Our results quantify the growth response of European beech on the tree- and stand-level to different thinning intensities on two sites with opposite aspects in Southwest Germany. We could show that the thinning effect is influenced not only by thinning intensity and initial tree size, but also by the status of a tree within a stand and by site aspect.
4.2. Effect of Tree Size
For the individual tree-level model, most of the variance was explained by initial tree size (
Table 5). Tree size at the beginning of the experiment represents the starting point in any growth modeling according to Assmann [
54] and Wykoff [
46]. The relation is positive,
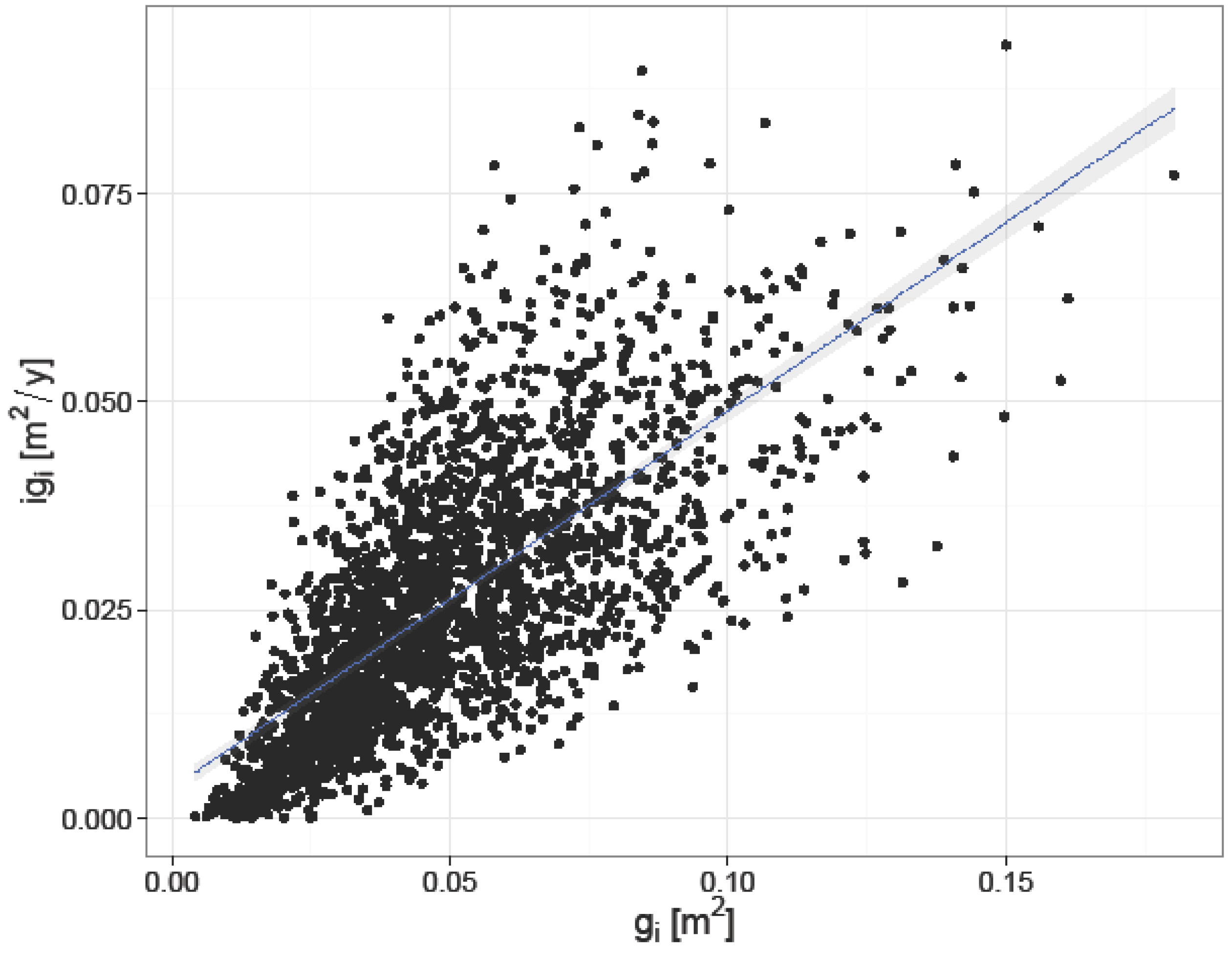
i.e., the larger the initial basal area was, the higher the basal area increment. The relationship between
gi and
igi followed a linear model (
Figure 4). Our results are in agreement with those of Monserud and Sterba [
47] and Cescatti and Piutti [
27], who found that most of the variance in their basal area increment models for individual trees was explained by size factors.
