) [Google Scholar]
- Mühlenberg, M.; Appelfelder, J.; Hoffmann, H.; Ayush, E.; Wilson, K.J. Structure of the montane taiga forests of West Khentii, Northern Mongolia. J. For. Sci. 2012, 58, 45–56. [Google Scholar]
- Ministry of Environment and Tourism. Multipurpose National Forest Inventory 2014–2016, 1st ed.; Ministry of Environment and Tourism: Ulaanbaatar, Mongolia, 2016. [Google Scholar]
- Dorjsuren, C. Forest Ecosystems (in Climate change impact and exposure). In Mongolia Second Assessment Report on Climate Change—MARCC 2014; Damdin, D., Zamba, B., Luvsan, N., Eds.; Ministry of Environment and Green Development: Ulaanbaatar, Mongolia, 2014; pp. 94–100. [Google Scholar]
- Antropov, V.F.; Seredkin, A.D.; Shhepin, A.A. Forestry in Buryatia; EKOS: Ulan-Ude, Russian Federation, 2013; p. 184. (In Russian) [Google Scholar]
- Martinsson, O.; Lesinski, J. Siberian Larch—Forestry and Timber in a Scandinavian Perspective; Jämtlands County Council Institute of Rural Development: Östersund, Sweden, 2007; p. 91. [Google Scholar]
- EIC, Environmental Information Centre Ulaanbaatar (Oin nociin medeellin san. Oin modny torol.). Available online: http://www.eic.mn/forestresource/forestresource.php?id=10 (accessed on 15 June 2016). (In Mongolian).
- Puhua, H. Betula platyphylla SUK. In Enzyklopädie der Holzgewächse—Handbuch und Atlas der Dendrologie; Schütt, P., Weisgerber, H., Schuck, H., Lang, U.M., Roloff, A., Eds.; Schütt, P., Translator; Ecomed: Landsberg am Lech, Germany, 2013; p. 6. (In German) [Google Scholar]
- Zyryanova, O.A.; Terazawa, M.; Koike, T.; Zyryanov, V.I. White birch trees as resource species of Russia: Their distribution, ecophysiological features, multiple utilizations. Eurasian J. For. Res. 2010, 13, 25–40. [Google Scholar]
- Hynynen, J.; Niemistö, P.; Viherä-Aarno, A.; Brunner, A.; Hein, S.; Velling, P. Silviculture of birch (Betula pendula Roth and Betula pubescens Ehrh.) in northern Europe. Forestry 2010, 83, 103–119. [Google Scholar] [CrossRef]
- Von Gadow, K. Forsteinrichtung. Analyse und Entwurf der Waldentwicklung; Universitätsverlag Göttingen, Reihe Universitätsdrucke: Göttingen, Germany, 2005; p. 342. (In German) [Google Scholar]
- Pretzsch, H. Forest Dynamics, Growth and Yield. From Measurement to Model; Springer: Berlin/Heidelberg, Germany, 2009; p. 664. [Google Scholar]
- Von Gadow, K.; Zhang, Y.C.; Wehenkel, C.; Pommerening, A.; Corral-Rivas, J.; Korol, M.; Myklush, S.; Hui, G.Y.; Kiviste, A.; Zhao, X.H. Forest Structure and Diversity. In Continuous Cover Forestry; Pukkala, T., von Gadow, K., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 29–83. [Google Scholar]
- Röhrig, E.; Bartsch, N.; Lüpke, B.V. Waldbau auf ökologischer Grundlage, 7th ed.; Eugen Ulmer: Stuttgart, Germany, 2006; p. 479. (In German) [Google Scholar]
- Assmann, E. Grundflächenhaltung und Zuwachsleistung Bayerischer Fichten-Durchforstungsreihen. Forstwiss. Cent. 1954, 73, 257–271. (In German) [Google Scholar] [CrossRef]
- Mäkinen, H.; Isomäki, A. Thinning intensity and growth of Norway spruce stands in Finland. Forestry 2004, 77, 349–364. [Google Scholar] [CrossRef]
- Gebhardt, T.; Häberle, K.H.; Matyssek, R.; Ammer, C. The more, the better? Water relations of Norway spruce stands after progressive thinning. Agric. For. Meteorol. 2014, 197, 235–243. [Google Scholar] [CrossRef]