Pretzsch and Dieler [
55] found a steeper slope of the linear size-growth relationship on fertile sites than on infertile sites. Our results correspond to Pretzsch and Dieler [
55], in that the size-growth relationship is represented by a straight line and that the increment is higher on more fertile sites (in our case, NE aspect). An interaction term between initial basal area and aspect was not included in our final model due to a lower AIC value; therefore, our results do not differentiate between aspects. Nevertheless, the three thinning treatments (CT, ST, VT) may cause the size-growth relationship to change in case of stands with different competition levels. Moreover, for the current analyses a three-way interaction term (
i.e., interaction between competition indexes, tree size, and aspect) was not included due to the different objectives we had.
4.3. Effect of Competition
At the stand-level the competition factor represented by the thinning treatment had the most significant effect on stand growth. This is in agreement with many other studies which highlight the importance of competition effects on growth patterns and climate sensitivity of European beech stands under different silvicultural treatments [
19,
24,
26,
27] and shows that the average stand diameter increment consistently increased with increasing thinning intensity (
Figure 2). The mean diameter increment was also calculated for the 100 largest trees in diameter per hectare as differences in average stand diameter increment between differently thinned plots can be caused by the different number of stems per hectare [
56]. The results showed the same trend of higher increment in plots where stronger thinning has been applied, which is in accordance with findings from van der Maaten [
19], Le Goff and Ottorini [
26], and Klädtke [
57], who report that the radial increment of European beech increases with the intensity of thinning and follow similar patterns for different thinning intensities. Likewise, work by Mäkinen and Isomäki [
56] and Çiçek
et al. [
58] found that larger trees are also capable to increase their diameter after thinning.
With respect to height growth, the larger height increment on the NE aspect was a clear indication of a higher site index. However, we found that the height increment was not significantly different between treatments and aspects and assume that this is due to the relatively low sample size and the high error probability during the assessment of tree height in the field. Similar results regarding the thinning effect on tree growth were found for various broadleaved species [
59,
60,
61] where stand density had a significant effect on diameter growth but not on height growth.
Our focus was to analyze the effect of wide spacing on growth of European beech trees in order to increase the production of high quality timber. Regarding the loss in volume increment, our results are similar to the results of Badoux [
62] and Assmann [
54], who investigated the growth response of European beech in long-term thinning experiments. They concluded that a very strong thinning regime decreases the volume increment of European beech stands by 20% and only occasionally, and for short phases, can it lead to a temporary acceleration of current volume increment. In our study no such growth-stimulating effect has been found on NE or SW aspects, and a higher reduction in volume increment has been registered. This effect could be explained by the intensity of thinning, as most of the thinnings in the Badoux [
62] and Assmann [
54] studies were less intensive (stand basal area after thinning 25–35 m
2/ha for heavily thinned stands) than those in this study (10 and 15 m
2/ha). The stand basal area in our thinned plots was reduced by 41% on the NE and 31% on the SW aspect with strong thinning, and up to 60% in very strong thinning. In addition to this, the volume increment of the thinned plots may be, to some extent, overestimated as the thinning accelerates the radial increment at 1.3 m more than at other heights within the tree [
63].
Changes in stand basal area increment in response to different thinning intensities it is often not noticeable on the stand-level but on the individual tree-level [
64]. Additionally, in our study, the positive effect of thinning on basal area increment was captured only at the individual tree-level.