- Olivar, J.; Bogino, S.; Rathgeber, C.; Bonnesoeur, V.; Bravo, F. Thinning has a positive effect on growth dynamics and growth-climate relationships in Aleppo pine (Pinus halepensis L.) trees of different crown classes. Ann. For. Sci. 2014, 71, 395–404. [Google Scholar] [CrossRef]
- Von Gadow, K. Messung und Modellforschung—Grundlagen der Forsteinrichtung. Allg. Forst Jagdztg. 2012, 184, 143–158. (In German) [Google Scholar]
- Gradel, A.; Ochirragchaa, N.; Altaev, A.A.; Voinkov, A.A.; Enkhtuya, B. Spatial distribution of trees on light taiga plots before selective thinning. Mong. J. Agric. Sci. 2015, 15, 91–99. [Google Scholar] [CrossRef]
- Gradel, A.; Ochirragchaa, N.; Altaev, A.A.; Voinkov, A.A.; Enkhtuya, B. Capacity development and forest research on the light taiga plots of the School of Agroecology and Business of the Mongolian University of Life Sciences in Darkhan. In Current Environmental Issues—Approaches to Solutions, Proceedings of the 20th Anniversary of the Professional Environmental Studies Program, Mongolian University of Life Sciences, Darkhan, Mongolia, 29 May 2015; pp. 44–51.
- Food and Agriculture Organization (FAO). Capacity Building and Institutional Development for Participatory Natural Resources Management and Conservation in Forest Areas of Mongolia (GCP/MON/002/NET); Financed by the Government of the Netherlands; FAO: Rome, Italy, 2006; p. 47. Available online: http://www.mne.mn/files/page792/Oi-4%20en.pdf (accessed on 30 June 2016).
- Kraznoshekov, Y.N. Soil Cover and Soils of Mountain forests of Northern Mongolia; Nauka: Novosibirsk, Russian Federation, 2013; p. 196. (In Russian) [Google Scholar]
- Von Gadow, K.; Hui, G.Y. Can the species-area relationship be derived from prior knowledge of the tree species richness? For. Stud./Metsanduslikud Uurim. 2007, 46, 13–22. [Google Scholar]
- Murray, D.M.; von Gadow, K. A flexible yield model for regional timber forecasting. South. J. Appl. For. 1993, 17, 112–115. [Google Scholar]
- Vitikova, L.; Dhubhain, A.N.; Pommerening, A. Agreement in Tree Marking: What is the uncertainty of human tree selection in selective forest management? For. Sci. 2016, 62, 288–296. [Google Scholar]
- Stoyan, D.; Stoyan, H. Fractals, Random Shapes, and Point Fields: Methods of Geometrical Statistics; Wiley: Chichester, UK, 1994; p. 406. [Google Scholar]
- Ripley, B.D. Spatial Statistics; Wiley: New York, NY, USA, 1981; p. 252. [Google Scholar]
- Wiegand, T. Introduction to Point Pattern Analysis with Ripley’s L and the O-Ring Statistic Using the Programita Software, 2nd ed.; UFZ-Centre for Environmental Research: Leipzig, Germany, 2004; p. 166. [Google Scholar]
- Besag, J. Contribution to the discussion of Dr. Ripley’s paper. J. R. Stat. Soc. B Met. 1977, 39, 193–195. [Google Scholar]
- Illian, J.; Penttinen, A.; Stoyan, H.; Stoyan, D. Statistical Analysis and Modelling of Spatial Point Patterns; Wiley: New York, NY, USA, 2008; p. 534. [Google Scholar]
- Zhang, C.; Zhao, X.; Gadow, K. Spatial distributions and spatial associations of dominant tree species in Korean pine broadleaved old-growth forests in Changbai Mountains. Balt. For. 2010, 16, 66–75. [Google Scholar]
- Wiegand, T.; Moloney, K.A. Rings, circles, and null-models for point pattern analysis in ecology. Oikos 2004, 104, 209–229. [Google Scholar] [CrossRef]
- Wiegand, T.; Moloney, K.A. A Handbook of Spatial Point Pattern Analysis in Ecology; Chapman and Hall/CRC Press: Boca Raton, FL, USA, 2014; p. 538. [Google Scholar]
- Mäkinen, H.; Vanninen, P. Effect of sample selection on the environmental signal derived from tree-ring series. For. Ecol. Manag. 1999, 113, 83–89. [Google Scholar] [CrossRef]
- Riemer, T. Über die Varianz von Jahrringbreiten—Statistische Methoden für die Auswertung der jährlichen Dickenzuwächse von Bäumen unter sich ändernden Lebensbedingungen; Forschungszentrum Waldökosysteme der Universität Göttingen: Göttingen, Germany, 1994; p. 375. (In German) [Google Scholar]
- Dulamsuren, C.; Hauck, M.; Leuschner, H.H.; Leuschner, C. Climate response of tree ring width in Larix sibirica growing in the drought-stressed forest-steppe ecotone of northern Mongolia. Ann. For. Sci. 2011, 68, 275–282. [Google Scholar] [CrossRef]
- Gärtner, H.; Nievergelt, D. The core-microtome: A new tool for surface preparation on cores and time series analysis of varying cell parameters. Dendrochronologia 2010, 28, 85–92. [Google Scholar] [CrossRef]
- Hegyi, F. A simulation model for managing jack-pine stands. In Growth Models for Tree and Stand Simulation; Royal College of Forestry: Stockholm, Sweden, 1974; pp. 74–90. [Google Scholar]
- Pretzsch, H. Zum Einfluß des Baumverteilungsmusters auf den Bestandeszuwachs. Jahrestagung Deutscher Verband Forstlicher Forschungsanstalten—Sektion Ertragskunde, Joachimsthal. Allg. Forst Jagdztg. 1995, 160, 190–200. (In German) [Google Scholar]
- Croсom, 3.0, Software Erstellt unter Delphi V (Münder, Schildbach und Schröder). Institut für Waldwachstum und Forstliche Informatik, Professur für Waldwachstums- und Holzmesskunde der TU Dresden: Tharandt, Germany, 2007. (In German)
- Münder, K. Konkurrenzuntersuchungen und Wachstumsmodellierung in Waldumbaubeständen des Mittleren Erzgebirges. Ph.D. Thesis, Technische Universität Dresden, Dresden, Germany, 2005; p. 160. (In German). [Google Scholar]
- Bachmann, M. Indizes zur Erfassung der Konkurrenz von Einzelbäumen. Methodische Untersuchungen in Bergmischwäldern; Forstliche Forschungsberichte München Nr. 171; Technische Universität München Wissenschaftszentrum Weihenstephan: München, Germany, 1998; p. 245. (In German) [Google Scholar]
- Crawley, M.J. The R-Book; Wiley: New York, NY, USA, 2007; p. 942. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009; p. 574. [Google Scholar]
- Robinson, A.P.; Hamann, J.D. Forest Analytics with R—An Introduction; Springer: New York, NY, USA, 2011; p. 354. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; Version 3.0.2; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 20 July 2016).
- Grothendieck, G. nls2: Non-Linear Regression with Brute Force. R Package Version 0.2. Available online: http://CRAN.R-project.org/package=nls2 (accessed on 30 May 2016).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Development Core Team. nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-113. Available online: http://packages.renjin.org/package/org.renjin.cran/nlme/3.1-113 (accessed on 29 March 2017).