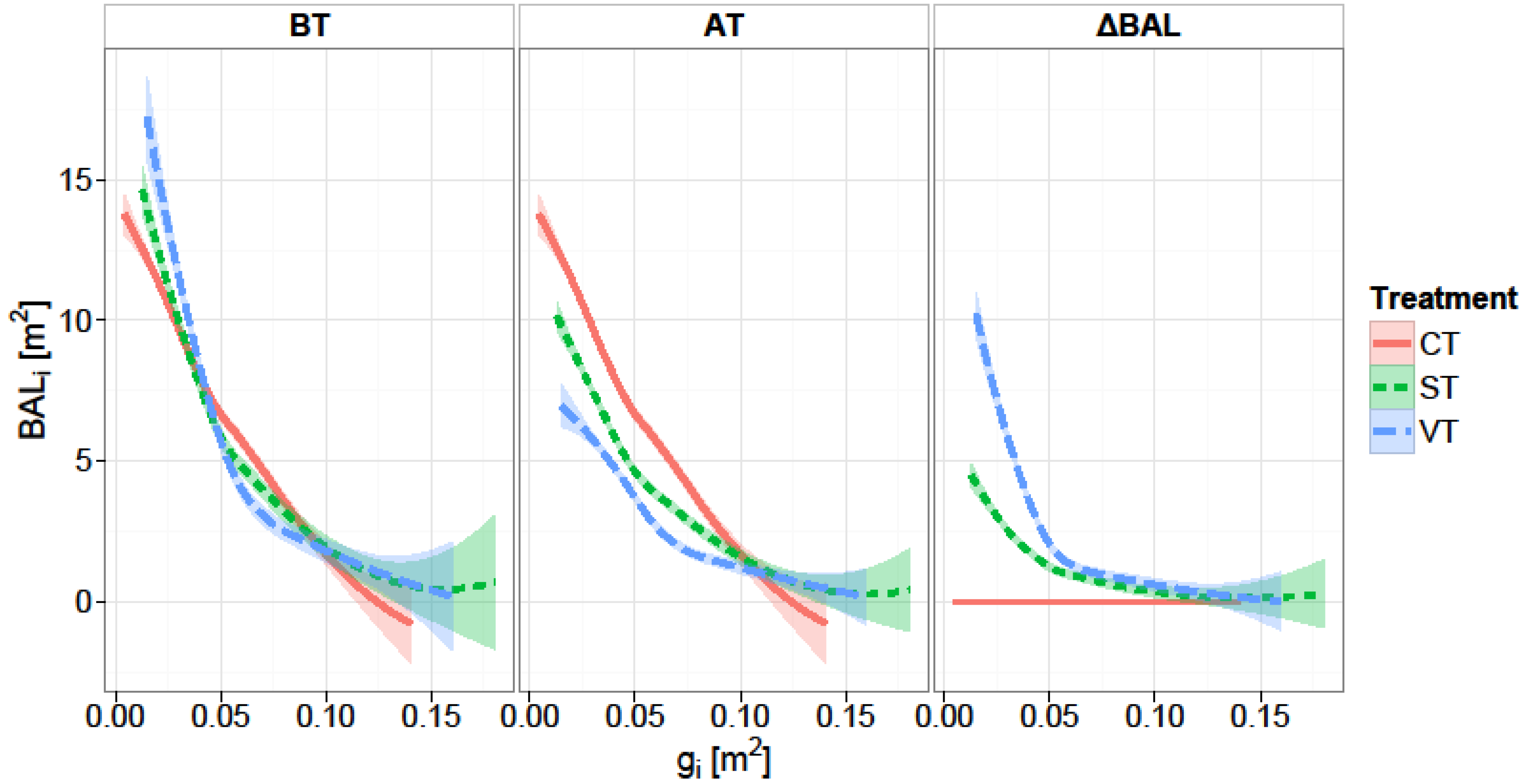
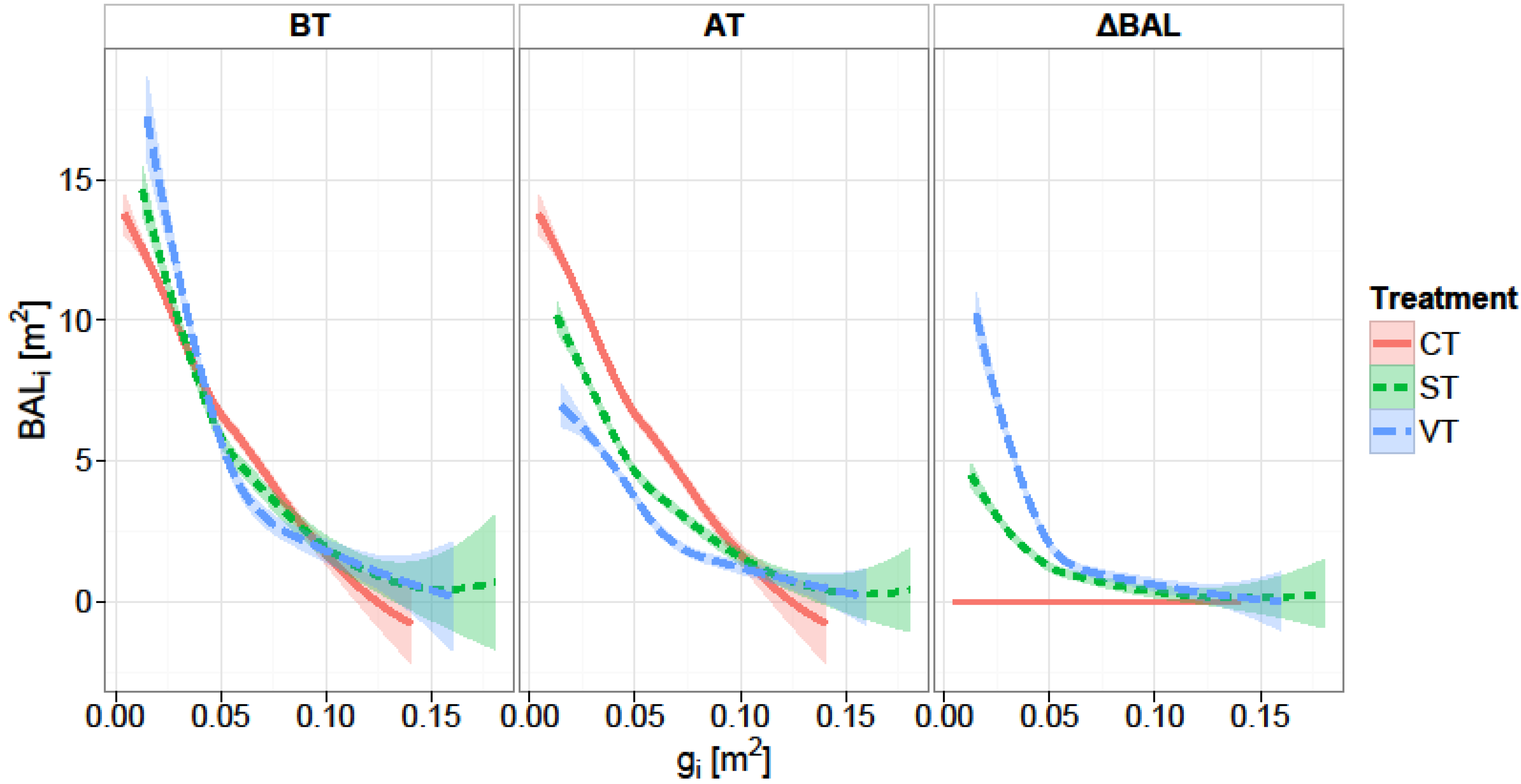
We used a distance-independent competition index, the Δ
BALi. Basal area of larger trees is commonly used as a measure of competition in individual-tree growth models [
46,
47,
52]. In a study concerning the effect of competition on individual tree basal area growth Rivas [
65] compared competition indexes, and concluded that distance-independent competition indexes performed as well as distance-dependent ones. In a similar study predicting basal area increment of individual trees, Ledermann [
66] used a semi-distance-independent competition indices (a distance independent competition measure calculated in very small sample plots including the subjected tree) and found that the semi-distance-independent competition indices explain as much variation as any of the analyzed distance-dependent ones. The significant interaction effect of initial basal area and release intensity on the increment of individual trees clearly indicates that the initial size of a tree determines its potential to respond to thinning. A significant influence of
BAL and different formulations of
BAL on basal area increment of individual trees has been found also by Wykoff [
46], Monserud and Sterba [
47], Laubhann [
48], and Stage and Ledermann [
67].