- Bjornstad, O.N. ncf: Spatial Nonparametric Covariance Functions. R Package Version 1.1-5. Available online: http://CRAN.R-project.org/package=ncf (accessed on 29 May 2016).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 2nd ed.; Sage: Thousand Oaks, CA, USA; Available online: http://socserv.socsci.mcmaster.ca/jfox/Books/Companion (accessed on 30 April 2016).
- Sarkar, D. Lattice: Multivariate Data Visualization with R; Springer: New York, NY, USA, 2008; p. 268. [Google Scholar]
- Von Gadow, K.; Hui, G.Y. Characterizing forest spatial structure and diversity. In Sustainable Forestry in Temperate Regions; Björk, L., Ed.; SUFOR, University of Lund: Lund, Sweden, 2002; pp. 20–30. [Google Scholar]
- Huss, J. Zur Durchforstung engbegründeter Fichtenjungbestände. Forstwiss. Cent. 1990, 109, 101–118. (In German) [Google Scholar] [CrossRef]
- Juodvalkis, A.; Kairiukstis, L.; Vasiliauskas, R. Effects of thinning on growth of six tree species in north-temperate forests of Lithuania. Eur. J. For. Res. 2005, 124, 187–192. [Google Scholar] [CrossRef]
- Štefančík, I. Development of target (crop) trees in beech (Fagus sylvatica L.) stand with delayed initial tending and managed by different thinning methods. J. For. Sci. 2013, 59, 253–259. [Google Scholar]
- Pretzsch, H. Stand density and growth of Norway spruce (Picea abies (L.) Karst.) and European beech (Fagus sylvatica L.): Evidence from long-term experimental plots. Eur. J. For. Res. 2005, 124, 193–205. [Google Scholar] [CrossRef]
- Gizachew, B.; Brunner, A. Density–growth relationships in thinned and unthinned Norway spruce and Scots pine stands in Norway. Scand. J. For. Res. 2011, 26, 543–554. [Google Scholar] [CrossRef]
- Gradel, A.; Mühlenberg, M. Spatial characteristics of near-natural Mongolian forests at the southern edge of the taiga. Allg. Forst Jagdztg. 2011, 182, 40–52. [Google Scholar]
- Lei, X.; Lu, Y.; Peng, C.; Zhang, X.; Cnag, J.; Hong, L. Growth and structure of semi-natural larch-spruce-fir (Larix olgensis-Picea jezoensis-Abies nephrolepis) forests in northeast-China: 12-year result after thinning. For. Ecol. Manag. 2012, 240, 165–177. [Google Scholar] [CrossRef]
- Bachofen, H.; Zingg, A. Effectiveness of structure improvement thinning on stand structure in subalpine Norway spruce (Picea abies (L.) Karst.) stands. For. Ecol. Manag. 2001, 145, 137–149. [Google Scholar] [CrossRef]
- Pukkula, T.; Lähde, E.; Laiho, O. Which trees should be removed in thinning treatments? For. Ecosyst. 2015, 2. [Google Scholar] [CrossRef]
- Gradel, A.; Haensch, C.; Batsaikhan, G.; Batdorj, D.; Ochirragchaa, N.; Günther, B. Response of white birch (Betula platyphylla Sukaczev) to temperature and precipitation in the mountain forest steppe and taiga of northern Mongolia. Dendrochronologia 2017, 41, 24–33. [Google Scholar] [CrossRef]
- Gradel, A.; Batsaikhan, G.; Ochirragchaa, N.; Batdorj, D.; Kusbach, A. Climate-growth relationships and pointer year analysis of a Siberian larch (Larix sibirica Ledeb.) chronology in the Mongolian mountain forest steppe. For. Ecosyst. Submitted.
- Abetz, P. Reaktionen auf Standraumerweiterung und Folgerungen fuer die Auslesedurchforstung bei Fichte. Allg. Forst Jagdztg. 1976, 147, 72–75. (In German) [Google Scholar]
- Zhang, S.; Burkhart, H.E.; Amateis, R.L. The influence of thinning on tree height and diameter relationships in loblolly pine plantations. South. J. Appl. For. 1997, 21, 199–205. [Google Scholar]
- Ammer, C.; Ziegler, C.; Knoke, T. Zur Beurteilung von intra- und interspezifischer Konkurrenz von Laubbaumbeständen im Dickungsstadium. Allg. Forst Jagdztg. 2005, 176, 85–94. (In German) [Google Scholar]
- Roberts, S.D.; Harrington, C.A. Individual tree growth response to variable-density thinning in coastal Pacific Northwest forests. For. Ecol. Manag. 2008, 255, 2771–2781. [Google Scholar] [CrossRef]
- Klädtke, J.; Kohnle, U.; Kublin, E.; Ehring, A.; Pretzsch, H.; Uhl, E.; Spellmann, H.; Weller, A. Wachstum und Wertleistung der Douglasie in Abhängigkeit von der Standraumgestaltung. Schweiz. Z. Forstwes. 2012, 163, 96–104. (In German) [Google Scholar] [CrossRef]
- Karlman, L.; Mörling, T.; Martinsson, O. Wood density, annual ring width and latewood content in Larch and Scots pine. Eurasian J. For. Res. 2005, 8, 91–96. [Google Scholar]
- Heräjärvi, H. Technical properties of mature birch (Betula pendula and B. pubescens) for saw milling in Finland. Silva Fenn. 2001, 35, 469–485. [Google Scholar] [CrossRef]
- Cameron, A.D.; Dunham, R.A.; Petty, J.A. The effects of heavy thinning on stem quality and timber properties of silver birch (Betula pendula Roth). Forestry 1995, 68, 275–285. [Google Scholar] [CrossRef]
- Oikarinen, M. Growth and yield models for silver birch (Betula pendula) plantations in southern Finland. In Communicationes Instituti Forestalis Fenniae 113; Finnish Forest Research Institute (Luke): Helsinki, Finland, 1983; pp. 1–75. (In Finnish) [Google Scholar]
- Rytter, L.; Karlsson, A.; Karlsson, M.; Stener, L.G. Skötsel av björk, al och asp. In Skogsskötselserien nr 9, 1st ed.; Swedish Forest Agency (Skogsstyrelsen): Helsinki, Sweden, 2008; pp. 1–122. (In Swedish) [Google Scholar]
- Khutakova, S.V.; Ubugunova, V.I.; Gradel, A.; Enkhtuya, B. Morphogenetic features of soils of larch forests of terrain Altan Sumber Orkhon-Selenga Middle Mountain. In Central Asian Environmental and Agricultural Problems, Potential Solutions; Institute of Plant and Agricultural Sciences, School of Agroecology and Business of the MULS in Darkhan: Darkhan, Mongolia, 2016; pp. 127–130. (In Russian) [Google Scholar]
- Sugimoto, A.; Yanagisawa, N.; Naito, D.; Fujita, N.; Maximov, T.C. Importance of permafrost as a source of water for plants in east Siberian taiga. Ecol. Res. 2002, 17, 493–503. [Google Scholar] [CrossRef]