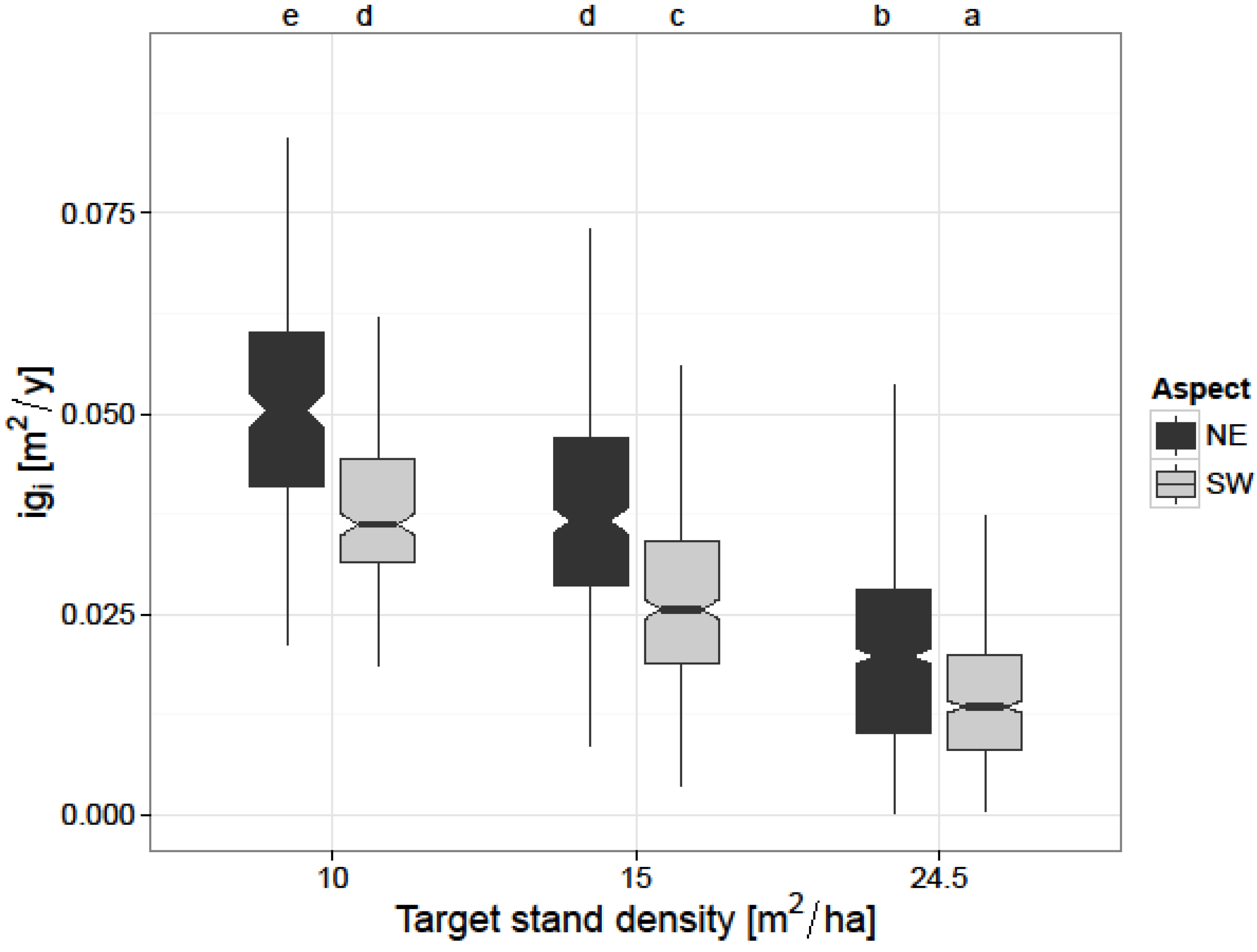
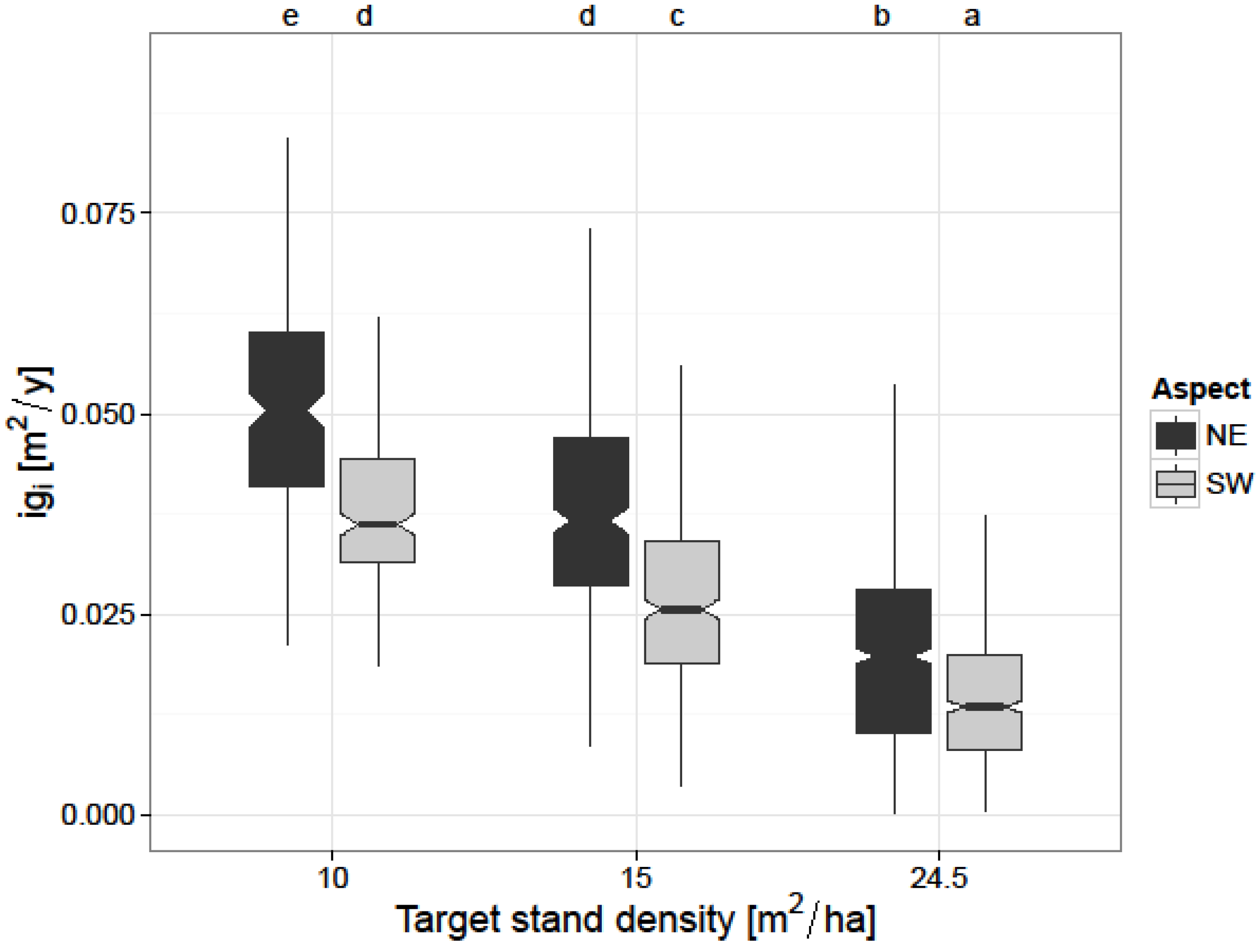
The second competition-related factor included in the model—target stand density—provided a major influence on basal area increment of individual trees, as well. This is in accordance with other authors who studied the effect of thinning on European beech stands [
24,
57,
68,
69]. It also corresponds with van der Maaten [
19], who found similar results in the study area regarding radial growth for the shorter time period 2001–2006. The increase in
igi was larger for the plots with a target basal area of 10 m
2/ha as compared to the control plots. Our results are similar with the results of Le Goff and Ottorini [
26] and Boncina [
24], who found larger growth on the individual tree-level with increasing thinning intensity in European beech stands. The large influence of target stand density on individual tree growth (
Table 5) highlights the importance of resource limitation within a stand. Contrary to Geßler [
21] who found that reducing stand density can negatively affect individual tree growth of European beech through increased soil evaporation and transpiration of understory vegetation, our results show that a reduction in stand basal area per hectare stimulates individual tree growth even under the warmer and drier climatic conditions on the SW aspect (
Figure 3).