- Savin, E.N.; Milyutin, L.I.; Krasnoshhekov, Ju.N.; Korotkov, I.A.; Suncov, A.V.; Dugarzhav, Ch.; Cogoo, Z.; Dorzhsuren, Ch.; Zhamjansurjen, S.; Gombosuren, N. Forests of the Mongolian People’s Republic (Larch forests of the Eastern Khentey), Soviet-Mongolian Expedition; Nauka, Siberian Department: Novosibirsk, Russia, 1988; p. 176. (In Russian) [Google Scholar]
- Kopp, B.J.; Minderlein, S.; Menzel, L. Soil moisture dynamics in a mountainous headwater area in the discontinuous permafrost zone of northern Mongolia. Arct. Alp. Res. 2014, 46, 459–470. [Google Scholar] [CrossRef]
- Vitikova, L. Transformation to Continuous Cover Forestry in Ireland. Ph.D. Thesis, University College Dublin, Dublin, Ireland, 2014; p. 159. [Google Scholar]
- Cameron, A.D. Managing birch woodlands for the production of quality timber. Forestry 1996, 69, 357–371. [Google Scholar] [CrossRef]
- Karlsson, M. Natural Regeneration of broadleaved tree Species in Southern Sweden. Ph.D. Thesis, Swedish University of Agricultural Sciences, Southern Swedish Forest Research Centre, Alnarp. Acta Universitatis Agriculturae Sueciae, Silvestria, Sweden, 2001; p. 44. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference Stand | Main Tree Species | Height above Sea Level | Exposition | N (Plots) | Plot Size (m2) 2009 | Indication of Disturbances | Year of First Assessment | |
|---|---|---|---|---|---|---|---|---|
| BI * | birch | 934 | N | 3 | 2500 (1550) | s.f. | 2009 | |
| BII | birch | 966 | N | 3 | 2500 | s.f. | 2009 | |
| LI | larch | 911 | NW | 3 | 2500 | s.f. | 2009 | |
| LII | larch | 976 | NW | 3 | 2500 | s.f., s.p.l. | 2009 |
| Forest Stand | N/ha | Dg | D | Dg_200 | SD | D_200 | SD | Stand Age | SD | Age (N ) |
|---|---|---|---|---|---|---|---|---|---|---|
| BI | 1229 | 11.2 | 10.4 | 18.8 | 3.650 | 18.4 | 1.436 | 44 | 18.820 | 38 |
| BII | 1103 | 14.2 | 13.6 | 20.1 | 2.913 | 19.9 | 1.478 | 68 | 13.314 | 34 |
| LI | 1389 | 11.9 | 11.0 | 19.4 | 6.891 | 18.2 | 3.987 | 22 | 2.392 | 22 |
| LII | 565 | 22.8 | 21.8 | 29.2 | 4.257 | 28.8 | 2.999 | 61 | 3.296 | 36 |
| Stand | Plot | 2009before | 2009after | 2012 | Stand | Plot | 2009before | 2009after | 2012 |
|---|---|---|---|---|---|---|---|---|---|
| N/ha | N/ha | ||||||||
| BI | m. int. | 1144 | 568 | 736 | LI | m. int. | 1528 | 776 | 868 |
| low int. | 1174 | 961 | 1045 | low int. | 1504 | 1148 | 1268 | ||
| unth. | 1368 | unth. | 1510 | unth. | 1136 | unth. | 1200 | ||
| BII | m. int. | 984 | 484 | 500 | LII | m. int. | 656 | 524 | 524 |
| low int. | 1136 | 808 | 796 | low int. | 624 | 564 | 624 | ||
| unth. | 1188 | unth. | 1192 | unth. | 416 | unth. | 420 | ||
| BA (m2)/ha | BA/ha | ||||||||
| BI | m. int. | 14.659 | 6.974 | 8.567 | LI | m. int. | 16.707 | 11.039 | 14.404 |
| low int. | 8.690 | 6.988 | 8.746 | low int. | 13.513 | 11.320 | 15.165 | ||
| unth. | 10.841 | unth. | 12.223 | unth. | 15.657 | unth. | 18.749 | ||
| BII | m. int. | 17.289 | 10.791 | 11.366 | LII | m. int. | 24.878 | 22.026 | 23.692 |
| low int. | 17.387 | 13.359 | 14.310 | low int. | 23.025 | 21.786 | 23.025 | ||
| unth. | 17.895 | unth. | 19.249 | unth. | 21.635 | unth. | 23.154 | ||
| dom. height (m) | dom. height (m) | ||||||||
| BI | m. int. | 12.3 | 11.8 | 11.9 | LI | m. int. | 12.1 | 12.0 | 12.6 |
| low int. | 10.2 | 10.3 | 10.7 | low int. | 11.4 | 11.4 | 12.0 | ||
| unth. | 11.0 | unth. | 11.6 | unth. | 12.8 | unth. | 13.1 | ||
| BII | m. int. | 14.9 | 14.8 | 15.0 | LII | m. int. | 16.4 | 16.4 | 16.5 |
| low int. | 14.4 | 14.4 | 14.6 | low int. | 16.0 | 16.0 | 16.0 | ||
| unth. | 14.5 | unth. | 14.7 | unth. | 16.5 | unth. | 16.7 | ||
| CV | CV | ||||||||
| BI | m. int. | 0.424 | 0.426 | 0.416 | LI | m. int. | 0.359 | 0.373 | 0.364 |
| low int. | 0.251 | 0.246 | 0.251 | low int. | 0.311 | 0.316 | 0.310 | ||
| unth. | 0.352 | unth. | 0.329 | unth. | 0.473 | unth. | 0.449 | ||
| BII | m. int. | 0.333 | 0.279 | 0.286 | LII | m. int. | 0.362 | 0.342 | 0.340 |
| low int. | 0.276 | 0.263 | 0.263 | low int. | 0.269 | 0.260 | 0.267 | ||
| unth. | 0.278 | unth. | 0.232 | unth. | 0.249 | unth. | 0.252 | ||
| Stand | Plot | 2009before | 2009after | 2012 | Stand | Plot | 2009before | 2009after | 2012 |
|---|---|---|---|---|---|---|---|---|---|
| D | D | ||||||||
| BI | m. int. | 11.8 | 11.5 | 11.2 | LI | m. int. | 11.1 | 12.6 | 13.7 |
| low int. | 9.4 | 9.3 | 10.0 | low int. | 10.2 | 10.7 | 11.8 | ||
| unth. | 9.5 | unth. | 9.7 | unth. | 12.0 | unth. | 12.9 | ||
| BII | m. int. | 14.0 | 16.0 | 16.4 | LII | m. int. | 20.7 | 21.9 | 22.7 |
| low int. | 13.4 | 14.0 | 14.6 | low int. | 20.8 | 21.5 | 20.9 | ||
| unth. | 13.3 | unth. | 13.8 | unth. | 24.9 | unth. | 30.9 | ||
| Dg | Dg | ||||||||
| BI | m. int. | 12.8 | 12.4 | 12.2 | LI | m. int. | 11.8 | 13.5 | 14.5 |
| low int. | 9.7 | 9.6 | 10.3 | low int. | 10.7 | 11.2 | 12.4 | ||
| unth. | 10.0 | unth. | 10.2 | unth. | 13.2 | unth. | 14.1 | ||
| BII | m. int. | 15.0 | 16.8 | 17.0 | LII | m. int. | 22.0 | 23.1 | 24.0 |
| low int. | 13.9 | 14.5 | 15.1 | low int. | 21.7 | 22.2 | 22.0 | ||
| unth. | 13.8 | unth. | 14.3 | unth. | 25.7 | unth. | 26.5 | ||
| D_200 | D_200 | ||||||||
| BI | m. int. | 21.1 | 16.8 | 17.7 | LI | m. int. | 18.0 | 17.9 | 20.0 |
| low int. | 13.4 | 12.7 | 14.0 | low int. | 15.6 | 15.6 | 17.5 | ||
| unth. | 16.1 | unth. | 16.3 | unth. | 20.3 | unth. | 21.8 | ||
| BII | m. int. | 21.0 | 20.3 | 21.0 | LII | m. int. | 28.9 | 28.7 | 29.8 |
| low int. | 19.4 | 19.0 | 19.8 | low int. | 22.2 | 22.6 | 28.4 | ||
| unth. | 19.4 | unth. | 20.2 | unth. | 30.0 | unth. | 30.9 | ||
| Dg_200 | Dg_200 | ||||||||
| BI | m. int. | 21.3 | 17.4 | 18.2 | LI | m. int. | 19.0 | 18.9 | 20.8 |