In the context of climate change adaptation, the future of European beech is controversially discussed [
13,
16,
70,
71]. Our results show that a high intensity crop-tree thinning regime, which reduces the stand basal area to 15 and 10 m
2/ha, respectively, at the age of 80–100 years, can still increase the growth of European beech in a warmer and drier climate.
4.4. Effect of Aspect
The site represents a major component influencing tree growth [
72]. This was also visible in our study at both levels of the analysis. At the stand-level, besides treatment the aspect had a significant influence on volume and diameter growth of dominant trees, on the SW aspect being significantly lower than that on the NE. This highlights the importance of site conditions, and suggests that warmer climatic conditions could represent a limiting factor for European beech growth. However, potential confounding effects could compromise this finding. This is due to the fact that in our study, aside from local climatic conditions, soil structure and soil depth also differed between aspects. Our findings are contrary to the results of van der Maaten [
73], whose study on seasonal stem growth patterns of European beech was conducted in the same study area. He did not find a significant effect of aspect on the growth response to thinning. This might be related to differences in the length of the study period (six
versus 13 years). In a recent study assessing the effects of competition on climate tree-growth relationships of European beech trees growing in contrasting ecological conditions, Lebourgeois [
74] did not find significant differences between sites on north- and south-facing slopes.
One of the most interesting findings in our study corresponds to the quantitative insights into the competitive relationships of individual trees growing on two different aspects. These insights were assessed by evaluating the interaction effect between aspect and target stand density at the tree-level, and between aspect and treatment at the stand-level. The relationship was significant (
p ≤ 0.05) only at the individual tree-level; the basal area increment of trees on the SW facing slope under VT treatment was significantly lower than on the opposite slope. The relationship was not significant at the stand-level and we assume that this is due to compensatory stand-level effects and, therefore, might not have been detected due to the restricted sample size (
n = 15) compared with the larger sample size at the individual tree-level (
n = 2458). It is interesting to note that trees growing at very low stand density produce a significantly larger increment on sites with less irradiation and more favorable moisture conditions. The results are in agreement with Assmann [
54] and Pretzsch [
25], who found that the thinning response of European beech is larger on favorable sites than on unfavorable sites. This significant interaction between aspect and stand density on tree growth response to thinning implies that, under warmer and drier climatic conditions, thinning can stimulate tree growth but to a smaller extent as it does on more favorable site conditions. The site-specific thinning response present in our results is probably a consequence of the sensitivity of beech to changes in water supply. In our study we studied site effects on the growth response to thinning by comparing sites differing in aspect without considering other site characteristics (e.g., elevation, slope steepness, soil structure, soil depth) and climate variability (e.g., temperature, precipitation, drought), which are also relevant for growth. These aspects were considered in previous studies done by Hildebrand
et al. [
33], Mayer
et al. [
35] and van der Maaten [
73]. Moreover, since our study is not replicated at other sites our findings are valid only for the conditions in our study region and for a generalization of our results broader analyses need to be carried out.




{kind=link}
{kind=link}
{kind=link}
{kind=link}