| low int. | 13.7 | 13.0 | 14.3 | low int. | 16.4 | 16.4 | 18.2 | ||
| unth. | 16.6 | unth. | 16.8 | unth. | 22.1 | unth. | 23.4 | ||
| BII | m. int. | 21.4 | 20.4 | 21.2 | LII | m. int. | 29.5 | 29.3 | 30.4 |
| low int. | 19.5 | 19.1 | 19.9 | low int. | 27.5 | 27.5 | 28.6 | ||
| unth. | 19.5 | unth. | 20.3 | unth. | 30.2 | unth. | 31.1 | ||
| Model | Species | Variable | Fixed Effects | Competitor Selection | Degrees of Freedom | Model Parameter (Fixed Effects) | AIC of the Model | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Intercept | p-Value | CIrel | p-Value | CIdiff | p-Value | DBH | p-Value | |||||||
| 1 | Birch | abs.gr | CIdiff + DBH | cone | 31 | −0.0009 | 0.0485 | 0.0003 | 0.0487 | 0.0002 | 0.0000 | −400.95 | ||
| Birch | abs.gr | CIdiff + DBH | NNDSR | 37 | −0.0008 | 0.1638 | 0.0003 | 0.0214 | 0.0002 | 0.0000 | −455.48 | |||
| Larch | abs.gr | CIdiff + DBH | cone | 27 | −0.0013 | 0.5163 | 0.0008 | 0.0440 | 0.0002 | 0.0000 | −336.97 | |||
| Larch | abs.gr | CIdiff + DBH | NNDSR | 31 | −0.0018 | 0.2836 | 0.0006 | 0.0044 | 0.0002 | 0.0000 | −388.26 | |||
| 2 | Birch | rel.gr | CIrel | cone | 29 | 2.1084 | 0.0000 | 0.7111 | 0.0387 | 96.48 | ||||
| Birch | rel.gr | CIrel | NNDSR | 29 | 1.9163 | 0.0000 | 1.2444 | 0.0052 | 87.82 | |||||
| Larch | rel.gr | CIrel | cone | 31 | 1.4020 | 0.0000 | 0.6776 | 0.0159 | 70.17 | |||||
| Larch | rel.gr | CIrel | NNDSR | 36 | 1.1756 | 0.0000 | 1.4156 | 0.0001 | 68.64 | |||||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gradel, A.; Ammer, C.; Ganbaatar, B.; Nadaldorj, O.; Dovdondemberel, B.; Wagner, S. On the Effect of Thinning on Tree Growth and Stand Structure of White Birch (Betula platyphylla Sukaczev) and Siberian Larch (Larix sibirica Ledeb.) in Mongolia. Forests 2017, 8, 105. https://doi.org/10.3390/f8040105
Gradel A, Ammer C, Ganbaatar B, Nadaldorj O, Dovdondemberel B, Wagner S. On the Effect of Thinning on Tree Growth and Stand Structure of White Birch (Betula platyphylla Sukaczev) and Siberian Larch (Larix sibirica Ledeb.) in Mongolia. Forests. 2017; 8(4):105. https://doi.org/10.3390/f8040105
Chicago/Turabian StyleGradel, Alexander, Christian Ammer, Batsaikhan Ganbaatar, Ochirrragchaa Nadaldorj, Batdorj Dovdondemberel, and Sven Wagner. 2017. "On the Effect of Thinning on Tree Growth and Stand Structure of White Birch (Betula platyphylla Sukaczev) and Siberian Larch (Larix sibirica Ledeb.) in Mongolia" Forests 8, no. 4: 105. https://doi.org/10.3390/f8040105
APA StyleGradel, A., Ammer, C., Ganbaatar, B., Nadaldorj, O., Dovdondemberel, B., & Wagner, S. (2017). On the Effect of Thinning on Tree Growth and Stand Structure of White Birch (Betula platyphylla Sukaczev) and Siberian Larch (Larix sibirica Ledeb.) in Mongolia. Forests, 8(4), 105. https://doi.org/10.3390/f8040105